Павлов И.П. Полное собрание сочинений. Том 4. [Иван Петрович Павлов] (fb2) читать онлайн
- Павлов И.П. Полное собрание сочинений. Том 4. 13.45 Мб скачать: (fb2) - (исправленную) читать: (полностью) - (постранично) - Иван Петрович Павлов
[Настройки текста] [Cбросить фильтры]
[Оглавление]




«Лекции работе больших полушарий головного мозга», читанные И. П. Павловым в 1924 г. на кафедре физиологии Военно-медицинской академии, впервые были опубликованы в 1927 г. В том же году вышло второе издание «Лекций».
В ноябре 1935 г. И. П. Павлов подготовил к печати третье издание «Лекций», вышедшее в свет в 1937 г. Все три издания содержат идентичный текст.
«Лекции» стереотипно были воспроизведены в «Полном собрании трудов" и так же воспроизводятся в настоящем издании «Полного собрания сочинений» И. П. Павлова.


Предисловие к первому изданию
Весной 1924 года в Военно-медицинской академии перед врачебной и естественно-научной аудиторией я в ряде лекций старался дать полное систематическое изложение нашей тогда почти двадцатипятилетней работы над физиологией больших полушарий собаки. Эти лекции были застенографированы, и я рассчитывал их потом опубликовать. Но, взявшись за просмотр стенографической записи, я нашел мое изложение неудовлетворительным и принялся за его существенную переделку. Эта работа заняла у меня полтора года, во время которых в заведуемых мной лабораториях продолжалась оживленная разработка того же предмета. За этот срок в ранее отделанных лекциях кое-что из фактического материала претерпело значительное изменение и исправление. Но в теперь выпускаемой книге я оставляю все так, как было первоначально написано, со специальным умыслом, чтобы, передавая в позже законченных лекциях новейшие результаты, этим ярче демонстрировать особенность работы в нашей области. В лекциях я ограничился передачей нашего фактического материала, почти совершенно не касаясь литературы предмета. Полная литературная обработка темы чрезвычайно увеличила бы мою работу, и без того нелегкую. А, кроме того, мне хотелось цельнее изобразить предмет в том виде, в каком он мне представлялся. Иначе мне пришлось бы встретиться с другими точками зрения, с другой постановкой вопросов, с сопоставлением противоположных фактов, с защитодних толкований, критикой других и т. д. Конечно, в нашем материале немало повторений фактов других авторов, но вопрос о приоритете не интересовал нас, глубоко убежденных в том, что в этой области достаточно возможности и простора для инициативы всех, к ней прикосновенных. Вместе с тем мы горячо верили в чрезвычайное торжество исследования при расширенной другими участниками исследования программе его, при других точках зрения, при другой постановке вопросов, при другой изобретательности в варьировании опытов и т. д. Мне остается исполнить долг благодарности. Сердечное спасибо всем моим сотрудникам, соединившим их добросовестный труд с моим трудом в нашем общем деле. Если я возбуждал, направлял и концентрировал нашу общую работу, то в свою очередь сам постоянно находился под влиянием наблюдательности и идейности моих сотрудников. В области мысли, при постоянном умственном общении, едва ли можно точно разграничить, что принадлежит одному и что другому. Зато каждый имеет удовлетворение и радость сознавать свое участие в общем результате. 12 июля 1926 годаПредисловие ко второму изданию
Первое издание этой книги разошлось неожиданно быстро. Но я мог бы уже, на основании продолжавшейся деятельности моих лабораторий, сделать в новом издании некоторые дополнения и изменения. К сожалению, постигшая меня болезнь сейчас мешает этому. Остается надеяться, что в третье, издании, если таковое осуществится, я буду иметь возможность привести содержание книги в полное соответствие с нашим лабораторным материалом. В этом издании сделаны только некоторые технические улучшения, исправлены немногие описки и опечатки и дополнен список опубликованных работ моих сотрудников. Май 1927 года даПредисловие к третьему изданию
Это третье издание моих «Лекций о работе больших полушарий головного мозга» есть перепечатка первых двух изданий (1926 [1] и 1927 гг.) без изменений и дополнений. Таким образом оно очень отстает от нашего лабораторного материала, чрезвычайно разросшегося с того времени. Тем не менее предлежащая книга имеет законное право на свое появление. Это есть основное и впервые систематизированное изложение наших фактов, отнимающее более чем три четверти срока всей доселе нашей работы, относящейся до физиологии и патологии высшей нервной деятельности. Все остальное, что собрано нами за восемь последних лет, может быть хорошо понято и зафиксировано в памяти лишь в системе этой книги. Сами новейшие факты и касающиеся их толкования необходимо искать в другой моей книге -- «Двадцатилетний опыт объективного изучения высшей нервной деятельности (поведения) животных". Таким образом обе эти книги тесно связаны друг с другом. Новое предстоящее издание «Двадцатилетнего опыта» познакомит читателя с достижениями моих лабораторий, можно сказать, до последних дней, но познакомит в очень кратком, так сказать, бездокументальном и вместе с тем в отрывочном изложении. Синтез этих книг, т. е. мое новое систематическое изложение всего нашего материала в виде одной книги, составит большую работу, осуществление которой я ставлю себе моей последней научной задачей. Эта работа займет у меня не один год. Если бы только судьба была благосклонна позволить мне исполнить этот мой важный жизненный долг при достаточной сохранности сил в моем возрасте! Академик Иван Павлов ноябрь 1935 г. Ленинград,Лекция первая. Обоснование и история принципиальной методики исследования работы больших полушарий. Понятие рефлекса. Разнообразие рефлексов. Сигнальная деятельность больших полушарии как самая общая физиологическая характеристика их
Мм. гг.! Нельзя не быть пораженным сопоставлением следующих фактов. Большие полушария, этот высший отдел центральной нервной системы, представляют собой довольно внушительную величину. Затем, эта масса - чрезвычайно сложной конструкции: она состоит из миллиарда (у человека из миллиардов) клеточек, т. е. центров, очагов нервной деятельности. Эти клеточки разных величин, форм и расположений соединены между собой бесчисленными разветвлениями их отростков. При такой сложной конструкции больших полушарий естественно предполагать, что они обладают и грандиозно сложной функцией. Следовательно, казалось бы, что для физиолога здесь открывается бесконечно широкое поле исследования. Это - во-первых. Во-вторых, представьте себе собаку, этого спутника и друга человека еще с доисторических времен, в ее разнообразных жизненных ролях: охотничьей, сторожевой и т. д. Мы знаем, что это ее сложное поведение, ее высшая нервная деятельность (ибо кто будет спорить, что это не высшая нервная деятельность) главнейшим образом связана с большими полушариями. Если мы удаляем (Гольц (Goltz) и за ним другие) у собаки большие полушария, то она не годится не только для тех ролей, о которых я только что упомянул, но не годится и для себя самой. Она глубокий инвалид и без посторонней помощи обречена на смерть. Значит, с одной стороны, по строению, с другой - по функции можно себе представить, какая большая физиологическая работа выпадает на долю больших полушарий. A человек? Разве вся его высшая деятельность не приурочена к нормальному строению и отправлению больших полушарий?! Как только сложная конструкция его полушарий так или иначе ломается, нарушается, человек тоже делается инвалидом, он не может свободно и равноправно жить среди близких и должен быть изолирован. И вот, в изумительном контрасте с этим необозримым объектом деятельности больших полушарий стоит содержание настоящей физиологии этих полушарий. До семидесятого года прошлого столетия не было даже никакой физиологии больших полушарий; они были для физиолога чем-то недоступным. Только в этом году впервые Фрич и Гитциг (Fritsch и Hitzig) с успехом применили к их изучению обычные физиологические методы раздражения и разрушения. При раздражении определенных мест коры больших полушарий регулярно наступали сокращения в определенных группах скелетных мышц (двигательная область коры). При удалении этих мест происходят известные нарушения в нормальной деятельности соответствующих групп мышц. Затем, вскоре после этого было показано Г. Мунком, Ферье (H. Munk, Ferrier) и др., что и другие отделы коры полушарий, которые оставались как будто искусственно нераздражимыми, тоже функционально обособлены. Удаление, экстирпация этих отделов ведет к определенным дефектам в деятельности некоторых рецепторных органов: глаза, уха и кожи. Эти факты подвергались и подвергаются энергичнейшей разработке со стороны многочисленных авторов вплоть до настоящего времени. Предмет уточнен и обогащен подробностями, специально в отношении двигательной области, и нашел даже важное практическое применение в медицине, но он доселе вращается главным образом все около первоначально намеченных пунктов. А существенное состоит в том, что все высшее и сложное поведение животного, приуроченное к большим полушариям, как показал это позднейший и выше уже мной упомянутый факт Гольца c удалением больших полушарий у собаки, осталось почти не затронутым этими исследованиями и не вошло в программу очередной физиологической работы. Что нам объясняют в поведении высших животных имеющиеся сейчас у физиологов факты относительно больших полушарий? I де общая схема высшей нервной деятельности? Где общие правила этой деятельности? Перед этими законнейшими вопросами современные физиологи стоят поистине с пустыми руками. Почему же объект так сложен конструктивно, так обилен функциями, а рядом с этим исследование его для физиолога уперлось как бы в угол, а не представляется почти безграничным, как можно было бы ожидать?


 Какая же этому причина? Причина есть и очень ясная, и заключается она в том, что та деятельность, которая производится большими полушариями, рассматривается не с той точки зрения, с которой рассматривается деятельность остальных органов тела и даже других частей центральной нервной системы. Эта деятельность больших полушарий получила название особой, психической деятельности, как мы чувствуем, воспринимаем ее в себе и как она предполагается у животных по аналогии с нами самими. Отсюда положение физиолого получилось в высшей степени своеобразным и затруднительным. С одной стороны, изучение деятельности полушарий, подобно другим частям организма, - как будто его дело, а с другой - выходит, что это есть предмет особой науки, психологии. Как же быть физиологу? Может быть, вопрос надо решитак, что физиолог должен запастись психологическими методами, знаниями и затем уже приступить к изучению деятельности больших полушарий? Но здесь есть существенное осложнение. Понятно, что физиологии постоянно в ее анализе жизни приходится опираться на более точные, совершенные науки: на механику, физику и химию. Но в этом случае - совсем другое. Теперь пришлось бы основываться на науке, которая своим совершенством сравнительно с физиологией похвалиться не может. Существовал даже еще недавно спор, можно ли вообще психологию считать естественной наукой и даже наукой вообще. Не входя в существо дела, я приведу лишь грубые внешние факты, хотя, как мне кажется, все же убедительные. Сами психологи не считают свою науку точной наукой. Выдающийся американский психолог Джемс (James) еще недавно называл психологию не наукой, а только «надеждой на науку». А вот еще более внушительное заявление, исходящее от Вундта (Wundt), который, будучи прежде физиологом, стал затем знаменитым психологом и философом и явился даже основателем так называемой экспериментальной психологии. Перед войной, в 1913 г., в Германии поднялся вопрос об отделении в университетах психологии от философии, т. е. об учреждении двух кафедр вместо прежней одной. Вундт оказался противником этого отделения и, между прочим, на том основании, что по психологии нельзя составить общеобязательной программы для экзамена, так как у каждого профессора своя особая психология. Не ясно ли, что психология еще не дошла до степени точной науки?
Но раз так, тогда для физиолога нет расчета обращаться к психологии. Имея в виду развитие естествознания, естественно ждать, что не психология должна помогать физиологии больших полушарий, а наоборот - физиологическое изучение этого органа у животных должно лечь в основание точного научного анализа субъективного мира человека. Следовательно, физиолог должен идти своим путем. И этот путь намечен уже давно. Считая деятельность животных, в противоположность человеческой, машинообразной, Декарт (Descartes) триста лет тому назад установил понятие рефлекса как основного акта нервной системы. Та или другая деятельность организма есть закономерный ответ на тот или другой внешний агент, причем эта связь деятельного органа с данным агентом, как причины со следствием, устанавливается при помощи определенного нервного пути. Таким образом изучение деятельности нервной системы животных было поставлено на твердую естественно-научную почву. В восемнадцатом, девятнадцатом и двадцатом столетиях физиологи фактически и подробно использовали идею рефлекса, но только на низших отделах центральной нервной системы, однако все выше и выше поднимаясь по отделам ее, пока, наконец, после классических работ Шеррингтона (Sherrington) над спинномозговыми рефлексами продолжатель его Магнус (Magnus) не доказал рефлекторного характера всех основных актов локомоторной деятельности. Таким образом идея рефлекса с полным экспериментальным оправданием была приложена к центральной нервной системе почти вплоть до больших полушарий. Можно надеяться, что и еще более сложные акты организма, куда элементом входят основные локомоторные рефлексы, - акты, обозначаемые пока психологическими терминами гнева, страха, игры и т. п., - будут скоро приурочены к простой рефлекторной деятельности части головного мозга, лежащей прямо под большими полушариями.
Смелый шаг в приложении идеи рефлекса к большим полушариям не только животных, но и человека сделал, стоя на почве современной ему физиологии нервной системы, русский физиолог И. М. Сеченов. В брошюре, появившейся на русском языке в 1863 г. под заглавием «Рефлексы головного мозга», он пытался представить деятельность больших полушарий как деятельность рефлекторную, т. е. ее детерминизировать. Мысли для него были рефлексы с задержанным эффекторным концом, а аффекты усиленные рефлексы с обширной иррадиацией раздражения. Ту же попытку в наше время повторил Рише (Ch. Richet), устанавливая понятие психического рефлекса, в котором реакция на данный раздражитель определялась комбинацией его со следами в больших полушариях прежних раздражений. Но и вообще у физиологов последнего времени высшая нервная деятельность, приуроченная к большим полушариям, характеризовалась связиванием наличных раздражений со следами старых (ассоциативная память у Ж. Леба (J. Loeb), обучаемость, пользование опытом - у других физиологов). Но все это было только теоризирование. Назревала потребность перехода к экспериментальному анализу предмета, и притом с объективной внешней стороны, как во всем остальном естествознании. Этот переход обусловила недавно народившаяся сравнительная физиология, возникшая сама как один из результатов влияния эволюционной теории. Обратившись ко всему животному миру, физиология, имея дело с низшими представителями, волей-неволей должна была отказаться от антропоморфической точки зрения в отношении их и сосредоточить научное внимание только на констатировании отношения между внешними влияниями, падавшими на животное, и ответной внешней деятельностью его, его движением. Отсюда - учение о животном тропизме M. Леба; отсюда - предложение объективной терминологии для обозначения реакций животных Бера, Бета и Икскюля (Beer, Bethe, Uexkull); отсюда, наконец, работы зоологов над низшими представителями животного мира, проводимые чисто объективно, только посредством сопоставления внешних влияний на животное с его ответной внешней деятельностью, например классическая работа Дженнингса (Jennings) и др.
Под влиянием этого нового течения в биологии и в силу специально делового склада американского ума возникает у американских психологов, также обратившихся к сравнительной психологии, тенденция подвергать экспериментальному анализу внешнюю деятельность животных при различных условиях, в которые они нарочно ставятся. Исходным пунктом систематических исследований такого рода справедливо поставить работу Торндайка (Thorndike) под заглавием «Animal Intelligence» (1898). При этих исследованиях животное помещалось в ящик, вне которого стояла видимая животному пища. Мивотное естественно стремилось к еде, а для этого должно было открыть дверцу ящика, которая в различных опытах запиралась разно. Цифры и составленные из них кривые показывали, как скоро и каким порядком достигалось решение задачи животным. Весь процесс понимался как образование ассоциации, связи между зрительным и осязательным раздражением и двигательным актом. Этим способом и разными вариациями его затемногочисленными авторами изучались разные вопросы, относящиеся до ассоциационной способности разнообразных животных. А почти одновременно с работой Торндайка, не зная о ней, я тоже был приведен к мысли стать в такое же отношение к предмету под влиянием одного лабораторного эпизода.
Изучая подробно деятельность пищеварительных желез, я должен был заняться и так называемым психическим возбуждением желез. Пробуя с одним из моих сотрудников анализировать этот факт глубже, сначала побщепринятому шаблону, т. е. психологически, соображаясь с тем, что животное могло думать и чувствовать при этом, я натолкнулся на необычайное в лаборатории событие. Я не мог сговориться со своим сотрудником; каждый из нас оставался при своем мнении, не имея возможности убедить другого определенными опытами. Это решительно восстановило меня против психологического обсуждения предмета, и я надумал исследовать предмет чисто объективно, с внешней стороны, т. е. точно отмечая, какое раздражение на животное падает в данный момент, и следя за тем, что животное в ответ на это раздражение проявляет в виде движении или (как это было в моем случае) в виде секреции.
Это было началом исследования, которое теперь продолжается уже двадцать пять лет и в котором участвовали многочисленные, мне дорогие сотрудники, соединившие их мысли и руки с моими мыслями и руками. Мы, конечно, переживали разные стадии; предмет только очень постепенно расширялся и углублялся. Сначала мы имели отдельные факты; теперь же материала уже накопилось столько, что он может быть как первая проба предложен в систематизированном до известной степени виде. Сейчас я в состоянии изложить вам такое физиологическое учение о работе больших полушарий, которое во всяком случае ближе подходит к действительной, конструктивной и функциональной, сложности этого органа, чем то, которое состояло до сих пор только из немногих совершенно отрывочных, хотя и большой важности, фактов современной физиологии.
Таким образом на этом новом пути строго объективного изучения высшей нервной деятельности работают главным образом мои лаборатории (это обнимает работу сотни моих сотрудников) и затем американские психологи. Что касается других физиологических лабораторий, то пока к этому предмету обратились только немногие, в более позднее время, и их исследование не вышло большей частью за пределы первой ориентировки в предмете. Между нами и американцами до сих пор существует, однако, следующая значительная разница. Раз там объективное изучение ведется психологами, то, хотя психологи и занимаются изучением чисто внешних фактов, тем не менее, что касается постановки задач, анализа и формулировки результатов, они думают большей частью психологически. Поэтому работы их не носят чисто физиологического характера, за исключением группы «бихевиористов». Мы же, выйдя из физиологии, все время строго придерживаемся физиологической точки зрения и весь предмет исследуем и систематизируем только физиологически.
Теперь перехожу к изложению нашего материала, предварительно остановившись на понятиях рефлекса вообще, рефлексов в физиологии и так называемых инстинктов.
Основным исходным понятием нас является декартовское понятие, понятие рефлекса. Конечно, оно вполне научно, так как явление, им обозначаемое, строго детерминизируется. Это значит, что в тот или другой рецепторный нервный прибор ударяет тот или другой агент внешнего мира или внутреннего мира организма. Этот удар трансформируется нервный процесс, в явление нервного возбуждения. Возбуждение по нервным волокнам, как проводам, бежит в центральную нервную систему и оттуда, благодаря установленным связям, по другим проводам приносится к рабочему органу, трансформируясь в свою очередь в специфический процесс клеток этого органа. Таким образом тот или другой агент закономерно связывается с той или другой деятельностью организма, как причина со следствием.
Совершенно очевидно, что вся деятельность организма должна быть закономерна. Если бы животное не было, употребляя биологический термин, точно приспособлено к внешнему миру, то оно скоро или медленно переставало бы существовать. Если бы животное, вместо того чтобы направляться к еде, отстранялось от нее, вместо того чтобы бежать от огня, кидалось в огонь и т. д. и т. д., оно было бы так или иначе разрушено. [1] Оно так должно реагировать на внешний мир, чтобы всей ответной деятельностью его было обеспечено его существование. То же самое окажется, если представлять себе жизнь в терминах механики, физики и химии. Каждая материальная система до тех пор может существовать как данная отдельность, пока ее внутренние силы притяжения, сцепления и т. д. уравновешиваются с внешними влияниями, среди которых она находится. Это относится ко всякому простому камню, как и к сложнейшему химическому веществу. Точно то же надо представлять себе и относительно организма. Как определенная замкнутая вещественная система он может существовать только до тех пор, пока он каждый момент уравновешивается с окружающими условиями. Как только это уравновешивание серьезно нарушается, он перестает существовать как данная система. Рефлексы суть элементы этого постоянного приспособления или постоянного уравновешивания. Физиологи изучили и постоянно изучают массу рефлексов, этих закономерно и машинообразно протекающих реакций организма и вместе с тем готовых со дня рождения, прирожденных, т. е. обусловленных организацией данной нервной системы. Рефлексы, как и приводы машин, сделанных человеческими руками, - двух родов: положительные и отрицательные, или задерживающие, тормозные, иначе сказать - то вызывающие определенную деятельность, то ее прекращающие. Конечно, изучение этих рефлексов, как ни давно оно в руках физиологов, еще очень, очень далеко от законченности. Открываются все новые рефлексы; рецепторные аппараты, принимающие удар агентов, как внешних, так и в особенности внутренних, еще в многочисленных случаях, что касается их свойств, остаются совершенно неисследованными; пути проведения нервного возбуждения в центральной нервной системе часто мало известны или совсем не установлены; центральный процесс задерживающих рефлексов, если исключить рефлексы на задерживающие эфферентные нервы, совершенно темен; так же очень немного ясного в соединении и взаимодействии различных рефлексов. Но тем не менее физиологи все более и более углубляются в механизм этой машинообразной деятельности организма и имеют все основания надеяться изучить ее рано или поздно с исчерпывающей полнотой, чтобы вполне управлять ею.
К этим обычным рефлексам, давнему объекту исследования физиологов в их лабораториях и касающимся главным образом деятельности отдельных органов, примыкают еще прирожденные реакции, также при посредстве нервной системы происходящие закономерно, т. е. строго приуроченные к определенным условиям. Это - реакции разнообразных животных, касающиеся деятельности целого организма под видом общего поведения животных и обозначаемые особым термином «инстинктов». Так как относительно этих реакции все еще нет полного согласия в отношении существенной однородности их с рефлексами, то приходится несколько больше остановиться на этом пункте.
Первой мыслью, что эти реакции тоже рефлексы, физиология обязана английскому философу Герберту Спенсеру (Herbert Spencer). А затем зоологи, физиологи и сравнительные психологи представили немало точных доказательств, что это действительно так. Сейчас я приведу в системе доводы за то, что нет ни одной существенной черты, отличающей рефлексы от инстинктов. Прежде всего есть множество совершенно незаметных переходов от обычных рефлексов к инстинктам. Возьмем цыпленка, который проделывает движение клевания (сейчас же после того, как он вышел из скорлупы) на всякое раздражение, попадающее ему в глаза, - от маленького предмета до пятна на плоскости, по которой он ходит. Чем это отличается, например, от отклонения головы и смыкания век, когда около глаза промелькнет близко какой-нибудь предмет? Про последнее скажут: оборонительный рефлекс, первое назовут пищевым инстинктом. А ведь в случае клевания, если оно вызвано пятном, все только наклонением головы и движением клюва и ограничивается.
Далее обращалось внимание на большую сложность инстинктов сравнительно с рефлексами. Но есть и чрезвычайно сложные рефлексы, которые, однако, никто инстинктами не называет. Возьмем простой рвотный акт. Он чрезвычайно сложен, в нем принимает экстренное согласованное участие огромное количество различных мускулов, и поперечнополосатых и гладких, обычно действующих при других функциях организма и раскинутых на большом протяжении, а также много различных секреции, опять же в другое время участвующих в других деятельностях организма.
Следующий пункт различия видели в длинном ряде последовательных инстинктивных действий сравнительно с одноэтажностью, так сказать, рефлекса. Возьмем для примера постройку незда или вообще жилища животных. Здесь имеется, конечно, длинная цепь действий: отыскивание и принесение материала на место, прилаживание его и укрепление. Если понимать это как рефлекс, то надо допустить, что конец одного рефлекса есть возбудитель следующего, т. е. что это цепные рефлексы. Но этот цепной характер актов вовсе не представляет исключительного свойства инстинктов. Мы знаем много и рефлексов, которые соединяются также в цепи. Возьмем следующий случай. Мы раздражаем какой-нибудь афферентный нерв, например n. ischiadicus; получается рефлекторное повышение кровяного давления. Это - первый рефлекс. Высокое давление внутри левого желудочка и начальной части аорты является возбудителем следующего рефлекса; оно раздражает концы п. depressores cordis, который вызывает депрессорный рефлекс, умеряющий эффект первого рефлекса. Возьмем еще новейший цепной рефлекс, установленный Магнусом. Кошка и без больших полушарий, брошенная с высоты, в большинстве случаев станет на ноги. Каким образом это достигается? Изменение в пространственном положении отолитового прибора уха вызывает определенное рефлекторное сокращение шейных мышц, устанавливающих голову животного в нормальное положение по отношению к горизонту. Это - первый рефлекс. Конец его - сокращение определенных шейных мышц и вообще постановка шеи - есть возбудитель другого рефлекса на определенные мышцы туловища и конечностей, дающего в окончательном результате правильное, стоячее положение животного.
Потом имелась в виду следующая якобы разница между рефлексами и инстинктами. Инстинкты часто зависят от определенных внутренних состояний, условий организма. Например постройка гнезда производится животным только тогда, когда оно собирается произвести новое поколение. Или более простой случай. Если животное сыто, оно не будет стремиться к пище, продолжать еду, а прекратит ее. То же относится и к половому инстинкту, связанному как с возрастным состоянием организма, так и с состоянием половых желез. Вообще здесь большую роль играют гормоны, продукты желез с внутренней секрецией. Но и это тоже не является исключительной принадлежностью инстинктов. Как интенсивность, так и наличие или отсутствие рефлексов прямо зависит от состояния возбудимости рефлекторных центров, а оно в свою очередь находится в постоянной зависимости от химических и физических свойств крови (автоматическое раздражение центров) и от взаимодействия разных рефлексов друг от друга.
Наконец иногда придается значение тому, что рефлексы относятся к деятельности отдельных органов, а инстинкты захватывают весь организм, т. е. собственно всю скелетно-мышечную систему. Но ведь, благодаря работам Магнуса и Клейна (Magnus und de Kleyn), мы знаем, что и стояние, и ходьба, и вообще уравновешивание тела в пространстве - рефлексы.
Таким образом как рефлексы, так и инстинкты - закономерные реакции организма на определенные агенты, и потому нет надобности обозначать их разными словами. Имеет преимущество слово «рефлекс», потому что ему с самого начала придан строго научный смысл.
Совокупность этих рефлексов составляет основной фонд нервной деятельности как человека, так животных. И поэтому, конечно, представляется делом огромной важности полное изучение всех этих основных нервных реакций организма. К сожалению, этого в настоящее время далеко нет, как это указано уже выше и как это надо в особенности подчеркнуть в отношении тех рефлексов, которые назывались инстинктами. Наше знание об этих инстинктах очень ограничено и отрывочно. Имеется лишь грубая группировка их: пищевой, самоохранетельный, половой, родительский и социальный. Но почти в каждой группе их заключается часто очень большое число отдельных членов, о существовании которых мы или совсем не догадываемся, или мешаем их с другими, или же по крайней мере не оцениваем в полной мере их жизненного значения. До какой степени этот предмет еще не полон, сколько в нем прорех, я могу иллюстрировать собственным примером.
П При нашей работе, о которой я буду сообщать, мы одно время стали в большой тупик, не понимая, что делается с нашим животным. Для опытов нам попалась собака, очевидно очень культурная, которая быстро вошла со всеми нами в дружеские отношения. Этой собаке была предложена, казалось, нетрудная задача. Ее ставили на столе в станок, ограничивая лишь ее движения мягкими петлями на ногах (к чему она сначала отнеслась совершенно спокойно), и затем на этом столе с ней ничего другого не делали, как только через промежутки в несколько минут повторно подкармливали. Сперва она мирно стояла и охотно ела, но чем больше она стояла, тем больше она возбуждалась: начиная воевать с обстановкой, она рвалась, царапала пол, грызла стойку станка, и т. д., причем от этой безустанной мышечной работы у нее начиналась одышка с постоянным слюнотечением, она делалась совершенно негодной для нашей работы, так как это продолжалось многие недели, все ухудшаясь. И мы долгое время были в недоумении: что бы это было? Мы проконтролировали много предположений о возможных причинах такого поведения, и, хотя мы имели уже достаточно знания о собаках, все оставалось без успеха, пока мы не напали на мысль, что это очень простая вещь, что это рефлекс свободы, т. е. что эта собака не переносила ограничения своих движений. Мы победили этот рефлекс другим рефлексом - пищевым. Мы стали давать собаке всю ее ежедневную порцию только в станке. Сначала она ела мало и порядочно захудала, но затем начала есть все больше, пока не стала съедать всю порцию и вместе с тем во время опытов сделалась спокойной; рефлекс свободы был задержан. Ясно, что рефлекс свободы - один из чрезвычайно важных рефлексов, или, общее сказать, реакций всякого живого существа. А рефлекс этот упоминается, но не всегда, он как бы не регламентирован окончательно. У Джемса его нет даже в числе специально человеческих рефлексов (инстинктов). Если бы у животного не было рефлекторного протеста борьбы против ограничения его движении, тогда стоило бы животному встретиться с пустым препятствием, и та или другая важная деятельность его оставалась бы неосуществленной. Мы знаем, что у некоторых животных этот рефлекс свободы так интенсивен, что, лишенные свободы, они отказываются от пищи, хиреют и умирают.
Возьмите другой пример. Едва ли достаточно оценивается рефлекс, который можно было бы назвать исследовательским рефлексом или, как я его навываю, рефлекс «что такое»», тоже один из фундаментальных рефлексов. И мы и животное при малейшем колебании окружающей среды устанавливаем соответствующий рецепторный аппарат по направлению к агенту этого колебания. Биологический смысл этого рефлекса огромен. Если бы у животного не было этой реакции, то жизнь его каждую минуту, можно сказать, висела бы на волоске. А у нас этот рефлекс идет чрезвычайно далеко, проявляясь, наконец, в виде той любознательности, которая создает науку, дающую и обещающую нам высочайшую, безграничную ориентировку в окружающем мире. Еще менее расчленен и оценен отдел отрицательных, задерживающих рефлексов (инстинктов) при всяких сильных, а также при необычных, хотя бы и слабых, раздражениях. Сюда, конечно, относится, между прочим, и так называемый животный гипнотизм.
Итак, основные нервные реакции как животного, так и человека прирождены в виде рефлексов. И я еще раз повторяю, что в высшей степени важно иметь полный список и надлежащую систематизацию этих рефлексов, потому что вся остальная нервная деятельность организма надстраивается, как увидим дальше, на фундаменте этих рефлексов.
Но, хотя описанные рефлексы представляют собой основное условие целости организма среди окружающей природы, однако их одних еще недостаточно для длительного, прочного и совершенного существования организма. Это доказывает опыт с собакой, у которой вырезаются большие полушария. У нее, не говоря o внутренних рефлексах, остаются фундаментальные внешние рефлексы. Она к еде стремится. От разрушительных раздражений отстраняется. У нее есть исследовательский рефлекс: она при звуке поднимает уши и голову. У нее есть и рефлекс свободы: при захватывании ее она сильно сопротивляется. И, однако, она инвалид, она не может существовать предоставленная сама себе. Значит, в теперешней нервной деятельности ее чего-то и очень важного нехватает. Чего же? Нельзя не заметить, что у этой собаки агенты, которые вызывают рефлексы, стали очень малочисленными, пространственно очень близкихи, очень элементарными и очень общими, недифференцированными, и поэтому при посредстве их уравновешивание этого высшего организма с окружающей средой в широком районе его жизни стало очень упрощенным, слишком ограниченным, явно недостаточным.
Возьмем самый простой пример, с которого мы начали свои исследования. Когда нормальному животному попадает в рот еда или что-нибудь из отвергаемых веществ, тогда на них изливается слюна, которая увлажняет, растворяет и химически изменяет пищевые вещества, а отвергаемые удаляет и отмывает от них рот. Это рефлекс, обусловленный физическими и химическими свойствами всех этих веществ при соприкосновении их со слизистой оболочкой рта. Но, кроме этого, ту же секреторную реакцию вызывают те же вещества, когда они находятся перед собакой вдали и могут действовать на собаку только через глаз и нос. Больше того, та же реакция появляется и тогда, когда перед собакой стоит лишь посуда, из которой раньше попадали эти вещества ей в рот. Еще больше того, то же делает вид человека, который обыкновенно приносил эти вещества, даже звуки его шагов из другой комнаты. И все эти многочисленные, отдаленные, сложные и тонко специализированные раздражители теряют навсегда свое действие, лишь только собака лишается больших полушарий; остаются действующими только физические и химические свойства веществ при их соприкосновении со слизистой оболочкой рта. А машинная выгода утерянных раздражителей в нормальном случае очень значительна. Сухая пища сейчас же встречается с большим количеством нужной ей жидкости; ответгаемые вещества, часто разрушающие оболочку рта, слоем наличной слюны оттесняются от нее и быстро разжижаются и т. д. Во сколько же значение тех же раздражителей повышается, когда ими приводится в действие двигательный компонент пищевого рефлекса, т. е. осуществляется добывание пищи!
Возьмем еще важный случай оборонительного рефлекса. Сильный зверь пользуется как пищей маленьким, слабеньким животным. Последнее должно перестать существовать, если оно начинает оборону лишь тогда, когда в par прикоснется к нему своими зубами и когтями. И другое дело, если оборонительная реакция возникнет при одном виде врага еще издали, при его звуках и т. д. Тогда слабенькое животное будет иметь возможность убежать, скрыться, т. е. уцелеть.
Как же характеризовать вообще различие в отношении животного к внешнему миру в норме и без больших полушарий? Каков общий механизм этих отношений? Каков принцип этого различия?
Нетрудно видеть, что в норме реакции организма вызываются не только существенными для организма агентами внешнего мира, т. e. непосредственно благоприятствующими организму или его разрушающими, но и прямо бесчисленным количеством других агентов, только сигнализирующих первые, как это очевидно в вышеприведенных примерах. Ведь вид и звуки сильного зверя не разрушают маленькое животное, но это делают его зубы и когти. Однако сигнализирующие, или, употребляя термин Шеррингтона, дистантные раздражители, хотя и в сравнительно ограниченном числе, имеют место и при тех рефлексах, о которых шла речь доселе. Существенный признак высшей нервной деятельности, которой мы будем заниматься и которая в высшем животном принадлежит, вероятно, исключительно большим полушариям, состоит не только в том, что здесь действуют бесчисленные сигнальные раздражители, но и в том существенно, что они при определенных условиях меняют свое физиологическое действие.
В вышеприведенном примере со слюнной реакцией действует то та, то другая посуда, то тот, то другой человек, в строгой зависимости от того, из какой посуды на глазах собаки попадали пищевые и отвергаемые вещества в рот собаки и кто их приносил и давал или вводил собаке. Это, очевидно, уточняет еще более машинную деятельность организма, придает ей свойство еще большего совершенства. Окружающая животное среда так бесконечно сложна и находится в таком постоянном движении, что сложная замкнутая система организма, лишь тоже соответственно колеблющаяся, имеет шансы быть с ней уравновешенной.
Итак, основная и самая общая деятельность больших полушарий есть сигнальная, с бесчисленным количеством сигналов и с переменной сигнализацией.
Какая же этому причина? Причина есть и очень ясная, и заключается она в том, что та деятельность, которая производится большими полушариями, рассматривается не с той точки зрения, с которой рассматривается деятельность остальных органов тела и даже других частей центральной нервной системы. Эта деятельность больших полушарий получила название особой, психической деятельности, как мы чувствуем, воспринимаем ее в себе и как она предполагается у животных по аналогии с нами самими. Отсюда положение физиолого получилось в высшей степени своеобразным и затруднительным. С одной стороны, изучение деятельности полушарий, подобно другим частям организма, - как будто его дело, а с другой - выходит, что это есть предмет особой науки, психологии. Как же быть физиологу? Может быть, вопрос надо решитак, что физиолог должен запастись психологическими методами, знаниями и затем уже приступить к изучению деятельности больших полушарий? Но здесь есть существенное осложнение. Понятно, что физиологии постоянно в ее анализе жизни приходится опираться на более точные, совершенные науки: на механику, физику и химию. Но в этом случае - совсем другое. Теперь пришлось бы основываться на науке, которая своим совершенством сравнительно с физиологией похвалиться не может. Существовал даже еще недавно спор, можно ли вообще психологию считать естественной наукой и даже наукой вообще. Не входя в существо дела, я приведу лишь грубые внешние факты, хотя, как мне кажется, все же убедительные. Сами психологи не считают свою науку точной наукой. Выдающийся американский психолог Джемс (James) еще недавно называл психологию не наукой, а только «надеждой на науку». А вот еще более внушительное заявление, исходящее от Вундта (Wundt), который, будучи прежде физиологом, стал затем знаменитым психологом и философом и явился даже основателем так называемой экспериментальной психологии. Перед войной, в 1913 г., в Германии поднялся вопрос об отделении в университетах психологии от философии, т. е. об учреждении двух кафедр вместо прежней одной. Вундт оказался противником этого отделения и, между прочим, на том основании, что по психологии нельзя составить общеобязательной программы для экзамена, так как у каждого профессора своя особая психология. Не ясно ли, что психология еще не дошла до степени точной науки?
Но раз так, тогда для физиолога нет расчета обращаться к психологии. Имея в виду развитие естествознания, естественно ждать, что не психология должна помогать физиологии больших полушарий, а наоборот - физиологическое изучение этого органа у животных должно лечь в основание точного научного анализа субъективного мира человека. Следовательно, физиолог должен идти своим путем. И этот путь намечен уже давно. Считая деятельность животных, в противоположность человеческой, машинообразной, Декарт (Descartes) триста лет тому назад установил понятие рефлекса как основного акта нервной системы. Та или другая деятельность организма есть закономерный ответ на тот или другой внешний агент, причем эта связь деятельного органа с данным агентом, как причины со следствием, устанавливается при помощи определенного нервного пути. Таким образом изучение деятельности нервной системы животных было поставлено на твердую естественно-научную почву. В восемнадцатом, девятнадцатом и двадцатом столетиях физиологи фактически и подробно использовали идею рефлекса, но только на низших отделах центральной нервной системы, однако все выше и выше поднимаясь по отделам ее, пока, наконец, после классических работ Шеррингтона (Sherrington) над спинномозговыми рефлексами продолжатель его Магнус (Magnus) не доказал рефлекторного характера всех основных актов локомоторной деятельности. Таким образом идея рефлекса с полным экспериментальным оправданием была приложена к центральной нервной системе почти вплоть до больших полушарий. Можно надеяться, что и еще более сложные акты организма, куда элементом входят основные локомоторные рефлексы, - акты, обозначаемые пока психологическими терминами гнева, страха, игры и т. п., - будут скоро приурочены к простой рефлекторной деятельности части головного мозга, лежащей прямо под большими полушариями.
Смелый шаг в приложении идеи рефлекса к большим полушариям не только животных, но и человека сделал, стоя на почве современной ему физиологии нервной системы, русский физиолог И. М. Сеченов. В брошюре, появившейся на русском языке в 1863 г. под заглавием «Рефлексы головного мозга», он пытался представить деятельность больших полушарий как деятельность рефлекторную, т. е. ее детерминизировать. Мысли для него были рефлексы с задержанным эффекторным концом, а аффекты усиленные рефлексы с обширной иррадиацией раздражения. Ту же попытку в наше время повторил Рише (Ch. Richet), устанавливая понятие психического рефлекса, в котором реакция на данный раздражитель определялась комбинацией его со следами в больших полушариях прежних раздражений. Но и вообще у физиологов последнего времени высшая нервная деятельность, приуроченная к большим полушариям, характеризовалась связиванием наличных раздражений со следами старых (ассоциативная память у Ж. Леба (J. Loeb), обучаемость, пользование опытом - у других физиологов). Но все это было только теоризирование. Назревала потребность перехода к экспериментальному анализу предмета, и притом с объективной внешней стороны, как во всем остальном естествознании. Этот переход обусловила недавно народившаяся сравнительная физиология, возникшая сама как один из результатов влияния эволюционной теории. Обратившись ко всему животному миру, физиология, имея дело с низшими представителями, волей-неволей должна была отказаться от антропоморфической точки зрения в отношении их и сосредоточить научное внимание только на констатировании отношения между внешними влияниями, падавшими на животное, и ответной внешней деятельностью его, его движением. Отсюда - учение о животном тропизме M. Леба; отсюда - предложение объективной терминологии для обозначения реакций животных Бера, Бета и Икскюля (Beer, Bethe, Uexkull); отсюда, наконец, работы зоологов над низшими представителями животного мира, проводимые чисто объективно, только посредством сопоставления внешних влияний на животное с его ответной внешней деятельностью, например классическая работа Дженнингса (Jennings) и др.
Под влиянием этого нового течения в биологии и в силу специально делового склада американского ума возникает у американских психологов, также обратившихся к сравнительной психологии, тенденция подвергать экспериментальному анализу внешнюю деятельность животных при различных условиях, в которые они нарочно ставятся. Исходным пунктом систематических исследований такого рода справедливо поставить работу Торндайка (Thorndike) под заглавием «Animal Intelligence» (1898). При этих исследованиях животное помещалось в ящик, вне которого стояла видимая животному пища. Мивотное естественно стремилось к еде, а для этого должно было открыть дверцу ящика, которая в различных опытах запиралась разно. Цифры и составленные из них кривые показывали, как скоро и каким порядком достигалось решение задачи животным. Весь процесс понимался как образование ассоциации, связи между зрительным и осязательным раздражением и двигательным актом. Этим способом и разными вариациями его затемногочисленными авторами изучались разные вопросы, относящиеся до ассоциационной способности разнообразных животных. А почти одновременно с работой Торндайка, не зная о ней, я тоже был приведен к мысли стать в такое же отношение к предмету под влиянием одного лабораторного эпизода.
Изучая подробно деятельность пищеварительных желез, я должен был заняться и так называемым психическим возбуждением желез. Пробуя с одним из моих сотрудников анализировать этот факт глубже, сначала побщепринятому шаблону, т. е. психологически, соображаясь с тем, что животное могло думать и чувствовать при этом, я натолкнулся на необычайное в лаборатории событие. Я не мог сговориться со своим сотрудником; каждый из нас оставался при своем мнении, не имея возможности убедить другого определенными опытами. Это решительно восстановило меня против психологического обсуждения предмета, и я надумал исследовать предмет чисто объективно, с внешней стороны, т. е. точно отмечая, какое раздражение на животное падает в данный момент, и следя за тем, что животное в ответ на это раздражение проявляет в виде движении или (как это было в моем случае) в виде секреции.
Это было началом исследования, которое теперь продолжается уже двадцать пять лет и в котором участвовали многочисленные, мне дорогие сотрудники, соединившие их мысли и руки с моими мыслями и руками. Мы, конечно, переживали разные стадии; предмет только очень постепенно расширялся и углублялся. Сначала мы имели отдельные факты; теперь же материала уже накопилось столько, что он может быть как первая проба предложен в систематизированном до известной степени виде. Сейчас я в состоянии изложить вам такое физиологическое учение о работе больших полушарий, которое во всяком случае ближе подходит к действительной, конструктивной и функциональной, сложности этого органа, чем то, которое состояло до сих пор только из немногих совершенно отрывочных, хотя и большой важности, фактов современной физиологии.
Таким образом на этом новом пути строго объективного изучения высшей нервной деятельности работают главным образом мои лаборатории (это обнимает работу сотни моих сотрудников) и затем американские психологи. Что касается других физиологических лабораторий, то пока к этому предмету обратились только немногие, в более позднее время, и их исследование не вышло большей частью за пределы первой ориентировки в предмете. Между нами и американцами до сих пор существует, однако, следующая значительная разница. Раз там объективное изучение ведется психологами, то, хотя психологи и занимаются изучением чисто внешних фактов, тем не менее, что касается постановки задач, анализа и формулировки результатов, они думают большей частью психологически. Поэтому работы их не носят чисто физиологического характера, за исключением группы «бихевиористов». Мы же, выйдя из физиологии, все время строго придерживаемся физиологической точки зрения и весь предмет исследуем и систематизируем только физиологически.
Теперь перехожу к изложению нашего материала, предварительно остановившись на понятиях рефлекса вообще, рефлексов в физиологии и так называемых инстинктов.
Основным исходным понятием нас является декартовское понятие, понятие рефлекса. Конечно, оно вполне научно, так как явление, им обозначаемое, строго детерминизируется. Это значит, что в тот или другой рецепторный нервный прибор ударяет тот или другой агент внешнего мира или внутреннего мира организма. Этот удар трансформируется нервный процесс, в явление нервного возбуждения. Возбуждение по нервным волокнам, как проводам, бежит в центральную нервную систему и оттуда, благодаря установленным связям, по другим проводам приносится к рабочему органу, трансформируясь в свою очередь в специфический процесс клеток этого органа. Таким образом тот или другой агент закономерно связывается с той или другой деятельностью организма, как причина со следствием.
Совершенно очевидно, что вся деятельность организма должна быть закономерна. Если бы животное не было, употребляя биологический термин, точно приспособлено к внешнему миру, то оно скоро или медленно переставало бы существовать. Если бы животное, вместо того чтобы направляться к еде, отстранялось от нее, вместо того чтобы бежать от огня, кидалось в огонь и т. д. и т. д., оно было бы так или иначе разрушено. [1] Оно так должно реагировать на внешний мир, чтобы всей ответной деятельностью его было обеспечено его существование. То же самое окажется, если представлять себе жизнь в терминах механики, физики и химии. Каждая материальная система до тех пор может существовать как данная отдельность, пока ее внутренние силы притяжения, сцепления и т. д. уравновешиваются с внешними влияниями, среди которых она находится. Это относится ко всякому простому камню, как и к сложнейшему химическому веществу. Точно то же надо представлять себе и относительно организма. Как определенная замкнутая вещественная система он может существовать только до тех пор, пока он каждый момент уравновешивается с окружающими условиями. Как только это уравновешивание серьезно нарушается, он перестает существовать как данная система. Рефлексы суть элементы этого постоянного приспособления или постоянного уравновешивания. Физиологи изучили и постоянно изучают массу рефлексов, этих закономерно и машинообразно протекающих реакций организма и вместе с тем готовых со дня рождения, прирожденных, т. е. обусловленных организацией данной нервной системы. Рефлексы, как и приводы машин, сделанных человеческими руками, - двух родов: положительные и отрицательные, или задерживающие, тормозные, иначе сказать - то вызывающие определенную деятельность, то ее прекращающие. Конечно, изучение этих рефлексов, как ни давно оно в руках физиологов, еще очень, очень далеко от законченности. Открываются все новые рефлексы; рецепторные аппараты, принимающие удар агентов, как внешних, так и в особенности внутренних, еще в многочисленных случаях, что касается их свойств, остаются совершенно неисследованными; пути проведения нервного возбуждения в центральной нервной системе часто мало известны или совсем не установлены; центральный процесс задерживающих рефлексов, если исключить рефлексы на задерживающие эфферентные нервы, совершенно темен; так же очень немного ясного в соединении и взаимодействии различных рефлексов. Но тем не менее физиологи все более и более углубляются в механизм этой машинообразной деятельности организма и имеют все основания надеяться изучить ее рано или поздно с исчерпывающей полнотой, чтобы вполне управлять ею.
К этим обычным рефлексам, давнему объекту исследования физиологов в их лабораториях и касающимся главным образом деятельности отдельных органов, примыкают еще прирожденные реакции, также при посредстве нервной системы происходящие закономерно, т. е. строго приуроченные к определенным условиям. Это - реакции разнообразных животных, касающиеся деятельности целого организма под видом общего поведения животных и обозначаемые особым термином «инстинктов». Так как относительно этих реакции все еще нет полного согласия в отношении существенной однородности их с рефлексами, то приходится несколько больше остановиться на этом пункте.
Первой мыслью, что эти реакции тоже рефлексы, физиология обязана английскому философу Герберту Спенсеру (Herbert Spencer). А затем зоологи, физиологи и сравнительные психологи представили немало точных доказательств, что это действительно так. Сейчас я приведу в системе доводы за то, что нет ни одной существенной черты, отличающей рефлексы от инстинктов. Прежде всего есть множество совершенно незаметных переходов от обычных рефлексов к инстинктам. Возьмем цыпленка, который проделывает движение клевания (сейчас же после того, как он вышел из скорлупы) на всякое раздражение, попадающее ему в глаза, - от маленького предмета до пятна на плоскости, по которой он ходит. Чем это отличается, например, от отклонения головы и смыкания век, когда около глаза промелькнет близко какой-нибудь предмет? Про последнее скажут: оборонительный рефлекс, первое назовут пищевым инстинктом. А ведь в случае клевания, если оно вызвано пятном, все только наклонением головы и движением клюва и ограничивается.
Далее обращалось внимание на большую сложность инстинктов сравнительно с рефлексами. Но есть и чрезвычайно сложные рефлексы, которые, однако, никто инстинктами не называет. Возьмем простой рвотный акт. Он чрезвычайно сложен, в нем принимает экстренное согласованное участие огромное количество различных мускулов, и поперечнополосатых и гладких, обычно действующих при других функциях организма и раскинутых на большом протяжении, а также много различных секреции, опять же в другое время участвующих в других деятельностях организма.
Следующий пункт различия видели в длинном ряде последовательных инстинктивных действий сравнительно с одноэтажностью, так сказать, рефлекса. Возьмем для примера постройку незда или вообще жилища животных. Здесь имеется, конечно, длинная цепь действий: отыскивание и принесение материала на место, прилаживание его и укрепление. Если понимать это как рефлекс, то надо допустить, что конец одного рефлекса есть возбудитель следующего, т. е. что это цепные рефлексы. Но этот цепной характер актов вовсе не представляет исключительного свойства инстинктов. Мы знаем много и рефлексов, которые соединяются также в цепи. Возьмем следующий случай. Мы раздражаем какой-нибудь афферентный нерв, например n. ischiadicus; получается рефлекторное повышение кровяного давления. Это - первый рефлекс. Высокое давление внутри левого желудочка и начальной части аорты является возбудителем следующего рефлекса; оно раздражает концы п. depressores cordis, который вызывает депрессорный рефлекс, умеряющий эффект первого рефлекса. Возьмем еще новейший цепной рефлекс, установленный Магнусом. Кошка и без больших полушарий, брошенная с высоты, в большинстве случаев станет на ноги. Каким образом это достигается? Изменение в пространственном положении отолитового прибора уха вызывает определенное рефлекторное сокращение шейных мышц, устанавливающих голову животного в нормальное положение по отношению к горизонту. Это - первый рефлекс. Конец его - сокращение определенных шейных мышц и вообще постановка шеи - есть возбудитель другого рефлекса на определенные мышцы туловища и конечностей, дающего в окончательном результате правильное, стоячее положение животного.
Потом имелась в виду следующая якобы разница между рефлексами и инстинктами. Инстинкты часто зависят от определенных внутренних состояний, условий организма. Например постройка гнезда производится животным только тогда, когда оно собирается произвести новое поколение. Или более простой случай. Если животное сыто, оно не будет стремиться к пище, продолжать еду, а прекратит ее. То же относится и к половому инстинкту, связанному как с возрастным состоянием организма, так и с состоянием половых желез. Вообще здесь большую роль играют гормоны, продукты желез с внутренней секрецией. Но и это тоже не является исключительной принадлежностью инстинктов. Как интенсивность, так и наличие или отсутствие рефлексов прямо зависит от состояния возбудимости рефлекторных центров, а оно в свою очередь находится в постоянной зависимости от химических и физических свойств крови (автоматическое раздражение центров) и от взаимодействия разных рефлексов друг от друга.
Наконец иногда придается значение тому, что рефлексы относятся к деятельности отдельных органов, а инстинкты захватывают весь организм, т. е. собственно всю скелетно-мышечную систему. Но ведь, благодаря работам Магнуса и Клейна (Magnus und de Kleyn), мы знаем, что и стояние, и ходьба, и вообще уравновешивание тела в пространстве - рефлексы.
Таким образом как рефлексы, так и инстинкты - закономерные реакции организма на определенные агенты, и потому нет надобности обозначать их разными словами. Имеет преимущество слово «рефлекс», потому что ему с самого начала придан строго научный смысл.
Совокупность этих рефлексов составляет основной фонд нервной деятельности как человека, так животных. И поэтому, конечно, представляется делом огромной важности полное изучение всех этих основных нервных реакций организма. К сожалению, этого в настоящее время далеко нет, как это указано уже выше и как это надо в особенности подчеркнуть в отношении тех рефлексов, которые назывались инстинктами. Наше знание об этих инстинктах очень ограничено и отрывочно. Имеется лишь грубая группировка их: пищевой, самоохранетельный, половой, родительский и социальный. Но почти в каждой группе их заключается часто очень большое число отдельных членов, о существовании которых мы или совсем не догадываемся, или мешаем их с другими, или же по крайней мере не оцениваем в полной мере их жизненного значения. До какой степени этот предмет еще не полон, сколько в нем прорех, я могу иллюстрировать собственным примером.
П При нашей работе, о которой я буду сообщать, мы одно время стали в большой тупик, не понимая, что делается с нашим животным. Для опытов нам попалась собака, очевидно очень культурная, которая быстро вошла со всеми нами в дружеские отношения. Этой собаке была предложена, казалось, нетрудная задача. Ее ставили на столе в станок, ограничивая лишь ее движения мягкими петлями на ногах (к чему она сначала отнеслась совершенно спокойно), и затем на этом столе с ней ничего другого не делали, как только через промежутки в несколько минут повторно подкармливали. Сперва она мирно стояла и охотно ела, но чем больше она стояла, тем больше она возбуждалась: начиная воевать с обстановкой, она рвалась, царапала пол, грызла стойку станка, и т. д., причем от этой безустанной мышечной работы у нее начиналась одышка с постоянным слюнотечением, она делалась совершенно негодной для нашей работы, так как это продолжалось многие недели, все ухудшаясь. И мы долгое время были в недоумении: что бы это было? Мы проконтролировали много предположений о возможных причинах такого поведения, и, хотя мы имели уже достаточно знания о собаках, все оставалось без успеха, пока мы не напали на мысль, что это очень простая вещь, что это рефлекс свободы, т. е. что эта собака не переносила ограничения своих движений. Мы победили этот рефлекс другим рефлексом - пищевым. Мы стали давать собаке всю ее ежедневную порцию только в станке. Сначала она ела мало и порядочно захудала, но затем начала есть все больше, пока не стала съедать всю порцию и вместе с тем во время опытов сделалась спокойной; рефлекс свободы был задержан. Ясно, что рефлекс свободы - один из чрезвычайно важных рефлексов, или, общее сказать, реакций всякого живого существа. А рефлекс этот упоминается, но не всегда, он как бы не регламентирован окончательно. У Джемса его нет даже в числе специально человеческих рефлексов (инстинктов). Если бы у животного не было рефлекторного протеста борьбы против ограничения его движении, тогда стоило бы животному встретиться с пустым препятствием, и та или другая важная деятельность его оставалась бы неосуществленной. Мы знаем, что у некоторых животных этот рефлекс свободы так интенсивен, что, лишенные свободы, они отказываются от пищи, хиреют и умирают.
Возьмите другой пример. Едва ли достаточно оценивается рефлекс, который можно было бы назвать исследовательским рефлексом или, как я его навываю, рефлекс «что такое»», тоже один из фундаментальных рефлексов. И мы и животное при малейшем колебании окружающей среды устанавливаем соответствующий рецепторный аппарат по направлению к агенту этого колебания. Биологический смысл этого рефлекса огромен. Если бы у животного не было этой реакции, то жизнь его каждую минуту, можно сказать, висела бы на волоске. А у нас этот рефлекс идет чрезвычайно далеко, проявляясь, наконец, в виде той любознательности, которая создает науку, дающую и обещающую нам высочайшую, безграничную ориентировку в окружающем мире. Еще менее расчленен и оценен отдел отрицательных, задерживающих рефлексов (инстинктов) при всяких сильных, а также при необычных, хотя бы и слабых, раздражениях. Сюда, конечно, относится, между прочим, и так называемый животный гипнотизм.
Итак, основные нервные реакции как животного, так и человека прирождены в виде рефлексов. И я еще раз повторяю, что в высшей степени важно иметь полный список и надлежащую систематизацию этих рефлексов, потому что вся остальная нервная деятельность организма надстраивается, как увидим дальше, на фундаменте этих рефлексов.
Но, хотя описанные рефлексы представляют собой основное условие целости организма среди окружающей природы, однако их одних еще недостаточно для длительного, прочного и совершенного существования организма. Это доказывает опыт с собакой, у которой вырезаются большие полушария. У нее, не говоря o внутренних рефлексах, остаются фундаментальные внешние рефлексы. Она к еде стремится. От разрушительных раздражений отстраняется. У нее есть исследовательский рефлекс: она при звуке поднимает уши и голову. У нее есть и рефлекс свободы: при захватывании ее она сильно сопротивляется. И, однако, она инвалид, она не может существовать предоставленная сама себе. Значит, в теперешней нервной деятельности ее чего-то и очень важного нехватает. Чего же? Нельзя не заметить, что у этой собаки агенты, которые вызывают рефлексы, стали очень малочисленными, пространственно очень близкихи, очень элементарными и очень общими, недифференцированными, и поэтому при посредстве их уравновешивание этого высшего организма с окружающей средой в широком районе его жизни стало очень упрощенным, слишком ограниченным, явно недостаточным.
Возьмем самый простой пример, с которого мы начали свои исследования. Когда нормальному животному попадает в рот еда или что-нибудь из отвергаемых веществ, тогда на них изливается слюна, которая увлажняет, растворяет и химически изменяет пищевые вещества, а отвергаемые удаляет и отмывает от них рот. Это рефлекс, обусловленный физическими и химическими свойствами всех этих веществ при соприкосновении их со слизистой оболочкой рта. Но, кроме этого, ту же секреторную реакцию вызывают те же вещества, когда они находятся перед собакой вдали и могут действовать на собаку только через глаз и нос. Больше того, та же реакция появляется и тогда, когда перед собакой стоит лишь посуда, из которой раньше попадали эти вещества ей в рот. Еще больше того, то же делает вид человека, который обыкновенно приносил эти вещества, даже звуки его шагов из другой комнаты. И все эти многочисленные, отдаленные, сложные и тонко специализированные раздражители теряют навсегда свое действие, лишь только собака лишается больших полушарий; остаются действующими только физические и химические свойства веществ при их соприкосновении со слизистой оболочкой рта. А машинная выгода утерянных раздражителей в нормальном случае очень значительна. Сухая пища сейчас же встречается с большим количеством нужной ей жидкости; ответгаемые вещества, часто разрушающие оболочку рта, слоем наличной слюны оттесняются от нее и быстро разжижаются и т. д. Во сколько же значение тех же раздражителей повышается, когда ими приводится в действие двигательный компонент пищевого рефлекса, т. е. осуществляется добывание пищи!
Возьмем еще важный случай оборонительного рефлекса. Сильный зверь пользуется как пищей маленьким, слабеньким животным. Последнее должно перестать существовать, если оно начинает оборону лишь тогда, когда в par прикоснется к нему своими зубами и когтями. И другое дело, если оборонительная реакция возникнет при одном виде врага еще издали, при его звуках и т. д. Тогда слабенькое животное будет иметь возможность убежать, скрыться, т. е. уцелеть.
Как же характеризовать вообще различие в отношении животного к внешнему миру в норме и без больших полушарий? Каков общий механизм этих отношений? Каков принцип этого различия?
Нетрудно видеть, что в норме реакции организма вызываются не только существенными для организма агентами внешнего мира, т. e. непосредственно благоприятствующими организму или его разрушающими, но и прямо бесчисленным количеством других агентов, только сигнализирующих первые, как это очевидно в вышеприведенных примерах. Ведь вид и звуки сильного зверя не разрушают маленькое животное, но это делают его зубы и когти. Однако сигнализирующие, или, употребляя термин Шеррингтона, дистантные раздражители, хотя и в сравнительно ограниченном числе, имеют место и при тех рефлексах, о которых шла речь доселе. Существенный признак высшей нервной деятельности, которой мы будем заниматься и которая в высшем животном принадлежит, вероятно, исключительно большим полушариям, состоит не только в том, что здесь действуют бесчисленные сигнальные раздражители, но и в том существенно, что они при определенных условиях меняют свое физиологическое действие.
В вышеприведенном примере со слюнной реакцией действует то та, то другая посуда, то тот, то другой человек, в строгой зависимости от того, из какой посуды на глазах собаки попадали пищевые и отвергаемые вещества в рот собаки и кто их приносил и давал или вводил собаке. Это, очевидно, уточняет еще более машинную деятельность организма, придает ей свойство еще большего совершенства. Окружающая животное среда так бесконечно сложна и находится в таком постоянном движении, что сложная замкнутая система организма, лишь тоже соответственно колеблющаяся, имеет шансы быть с ней уравновешенной.
Итак, основная и самая общая деятельность больших полушарий есть сигнальная, с бесчисленным количеством сигналов и с переменной сигнализацией.
Лекция вторая. Техническая методика объективного исследования работы больших полушарии. Сигнализация есть рефлекс. Безусловный и условный рефлекс. Условия образования условных рефлексов
Мм. гг.! Прошлый раз я описал повод и основания, в силу которых нами было решено изучать всю нервную деятельность высших животных исключительно объективным способом, т. е. изучать с чисто внешней, фактической стороны, как это делается во всем естествознании, совершенно не обращаясь к фантастическим соображениям о том, что может переживать внутри себя собака, по аналогии с нами самими. Вместе с тем я сообщил вам, что с этой точки зрения вся нервная деятельность животного представилась нам, во-первых, в виде прирожденных рефлексов, т. е. в виде закономерных связей известных внешних агентов, действующих на организм, с определенными ответными деятельностями организма, причем было выяснено, что таких агентов относительно немного, они оказались вообще близкими и общего характера. Это, конечно, до известной степени обеспечивает существование организма, но далеко не достаточно (в особенности же для более или менее высших животных); так что, если мы лишим животное известной части нервной деятельности, то располагающее только прирожденными рефлексами животное, предоставленное себе, обречено на инвалидное существование, обречено на смерть. Ежедневная полная жизнь требует более детальных, специальных соотношений животного с окружающим миром. И это второе дальнейшее соотношение устанавливается только при помощи высшего отдела центральной нервной системы, больших полушарии, причем дело, ближе говоря, обстоит так, что множество всевозможных агентов природы сигнализируют собой, и притом временно и переменно, те основные относительно немногочисленные агенты, которые обусловливают врожденные рефлексы. И таким только образом достигается точное и тонкое уравновешивание организма с окружающим миром. Эту деятельность больших полушарий я назвал сигнальной деятельностью. Теперь я прежде всего должен остановиться на технической стороне нашей методики. Как же мы будем изучать сигнальную деятельность полушарий, на каком органе и какими приемами? Понятно, что для этого изучения можно было бы взять любой рефлекс, так как со всеми рефлексами связываются сигнальные раздражители. Но по историческим условиям нашей работы, как уже упомянуто, мы сосредоточились на двух рефлексах: пищевом и самом обычном оборонительном, когда собаке (нашему экспериментальному объекту) попадает в рот какое-либо из отвергаемых веществ. И это оказалось очень выгодным во многих отношениях. В то время как оборонительный рефлекс, например на электрический ток, приложенный к коже, делает животное очень возбужденным, постоянно беспокойным, половой - требует особенной обстановки (не говоря уже об его длинной периодичности и зависимости от возраста), пищевой и легкий оборонительный на попадающие в рот негодные в пищу вещества являются ежедневными, нормальными и простыми актами. Другая существеннейшая особенность нашей методики заключается в следующем. Пищевой рефлекс, как и реакция на негодные вещества, попавшие в рот, состоит из двух компонентов. С одной стороны, животное стремится к пище, вводит ее в рот, жует, проглатывает, а негодные вещества выбрасывает изо рта. С другой стороны, к этоймускульной деятельности присоединяется секреторная деятельность. На пищу, как и на негодные вещества, сейчас же льется слюна для механической и химической обработки пищи и для очищения рта от негодных веществ. Мы пользовались в наших опытах специально секреторным компонентом рефлексов. На двигательную же сторону обращалось внимание лишь тогда, когда это являлось почему-либо нужным. Секреторный Сек рефлекс был очень выгоден. При секреции допускается очень точное измерение: мы можем мерить интенсивность слюнного рефлекса или каплями, или делениями цилиндрика или трубки. Это было бы гораздо труднее сделать с двигательным, в этом случае очень сложным и разнообразным, компонентом рефлексов. Для этого потребовались бы тонкие инструменты, да и они, что касается градаций реакции, не достигли бы той точности, как в случае секреторного компонента. Не без значения на первых порах было и то, что при наблюдении за слюноотделением менее навязываются антропоморфические истолкования, чем при наблюдении движений. Все наши опытные собаки подвергаются предварительной легкой операции выведения нормального конца протока слюнной железы на кожу, наружу. Вырезается кусочек слизистой оболочки вокруг того места, где открывается во рту слюнной проток, отпрепаровывается несколько в глубину трубочка протока, и затем конец протока выводится через прорез в стенке рта на кожу и здесь пришивается. В результате слюна течет не в рот, а по щеке или внизу морды. Тогда следить за работой желез очень легко. Стоит приклеить какой-нибудь замазкой (мы употребляем менделеевскую замазку) воронку, и вы можете различным образом точно следить за работой слюнных желез. Или мы наклеиваем герметически на это место стеклянный полушар с двумя трубочками, направленными кверху и книзу. Через нижнюю отсасывается слюна после каждого раздражения. Верхняя трубочка соединяется воздушной передачей со стеклянной трубкой, лежащей горизонтально и наполненной окрашенной жидкостью. И тогда всякое наполнение полушара слюной будет вызывать движение окрашенной жидкости и вы можете отмечать величину этого движения по шкале с делениями. Легко устанавливается и точная автоматическая электрическая регистрация числа капель совершенно одинакового объема. Затем идет общая обстановка опыта. Так как дело касается изучения деятельности больших полушарий, грандиозного сигнального прибора высшей чувствительности, то понятное дело, что через этот прибор постоянно действует на животное множество разнообразных раздражений. Каждое их них оказывает на животное известное действие, а вместе все они сталкиваются и взаимодействуют. Следовательно, если вы не примете никаких мер против этих влияний, часто хаотических, то вы ни в чем не разберетесь, перед вами все перепутается. Необходимо упростить обстановку. Прежде всего обычно мы ставим животное в станок. В прежнее время поступали так, что в отдельной комнате около собаки позволялось находиться только экспериментатору. Но потом оказалось, что и этого недостаточно. Экспериментатор заключает в себе массу раздражений. Каждое его малейшее движение, дыхание, шум дыхания, движения глаз - все это влияет на наше животное и осложняет те явления, которые мы изучаем. Поэтому пришлось вывести экспериментатора за дверь и исключить, хотя и не вполне, его действие на животное. Но и это в обычных лабораториях оказалось недостаточным. Действительно, в них среда около собаки постоянно колеблется: появляются новые звуки, кто-нибудь пройдет, стукнет, заговорит, с улицы доносится шум, дрожит стена от проехавшего экипажа, пробегают в окнах тени, и т. д. В большие полушария, таким образом, вторгаются посторонние случайные раздражения, c которыми приходится считаться. Поэтому при Институте экспериментальной медицины, благодаря средствам одного просвещенного московского коммерсанта, была построена специальная лаборатория. Задача состояла прежде всего в том, чтобы возможно обеспечить эту лабораторию от доступа влияний снаружи. Для этого она была окружена рвом и применено несколько других строительных средств. Затем внутри здания все рабочие комнаты (по четыре в каждом этаже) разделены крестообразным коридором; верхний этаж и нижний, где находятся эти рабочие комнаты, разобщены средним этажом. Наконец в каждой отдельной рабочей комнате особенно старательно, при помощи нескольких плохо проводящих звук материалов, отделено помещение, где находится животное, от части комнаты, где ведет опыт экспериментатор. Для того же, чтобы действовать на животное и отмечать его реакции, устроены вовдушные или электрические провода. Таким образом гарантировано возможное упрощение и постоянство обстановки, в которой находится животное во время экспериментального сеанса. Наконец надо упомянуть еще одно, пока больше pium deciderium. Раз исследуется вся сложность влияний, падающих извне на животное, то, понятно, эта сложность должна быть в руках экспериментатора. Он должен иметь в своем распоряжении очень много инструментов, чтобы можно было действовать то одним, то другим раздражителем и делать из этих раздражителей различные комбинации, как это делает и сама жизнь. И нам часто приходилось и приходится очень чувствовать недостаток в особенности наших, а также вообще современных инструментов для исследования. Работа больших поостоянно идет дальше, чем это позволяли исследовать нам наши инструменты. Может быть, кто-нибудь, слушая об обстановке наших опытов, возразит, что это очень искусственная обстановка. Вот наш ответ на это. Во-первых, едва ли возможно, ввиду бесконечного разнообразия жизненных отношений, применить действительно что-нибудь небывалое, абсолютно новое. Во-вторых, при изучении хаотически сложных явлений неизбежно нарочитое раздробление их, их разгруппирование. Разве физиология животных не пользовалась и не пользуется постоянно до сих пор вивисекцией и даже методом изолированных органов и тканей? Мы ставим наше животное под ограниченное числопределенных условий и таким образом получаем возможность изучать влияние их одно за другим. И вы увидите впоследствии много раз, как, связанные с нашей обстановкой исследования, вариации жизненных состояний животного дали нам в руки очень важные факты. Вот в общем наша принципиальная наша техническая метедика. А затем приступаем к изучению самой сигнальной деятельности больших полушарий и начнем с опыта. О пы т. Вот животное, которое приготовлено так, как я вам описывал. Как видите, пока на него не действует специальный агент, слюнная железа его находится в покое, слюны нет. А вот сейчас мы начнем действовать на ухо собаки ударами метронома. Вы видите, что через 9 секунд начала отделяться слюна и за 45 секунд упало 11 капель слюны. Следовательно, на ваших глазах под влиянием постороннего для еды раздражения (метрономом) получилась деятельность слюнной железы. и эту деятельность надо рассматривать как компонент пищевого рефлекса. Вы видели и другой, двигательный, компонент этого рефлекса. Собака повернулась в сторону, откуда она обычно получает еду, и начала облизываться. Вот это и есть центральное явление, производимое специально большими полушариями, которым мы и будем все время заниматься. Если взять собаку без больших полушарий, то ни на какой такой раздражитель вы слюны никогда не получите. И вы вместе с тем ясно видите, что эта деятельность есть сигнальная: удары метронома сигнализируют пищу, потому что на них животное отвечает той же реакцией, как и на пищу. Если мы покажем собаке пищу, то будет то же. О п ы т. Мы показываем животному еду, и вы видите, что через 5 5 секунд началось слюноотделение и за 15 секунд выделилось 6 капель слюны. Совершенно то же, как и при метрономе. И это также сигнализация, т. е. дело больших полушарий; образовалась она в течение индивидуального существования животного, а не есть прирожденная реакция. Это установлено И. С. Цитовичем в лаборатории покойного проф. В. И. Вартанова. Цитович взял от матки щенков и кормил их только одним молоком в течение довольно продолжительного времени. Когда щенкам было несколько месяцев, он вывел им наружу слюнные протоки, так что мог следить за отделением слюны. Когда он этим щенкам показывал другую еду, кроме молока, именно мясо и хлеб, то никакого слюноотделения не было. Следовательно, сам по себе вид еды не есть возбудитель слюнной реакции, не есть такой агент, который до рождения связан с ней. И только тогда, когда эти щенки несколько раз поели хлеба и мяса, только тогда и один вид хлеба и мяса стал гнать слюну. А сейчас мы увидим то, что называется рефлексом. О пы т. Мы сразу даем собаке есть, и через 1-2 секунды течет слюна. Это уже действие механических и химических свойств пищи на слизистую оболочку рта, это - рефлекс. Вот почему собака без больших полушарий может умереть с голоду среди пищи: она начнет есть только тогда, когда прикоснется ртом к пище. Теперь становится ясным недостаток рефлексов, как они грубы, ограничены, и как высоко значение сигналов. Затем мы должны заняться ответом на серьезнейший вопрос: что же такое эта сигнализация, как понимать с чисто физиологической точки зрения? Мы знаем, что рефлекс есть непременная закономерная реакния организма на внешний агент, которая осуществляется при помощи определенного отдела нервной системы. Совершенно очевидно, что в сигнализации мы имеем все составные части нервного акта, который называется рефлексом. Для рефлекса необходимо внешнее раздражение; здесь, как вы видели в первом нашем опыте, оно налицо - удары метронома. Они приводят в действие слуховой аппарат собаки, и дальше раздражение по слухувому нерву идет в центральную нервную систему, где оно перебрасывается на нервы, идущие к слюнной железе и возбуждающие ее к работе. В опыте с метрономомогло обратить на себя ваше внимание то обстоятельство, что между началом действия метронома и началом слюноотделения протекло значительное число секунд, между тем как в рефлексах этот промежуток измеряется частями секунды. Но этот длинный латентный период сделан нами самими благодаря особенным приемам. Вообще же и при сигнализации эффект наступает так же быстро и отнюдь не позже, чем при обыкновенных рефлексах, о чем речь будет впереди. Рефлекс характеризует закономерность ответа при совершенно определенных условиях. То же самое и при сигнализации. Правда, здесь эффект зависит от большего количества условий. Но это, конечно, никакой существенной разница с рефлексами не делает. Ведь и рефлексы сплошь и рядом отменяются, задерживаются, при строго определенных условиях. Совершенно так же и тут. Если мы изучили предмет хорошо, то и здесь никаких случайностей нет. Опыт и здесь идет точно по нашему расчету. В той специальной лаборатории, о которой я упомянул выше, очень часто достигается такое положение, что вы сидите 1-2 часа, и ни одной капли слюны не упадет вне зависимости от ваших раздражений; в других, обыкновенных лабораториях, конечно, случайные раздражители часто искажают опыт. После сообщенного не остается ни одного основания не считать и не называть рефлексом то, что я до сих пор обозначал термином «сигнализация». Но есть еще одна сторона дела, которая, как может показаться с первого раза, указывает на существенную разницу между старым рефлексом и этим новым явлением, которое я сейчас также назвал рефлексом. Пища своими механическими и химическими свойствами вызывает рефлекс со дня рождения у всякого животного. А новый рефлекс, пример которого вы видели, постепенно образовался в течение индивидуального существования животного. Не составляет ли это существенную разницу? Не отнимает ли это у нас самое основание называть наш новый рефлекс рефлексом? Да, это бесспорно довод к тому, чтобы выделить, отличить эту реакцию, но что касается научного права называть ее рефлексом, то оно нисколько не нарушается этим. Здесь идет дело совсем о другом вопросе: не о механизме, а об образовании рефлекторного механизма. Возьмите для примера телефонное сообщение. Его можно осуществить на два лада. Я мог бы со своей лабораторией из квартиры соединиться специальным проводом и сразу звонить туда, когда мне нужно. Но и теперь, когда я соединяюсь с лабораторией через центральную станцию, это совершенно такое же телефонное сообщение. Разница только в том, что один раз существует готовый проводниковый путь, а в другой - требуется предварительное замыкание; один раз механизм сообщения готов вполне, в другой раз механизм каждый раз несколько дополняется до полной готовности. То же и в нашем случае: один раз рефлекс готов, в другой раз он должен быть предварительно несколько подготовлен. Таким образом перед нами следующий вопрос, вопрос о конструировании нового рефлекторного механизма. Так как образование нового рефлекса наступает непременно и легко при определенных физиологических условиях, что мы увидим сегодня же несколько позже, то при этом нет никакого основания беспокоиться и думать о том, что мы не принимаем во внимание внутреннего состояния собаки. Явление это при полном знании дела вполне в наших руках, оно совершенно закономерно, и нет никакого повода не считать его таким же физиологическим, как и остальные, с которыми физиолог имеет дело. Новые рефлексы мы назвали условными, противопоставляя их врожденным, как безусловным. Это прилагательное начинает входить в общее употребление. С исследовательской точки зрения такое название вполне оправдывается. По сравнению с врожденными рефлексами это действительно очень обусловленные рефлексы: они, во-первых, требуют для своего возникновения известных условий, во-вторых, они в своей работе зависят от очень большого числа условий. Исследователю при их изучении приходится считаться с очень, очень многим. Но, конечно, наши прилагательные могут быть с правом заменены и другими прилагательными. Можно старые рефлексы назвать прирожденными, а новые приобретенными, или же назвать первые видовыми, потому что они характеризуют вид животного, а последние индивидуальными, потому что они будут варьировать у отдельных животных и даже у одного и того же в разное время, при разных условиях. Будет оправдано и название первых проводниковыми, а вторых замыкательными. Что касается признания нервного замыкания в больших полушариях, образования новых связей, то здесь не может возникнуть никакого возражения с теоретической стороны. В технике, как и в нашей обыденной жизни, так часто применяется сейчас принцип замыкания, что было бы странным, если бы в механизме высшей нервной системы, устанавливающей сложнейшие, тончайшие отношения, этот принцип представлялся неожиданным. Вполне натурально, что, кроме проводникового прибора, существует и замыкательный. Физиолог же тем более не должен ничего иметь против этого, что уже много десятков лет назад в физиологии нервной системы приобрело право гражданства немецкое слово «Bahnung», т. е. понятие о проторении пути, об образовании новых связей. Факт условного рефлекса есть повседревнейший и распространеннейший факт. Это есть, очевидно, то, что мы знаем в себе и в животных под разными названиями: дрессировки, дисциплины, воспитания, привычка. Ведь все это есть связи, которые образовались в течение индивидуальной жизни, связи между определенными внешними агентами и определенной ответной деятельностью. Таким образом с фактом условного рефлекса отдается в руки физиолога огромная часть высшей нервной деятельности, а может быть и вся. Теперь перейдем к вопросу: при каких же условиях образуется условный рефлекс, происходит замыкание нового нервного пути? Основное условие - это совпадение во времени всякого внешнего агента с действием безусловного раздражителя. В нашем случае пища есть безусловный раздражитель пищевой реакции. Так вот, если прием пищи у животного совпадает с действием на него агента, который ранее никакого отношения к еде не имел, то этот агент делается возбудителем той же реакции, как и сама пища. В том случае, который прошел перед вашими глазами, так и было. Мы несколько раз начинали действовать на собаку ударами метронома и сейчас же собаку кормили, т. е. производили врожденный пищевой рефлекс. И когда это повторилось несколько раз, то получилось то, что один метроном начал вызывать слюноотделение и соответствующие движения. То же и в случае оборонительного рефлекса при попадании в рот собаки отвергаемых веществ. Если мы вливаем в рот нашего животного слабый раствор кислоты, то получается безусловный кислотный рефлекс: животное проделывает разные движения, порывисто машет головой с открытым ртом, выбрасывает языком кислоту, и т. д., и вместе с этим у него обильно течет слюна. Совершенно такая же реакция получается и от всякого внешнего агента, который несколько раз по времени совпадал с вливанием собаке в рот кислоты. Итак, первое и основное условие для образования данного условного рефлекса - это совпадение во времени действия ранее индифферентного агента с действием безусловного агента, который вызывает определенный безусловный рефлекс. Второе важное условие состоит в следующем. При образовании условного рефлекса индифферентный агент должен несколько предшествовать действию безусловного раздражителя. Если мы сделаем наоборот и сначала начнем действовать безусловным раздражителем, а затем присоединим индифферентный агент, то условного рефлекса не образуется. A. Н. Крестовников в нашей лаборатории очень разнообразил относящиеся сюда опыты, но результат их остался неизменным. Вот некоторые из его результатов. й собаки было сделано 427 комбинирований запаха ванилина с вливанием кислоты, причем дело начиналось с вливания кислоты, а запах присоединялся спустя 5-10 секунд. Ванилин не сделался условным возбудителем кислотной реакции, между тем как в последующих опытах запах уксусного амила, предшествующий вливанию кислоты, был хорошим условным возбудителем уже только после 20 сочетаний. У другой собаки сильный электрический звонок, начинающий действовать 5-10 секунд после начала еды, не сделался условным возбудителем пищевой реакции после 374 сочетаний, между тем как вертящийся перед глазами собаки предмет, предшествующий еде, уже через 5 сочетаний оказывался условным раздражителем, а затем и тот же электрический звонок, примененный перед едой, стал условным раздражителем после одного такого сочетания. Опыты были проделаны на 5 собаках. Результат оставался тем же, присоединялся ли новый агент к безусловному раздражителю через 5-10 секунд или через 1-2 секунды после начала последнего. При образовании условных рефлексов в этих случаях, ради большей гарантии, тщательно следили как за секреторной, так и за двигательной реакцией животных. Итак, первая группа условий - это отношения во времени между безусловным раздражителем и тем агентом, из которого должен образоваться условный раздражитель. Что касается состояния самих больших полушарий, то здесь для возможности образования условных рефлексов требуется, во-первых, деятельное состояние. Если экспериментальное животное в большей или меньшей степени сонливо, то образование условного рефлекса или очень затягивается и затрудняется, или даже делается совершенно невозможным, т. е. образование новых связей, процедура замыкания новых нервных путей есть функция бодрого состояния животного. Во-вторых, большие полушария должны на время образования нового условного рефлекса быть свободными от других деятельностей. Когда мы вырабатываем новый условный рефлекс, необходимо, чтобы на животное не падали другие внешние раздражения, которые вызывают какую-нибудь другую деятельность организма. Иначе это будет сильно мешать, а во многих случаях и совсем не допустит образования нашего условного рефлекса. Например, если, в то время как мы стараемся образовать условный рефлекс, собаке, стоящей в станке, какой-нибудь частью станка причиняется разрушительное действие (давление, ущемление и т. д.), тогда мы можем много раз соединять наш раздражитель с безусловным раздражителем, по крайнее мере с некоторыми из них, и условный рефлекс не образуется. Или вспомните собаку, о которой я упоминал раньше и которая не выносила стеснения свободных движений при нахождении в станке. Поэтому правило почти без исключения: если мы берем для опыта свежее животное, т. е. не подвергавшееся опытам, о которых идет речь, первый условный рефлекс образуется трудно и часто требует большого времени. И это понятие, так как вся обстановка нашего опыта может вызывать у разных животных массу особенных реакций, т. е. обусловливать ту или другую постороннюю деятельность больших полушарий. Надо прибавить, что если мы не всегда дознаем, что это за посторонние рефлексы, которые мешают образованию нашего условного рефлекса, и не можем их устранить, то в этом случае нам помогает свойство самой нервной деятельности. Если обстановка, в которой постоянно находится животное во время опыта, не заключает в себе чего-либо специально разрушительно действующего, то почти все посторонние мешающие рефлексы со временем, постепенно сами собой теряют силу. Сюда же, в эту группу условий, входит, конечно, и здоровье животных, которое гарантирует нормальное состояние самих больших полушарий и исключает влияние внутренних патологических раздражений, посылаемых в большие полушария. Наконец последняя группа условий касается свойств как того агента, из которого должен образоваться условный раздражитель, так и свойств безусловного раздражителя. Условный рефлекс легко образуется из агентов более или менее индифферентных. Собственно говоря, абсолютно индифферентных агентов нет. Если вы имеете нормальное животное, то y него на малейшее изменение обстановки - какой-нибудь, хотя бы и слабый, звук, малейший запах, изменение освещения комнаты, и т. д. - сейчас же произойдет вышеупомянутый исследовательский рефлекс «что такое?» в виде соответствующей двигательной реакции. Но если этот относительно индифферентный агент будет повторяться, то он очень быстро сам собой теряет свое действие на большие полушария, и таким образом устраняется препятствие для образования условного рефлекса. Но если агент принадлежит к группе сильных разражителей вообще или, тем более, специальных, то, конечно, образование нашего условного рефлекса будет очень затруднено или даже, в исключительных случаях, окажется невозможным. Надо иметь в виду и то, что ведь перед нами в большинстве случаев биография собаки не проходила: мало ли какие у нее в жизни были встречи, мало ли какие у нее могли быть раньше образованы условные связи! Но зато, с другой стороны, оказалось, что мы можем применить в качестве нашего агента даже и сильный безусловный раздражитель и все же превратить его в условных. Возьмем разрушительный раздражитель: сильный электрический ток, приложенный к коже, - поранение и прижигание кожи. Это есть, конечно, безусловный раздражитель оборонительного рефлекса: организм ответит на него сильнейшей двигательной реакцией, направленной или на удаление этого раздражителя, или на устранение от него. Однако и на эти раздражители возможно образовать условный рефлекс другого рода. Разрушительный раздражитель был превращен в условный возбудитель пищевого рефлекса. Когда на кожу действовал сильнейший электрический ток, оборонительной реакции не было и следа, а вместо нее налицо была пищевая реакция: животное поворачивалось и тянулось в сторону, откуда подавалась еда, облизывалась, и у него обильно текла слюна. Вот подлинный протокол опыта из работы М. Н. Ерофеевой на данную тему. После каждого электрического раздражения собака подкарсливалась несколько секунд.
То же самое получалось у собаки, когда кожа ее подвергалась ряду уколов до крови или прижиганию. Когда чувствительные души возмущались этим опытом, то мы могли показать, что они возмущались по недоразумению. Конечно, во внутренний мир собаки мы и при этом не хотели проникать и доискиваться, что она чувствует. Но мы имели совершенно точные доказательства того, что и тончайших объективных явлений, сопровождающих состояние животных, когда они подвергаются сильным разрушительным раздражителям, в данном случае не было. У наших собак, у которых рефлексы были переделаны описанным образом, не происходило при таком раздражении сколько-нибудь значительного изменения ни в пульсе, ни в дыхании, как это непременно было бы в сильной степени, если бы разрушительное раздражение предварительно не было связано с пищевой реакцией. Вот к чему ведет перевод нервного возбуждения с одного пути на другой. Но эта переделка рефлексов зависит вт определенного условия - здесь требуется известное соотношение между безусловными рефлексами. Это превращение безусловного раздражителя одного рефлекса в условный другого делается возможным только в том случае, если первый рефлекс физиологически слабее, биологически менее важен, чем второй. Это, как кажется, следует вывести из дальнейших результатов работы Ерофеевой. Мы разрушали у собаки кожу и из этого сделали условный пищевой раздражитель. Но это произошло, можно думать, только потому. что пищевой рефлекс сильнее, чем оборонительный при разрушении кожи. Все мы хорошо знаем из обыденного наблюдения, что когда у собак идет борьба из-за еды, то кожа у соперников часто оказывается пораненной, т. е. пищевой рефлекс берет перевес над оборонительным. Но есть и предел этому. Есть рефлекс посильнее пищевого рефлекса - это рефлекс жизни и смерти, быть или не быть. С этой точки зрения можно было бы понимать смысл нашего следующего факта, именно, что сильный электрический ток, приложенный к коже, лежащей непосредственно, без толстого мышечного слоя, на кости, не удалось сделать условным возбудителем пищевой реакции вместоборонительной, т. е. афферентные нервы, раздражаемые при разрушении кости и сигнализирующие наиболее серьезную опасность для существования организма, с трудом или совсем не могут временно связываться с отделом мозга, от которого возбуждается пищевая реакция. Кстати, из только что приведенных фактов становится очевидной выгода того, что нами для опытов применяется безусловный пищевой рефлекс, так как он находится у вершины иерархической лестницы рефлексов.
Если, с одной стороны, сильные и даже специализированные агенты, как мы только что видели, призвестных условиях делаются условными раздражителями, то, с другой стороны, конечно, есть минимальная сила агента, ниже которой он не может функционировать в качестве условного раздражителя. Так, из температуры ниже 38-39° С, приложенной к коже, условный тепловой раздражитель не образуется (опыты О. С. Соломонова).
Точно так же, если при помощи очень сильного безусловного раздражителя - пищи, как в нашем случае, - можно превратить в условный раздражитель очень неблагоприятный агент, уже входящий в состав другого рефлекса, даже безусловного, то, наоборот, при слабом безусловном раздражителе и из наименее неблагоприятных агентов, т. е. почти совершенно индифферентных, или совсем не удается образование условного раздражителя, или же он образуется с трудом и только незначительный; причем это или постоянно слабые безусловные раздражители, или временно слабые безусловные раздражители, которые при других состояниях организма являются, наоборот, очень сильными, как, например, та же пища. Если мы возьмем голодное животное, то пища вызывает, конечно, сильный безусловный пищевой рефлекс, и условный рефлекс в таком случае образуется скоро и тоже значительный. У постоянно накормленного экспериментального животного будет меньший безусловный рефлекс и условный или совсем не получится, или образуется очень не скоро.
Принимая во внимание все перечисленные условия, а это совсем не представляет трудного дела, мы непременно получаем условный рефлекс. Тогда почему же не считать образование условного рефлекса чисто физиологическим явлением? Мы произвели на нервную систему собаки ряд определенных внешних воздействий, и в результате закономерно образовалась новая нервная связь, произошло определенное нервное замыкание. После этого перед нами типичный рефлекторный акт, как показано выше. А тогда, где же тут место для каких-то внефизиологических отношений? Почему же и условный рефлекс и образование его не физиология, а что-то другое? не вижу основания думать об этих явлениях иначе и позволяю себе догадываться, что при этих вопросах обычно играет вредную роль человеческое предубеждение, вообще несклонность к детерминизации высшей нервной деятельности вследствие чрезвычайной сложности наших субъективных переживаний, деятельности, конечно, в настоящее время в подавляющем числе случаев еще не анализируемой до конечных определенных раздражений.
После каждого электрического раздражения собака подкарсливалась несколько секунд.
То же самое получалось у собаки, когда кожа ее подвергалась ряду уколов до крови или прижиганию. Когда чувствительные души возмущались этим опытом, то мы могли показать, что они возмущались по недоразумению. Конечно, во внутренний мир собаки мы и при этом не хотели проникать и доискиваться, что она чувствует. Но мы имели совершенно точные доказательства того, что и тончайших объективных явлений, сопровождающих состояние животных, когда они подвергаются сильным разрушительным раздражителям, в данном случае не было. У наших собак, у которых рефлексы были переделаны описанным образом, не происходило при таком раздражении сколько-нибудь значительного изменения ни в пульсе, ни в дыхании, как это непременно было бы в сильной степени, если бы разрушительное раздражение предварительно не было связано с пищевой реакцией. Вот к чему ведет перевод нервного возбуждения с одного пути на другой. Но эта переделка рефлексов зависит вт определенного условия - здесь требуется известное соотношение между безусловными рефлексами. Это превращение безусловного раздражителя одного рефлекса в условный другого делается возможным только в том случае, если первый рефлекс физиологически слабее, биологически менее важен, чем второй. Это, как кажется, следует вывести из дальнейших результатов работы Ерофеевой. Мы разрушали у собаки кожу и из этого сделали условный пищевой раздражитель. Но это произошло, можно думать, только потому. что пищевой рефлекс сильнее, чем оборонительный при разрушении кожи. Все мы хорошо знаем из обыденного наблюдения, что когда у собак идет борьба из-за еды, то кожа у соперников часто оказывается пораненной, т. е. пищевой рефлекс берет перевес над оборонительным. Но есть и предел этому. Есть рефлекс посильнее пищевого рефлекса - это рефлекс жизни и смерти, быть или не быть. С этой точки зрения можно было бы понимать смысл нашего следующего факта, именно, что сильный электрический ток, приложенный к коже, лежащей непосредственно, без толстого мышечного слоя, на кости, не удалось сделать условным возбудителем пищевой реакции вместоборонительной, т. е. афферентные нервы, раздражаемые при разрушении кости и сигнализирующие наиболее серьезную опасность для существования организма, с трудом или совсем не могут временно связываться с отделом мозга, от которого возбуждается пищевая реакция. Кстати, из только что приведенных фактов становится очевидной выгода того, что нами для опытов применяется безусловный пищевой рефлекс, так как он находится у вершины иерархической лестницы рефлексов.
Если, с одной стороны, сильные и даже специализированные агенты, как мы только что видели, призвестных условиях делаются условными раздражителями, то, с другой стороны, конечно, есть минимальная сила агента, ниже которой он не может функционировать в качестве условного раздражителя. Так, из температуры ниже 38-39° С, приложенной к коже, условный тепловой раздражитель не образуется (опыты О. С. Соломонова).
Точно так же, если при помощи очень сильного безусловного раздражителя - пищи, как в нашем случае, - можно превратить в условный раздражитель очень неблагоприятный агент, уже входящий в состав другого рефлекса, даже безусловного, то, наоборот, при слабом безусловном раздражителе и из наименее неблагоприятных агентов, т. е. почти совершенно индифферентных, или совсем не удается образование условного раздражителя, или же он образуется с трудом и только незначительный; причем это или постоянно слабые безусловные раздражители, или временно слабые безусловные раздражители, которые при других состояниях организма являются, наоборот, очень сильными, как, например, та же пища. Если мы возьмем голодное животное, то пища вызывает, конечно, сильный безусловный пищевой рефлекс, и условный рефлекс в таком случае образуется скоро и тоже значительный. У постоянно накормленного экспериментального животного будет меньший безусловный рефлекс и условный или совсем не получится, или образуется очень не скоро.
Принимая во внимание все перечисленные условия, а это совсем не представляет трудного дела, мы непременно получаем условный рефлекс. Тогда почему же не считать образование условного рефлекса чисто физиологическим явлением? Мы произвели на нервную систему собаки ряд определенных внешних воздействий, и в результате закономерно образовалась новая нервная связь, произошло определенное нервное замыкание. После этого перед нами типичный рефлекторный акт, как показано выше. А тогда, где же тут место для каких-то внефизиологических отношений? Почему же и условный рефлекс и образование его не физиология, а что-то другое? не вижу основания думать об этих явлениях иначе и позволяю себе догадываться, что при этих вопросах обычно играет вредную роль человеческое предубеждение, вообще несклонность к детерминизации высшей нервной деятельности вследствие чрезвычайной сложности наших субъективных переживаний, деятельности, конечно, в настоящее время в подавляющем числе случаев еще не анализируемой до конечных определенных раздражений.
Лекция третья. Образование условных рефлексов при помощи условных и автоматических раздражителей. Агенты, из которых могут образоваться условные раздражители. Торможение условных рефлексов: 1) внешнее торможение
Мм. гг.! Предшествующую лекцию мы кончили перечислением тех условий, при которых образуется условный рефлекс, причем этот рефлекс образовывался при помощи безусловного, т. е. новый агент связывался с реакцией, вызываемой одновременно с ним безусловным раздражителем. Но последнее обстоятельство, соучастие именно безусловного раздражителя, не является непременным требованием. Новый условный рефлекс может образоваться и при помощи условного же, только хорошо выработанного. Вы видели условное действие метронома. Это был сильный пищевой раздражитель, хорошо выработанный. Несмотря на необычную обстановку опыта (многолюдная аудитория), он действовал очень точно и значительно. И вот оказывается, что при помощи такого сильного условного раздражителя можно образовать другой условный рефлекс. Если теперь мы возьмем какой-нибудь новый более или менее индифферентный агент и применим его в соединении только с метрономом, т. е. не давая при этом еды животному, то он превращается в пищевой раздражитель (опыты Г. П. Зеленого, Д. С.рсикова и Ю. П. Фролова). Такой рефлекс мы называем вторичным условным рефлексом, или рефлексом второго порядка, В подробностях процедуры образования этого рефлекса есть, однако, существенные особенности. Новый агент не только не должен продолжаться после присоединения выработанного условного раздражителя, но необходимо его прекращение за известный срок до начала действия этого раздражителя. Тогда только новый агент делается значительным и постоянным положительным условным раздражителем. При новых агентах средней физиологической силы этот срок составляет не менее 10 секунд. При сильных он значительно растет. Укорачивая этот срок, сливая во времени действие нового агента с действием условноео раздражителя, мы встречаемся уже с явлением совершенно другого рода. Это один из чрезвычайно интересных и тонких пунктов физиологии больших полушарий, анализируемый в настоящее время, однако, уже достаточно хорошо. Полностью этот предмет может быть изложен только позже, в другом месте лекций. Вот относящийся сюда протокол опыта из работы Фролова (опыт 15 XI 1924). У собаки первичные условные пищевые раздражители удары метронома и звонок; появление перед глазами собаки черного квадрата - вторичный условный раздражитель, полученный сочетанием только с ударами метронома при паузе между ними в 15 секунд. Черный квадрат держится перед глазами 10 секунд, звонок и удары метронома продолжается всякий раз 30 секунд. В данном опыте квадрат применяется десятый раз. При помощи же вторичного условного пищевого раздражителя достигнуть образования третичного условияго рефлекса не удается. Вместо этого при применении указанной процедуры теперь и тут выступают те же явления другого рода. Третичный условный рефлекс получается от условных рефлексов, связанных с безусловным оборонительным рефлексом, именно от условных оборонительных рефлексов на значительный электрический ток, приложенный к коже животного. Однако и здесь дело не идет далее третичного условного рефлекса. А затем снова выступают все те же явления другого рода.
Привожу описание опыта с третичным условным рефлексом из работы Фурсикова.
У собаки безусловным раздражителем был электрический ток, приложенный к коже передней лапы и вызывающий оборонительную реакцию. Механическое раздражение кожи задней лапы, ранее индифферентное для собаки, обычное процедурой стало первичным условным возбудителем оборонительного рефлекса. Из звука бульканья только что описанным приемом образован вторичный условный раздражитель. А сочетанием с бульканьем ранее индифферентный тон в 760 колебаний в секунду стал третичным условным раздражителем. Латентный период при этих условных раздражителях (от первичного к третичному) постепенно увеличивался, a оборонительная реакция становилась менее интенсивной. Но все эти рефлексы при соответствующих подкреплениях держались в течение года. Проба сочетания нового агента (вертящийся предмет перед глазами собаки) с третичным раздражителем повела к явлениям другоео рода, о которых речь будет позже. Полученные таким образом условные рефлексы в целом называются у нас цепными.
Итак, мы имеем два случая образования условных рефлексов: при помощи безусловного рефлекса и при помощи хорошо выработанного условного же.
Но есть и еще более особенный, повидимому, случай образования их.
Давно уже, на основании некоторых соображений, нами были поставлены следующие опыты (опыты H. Подкопаева). Собаке под кожу впрыскивалась небольшая доза апоморфина, и 1--2 минуты спустя в экспериментальной комнате пускался тон определенной высоты, продолжавшийся некоторое время. Во время звучания тона начиналась тошнотная реакция собаки на апоморфин: собака становилась несколько беспокойной, начинала облизываться, наступало слюноотделение, и иногда дело доходило до небольших рвотных движений. Когда это было повторено много раз, то начинающееся звучание тона одно стало вызывать в слабой степени ту же реакцию. К сожалению, в силу служебных отношений Подкопаева, опыты эти не могли быть закончены и варьированы. В недавнее время бактериолог ташкентской лаборатории В. А. Крылов, делая серологические исследования, наблюдал следующее. Он хронически впрыскивал собакам под кожу морфий. Известно, что при подкожном впрыскивании морфия действие его начинается с тошнотного акта, с сильного слюнотечения, рвоты, и затем только наступает сон. Крыловым было замечено, что если регулярно изо дня в день впрыскивался морфий, то очень быстро, через 5-6 дней, от одних только приготовлении и обстановки впрыскивания развивается та же картина на животном, как и при самом впрыскивание морфия: появляются сильнейшее слюнотечение, рвотные движения и рвота, а затем и сон. Таким образом теперь рвотный акт вызывался не самим морфием, который через кровь действует на рвотный центр, а всеми теми внешними раздражениями, которые совпадали по времени с введением морфия. И связь здесь имеется очень разнообразная и отдаленная. лучшем случае уже достаточно появиться перед собакой экспериментатору, чтобы начался указанный симптомокомплекс. Если этого оказывалось мало, то следовало присоединить вынимание и открывание ящика со шприцем, затем - постричь место на коже, протереть его алкоголем и, наконец, произвести впрыскиваниндифферентной жидкости. . Чем больше предшествовало впрыскиваний морфия, тем меньше требовалось перечисленных приемов, чтобы воспроизвести картину отравления. Крылов с легкостью демонстрировал этот факт в нашей лаборатории и здесь несколькими формами опыта установил тождество своего факта с нашими условными рефлексами.
Описанный опыт без труда может быть проделан и перед вами. Вот собака, подвергавшаяся несколько раз впрыскиванию морфия. Ее держит на столе незнакомый ей человек. Она остается спокойной. Теперь перед ней появляется экспериментатор, который производил впрыскивания. Собака начинает беспокоиться и облизываться. Когда он приступил к протиранию кожи, у собаки обильно полилась слюна изо рта появились рвотные движения.
Этот опыт делает понятным давно известный факт, что, например, собаки, лишенные паратиреоидных желез, или собаки с экковской фистулой и перевязанной vena portae, раз поев мяса, затем больше к нему не прикасаются. Очевидно, теперь вид и запах мяса уже вызывают у них в известной степени те патологические раздражения, которые наступали у них при отравлении мясом в указанных условиях, и обусловливают этим отрицательную реакцию на мясо.
После всех приведенных опытов возникает вопрос: как, каким процессом происходит новая нервная связь, нервное замыкание? Ответ с чисто фактической стороны не представляется трудным. Безусловный или хорошо выработанный условный раздражитель, конечно, вызывает деятельное состояние определенного отдела головного мозга. Будем называть этот отдел по общепринятой терминологии центром, не связывая, однако, с этим словом представления о точной анатомической обособленности. Очевидно, что к этому центру направляются раздражения, производимые в то же время внешними агентами в клетках коры больших полушарий; путь для этих раздражений именно к этому центру является особенно облегченным, а после нескольких совпадений - проторенным. Таков явный смысл фактов. Исходя из такого понимания дела, мы в свое время поставили вышеописанный опыт с апоморфином, потом так хорошо подтвержденный опытом с морфием. Если к рефлекторно раздраженному центру направляются раздражения из клеток коры больших полушарий, то то же самое должно было произойти и в отношении автоматически, внутренними агентами, составом и свойствами крови, раздражаемого центра, что подтвердилось. В уже переданном вам фактическом материале заключается относящаяся сюда еще важная подробность. Внешние раздражения, даже со дня рождения животного направлявшиеся в известный центр, могут быть отклонены от него в направлении к другому центру и связаться с ним, если этот последний центр физиологически сильнее первого.
Таким образом это слияние, соединение раздражений, возникающих в разных пунктах головного мозга, это нервное замыкание и есть первый нервный механизм, с которым мы встречаемся при изучении деятельности коры больших полушарий. Конечно, остается дальнейшим вопросом: где именно происходит это замыкание - в коре ли больших полушарий исключительно или при участии и нижележащих частей головного мозга? Мыслимы и та и другая возможности. Возможно, что раздражения из клеток коры больших полушарий направляются прямо в центры, находящиеся вне коры. Но вероятно и другое. Имеющиеся в коре клетки афферентных волокон от всех рабочих органов, от всех пунктов организма, и представляющие собой корковые рецепторные центры всего организма, становясь деятельными во время работы органов, под влиянием безусловных, условных и автоматических раздражителей концентрируют на себя раздражения других клеток коры, производимые агентами внешнего мира. А еще вероятнее, что при целом мозге все раздражения, вызывающие в бодром состоянии животного безусловные рефлексы, прежде всего попадают в определенные клетки больших полушарий, и тогда эти клетки являются пунктами, куда направляются различные раздражения, которые и служат для образования условных рефлексов.
Затем мы можем по порядку перейти к вопросу о том: что же может сделаться условным раздражителем? Этот вопрос не такой простой, как это может представляться с первого взгляда. В общей форме, конечно, ответить легко. Условным раздражителем может сделаться всякий агент природы, для которого только имеется рецепторный аппарат у данного организма. Но это общее положение должно быть затем, с одной стороны, расчленено и дополнено, а с другой - ограничено. Самая первая классификация внешних раздражителей - это по составу раздражителей. Отдельным раздражителем может быть чрезвычайно дробный элемент внешнего агента, например очень малая часть одного и того же тона, определенная, хотя бы и ничтожно разнящаяся, интенсивность света и т. д. Таким образом через одно это почти неизмеримо увеличивается количество возможных раздражителей. Конечно, и здесь предел определяется совершенством, тонкостью рецепторных аппаратов. С другой стороны, природа действует на животное и совокупностью нескольких и часто очень многих элементов, комплексными раздражителями. Когда мы отличаем лицо одного человека от лица другого, мы принимаем одновременно в расчет и формы, и краски, и тени, и размеры; или, когда речь идет об ориентировке в местности, и т. д. Количество этих комплексных раздражителей можно было бы назвать безграничным. Ведь сколько может быть соединений из огромного ряда элементарных раздражителей, как это только что указано выше! Но и здесь имеется предел, определяемый конструкцией больших полушарий. Сейчас я хочу дать только приблизительное понятие о возможном количестве отдельных условных раздражителей. Весь же этот предмет огромной важности будет изложен подробно в позднейших лекциях.
Значит, в условный раздражитель может превратиться возникающее перед организмом явление природы. Но условным раздражителем может сделаться и прекращение явления. Пусть в экспериментальной комнате, например, раздаются удары метронома; собака вводится в комнату при этих ударах, и они все время продолжаются. Теперь, если при такой обстановке мы будем прерывать удары метронома и сейчас же после перерыва будем производить безусловный рефлекс, в нашем случае - подкармливание или вливание кислоты, то после нескольких таких совпадений перерыв ударов окажется условным возбудителем этих безусловных реакций (опыты Г. П. Зеленого и И. С. Маковского).
Условным раздражителем может быть не только прекращение явления, но и его ослабление, происходящее, однако, с известной быстротой. Если условный рефлекс был выработан на прекращение какого-нибудь явления, а затем это явление ослабляется медленно и постепенно, то оно не будет сопровождаться узловным действием. Вот один из опытов Зеленого. Тон ре2 духового камертона известной интенсивности, прекращенный сразу, произвел условное действие в размере 32 капель слюны за минуту. Тот же тон, постепенно в течение 12 минут ослабляемый до полного прекращения, не имел ни малейшего действия.
Следовательно, возбудителем теперь будет ие появление действующего агента, а его исчезание, прекращение или более или менее быстрое ослабление. И таких условных раздражителей можно, конечно, получить множество.
После этогопрежнее положение об агентах, из которых могут образоваться условные раздражители, мы должны видоизменить так, что колебание внешней среды как в положительную, так и в отрицательную сторону может сделаться условным раздражителем.
Следующая группа условных раздражителей, представляющая некоторую вариацию предшествующей, такая. У нас условным раздражителем является не наличный раздражитель, как он сейчас действует на животное, а остаток его действия в нервной системе по его прекращении. Опыт производится таким образом. Мы берем какой-нибудь внешний агент, положим, какой-либо звук, производим его 0.5-1 минуту, затем прекращаем и спустя 1-3 минуты присоединяем подкармливание или вливание кислоты. Если мы повторим такую комбинацию несколько раз, то произойдет следующее. Наш агент сам по себе не будет вызывать реакцию, но, после того как он прекратится, получится соответственно пищевая или кислотная реакция. Условным раздражителем станет не наличный раздражитель, не наш примененный звук, а след его в центральной нервной системе. На этом основании мы различаем рефлексы наличные и следовые.
Представляю протокол опыта со следовым рефлексом из работы Ф. С. Гроссмана.
Механическое раздражение кожи - условный следовой кислотный раздражитель. Это раздражение всякий раз продолжалось минуту, затем следовала пауза тоже в минуту, и потом вливалась в рот кислота.
При помощи же вторичного условного пищевого раздражителя достигнуть образования третичного условияго рефлекса не удается. Вместо этого при применении указанной процедуры теперь и тут выступают те же явления другого рода. Третичный условный рефлекс получается от условных рефлексов, связанных с безусловным оборонительным рефлексом, именно от условных оборонительных рефлексов на значительный электрический ток, приложенный к коже животного. Однако и здесь дело не идет далее третичного условного рефлекса. А затем снова выступают все те же явления другого рода.
Привожу описание опыта с третичным условным рефлексом из работы Фурсикова.
У собаки безусловным раздражителем был электрический ток, приложенный к коже передней лапы и вызывающий оборонительную реакцию. Механическое раздражение кожи задней лапы, ранее индифферентное для собаки, обычное процедурой стало первичным условным возбудителем оборонительного рефлекса. Из звука бульканья только что описанным приемом образован вторичный условный раздражитель. А сочетанием с бульканьем ранее индифферентный тон в 760 колебаний в секунду стал третичным условным раздражителем. Латентный период при этих условных раздражителях (от первичного к третичному) постепенно увеличивался, a оборонительная реакция становилась менее интенсивной. Но все эти рефлексы при соответствующих подкреплениях держались в течение года. Проба сочетания нового агента (вертящийся предмет перед глазами собаки) с третичным раздражителем повела к явлениям другоео рода, о которых речь будет позже. Полученные таким образом условные рефлексы в целом называются у нас цепными.
Итак, мы имеем два случая образования условных рефлексов: при помощи безусловного рефлекса и при помощи хорошо выработанного условного же.
Но есть и еще более особенный, повидимому, случай образования их.
Давно уже, на основании некоторых соображений, нами были поставлены следующие опыты (опыты H. Подкопаева). Собаке под кожу впрыскивалась небольшая доза апоморфина, и 1--2 минуты спустя в экспериментальной комнате пускался тон определенной высоты, продолжавшийся некоторое время. Во время звучания тона начиналась тошнотная реакция собаки на апоморфин: собака становилась несколько беспокойной, начинала облизываться, наступало слюноотделение, и иногда дело доходило до небольших рвотных движений. Когда это было повторено много раз, то начинающееся звучание тона одно стало вызывать в слабой степени ту же реакцию. К сожалению, в силу служебных отношений Подкопаева, опыты эти не могли быть закончены и варьированы. В недавнее время бактериолог ташкентской лаборатории В. А. Крылов, делая серологические исследования, наблюдал следующее. Он хронически впрыскивал собакам под кожу морфий. Известно, что при подкожном впрыскивании морфия действие его начинается с тошнотного акта, с сильного слюнотечения, рвоты, и затем только наступает сон. Крыловым было замечено, что если регулярно изо дня в день впрыскивался морфий, то очень быстро, через 5-6 дней, от одних только приготовлении и обстановки впрыскивания развивается та же картина на животном, как и при самом впрыскивание морфия: появляются сильнейшее слюнотечение, рвотные движения и рвота, а затем и сон. Таким образом теперь рвотный акт вызывался не самим морфием, который через кровь действует на рвотный центр, а всеми теми внешними раздражениями, которые совпадали по времени с введением морфия. И связь здесь имеется очень разнообразная и отдаленная. лучшем случае уже достаточно появиться перед собакой экспериментатору, чтобы начался указанный симптомокомплекс. Если этого оказывалось мало, то следовало присоединить вынимание и открывание ящика со шприцем, затем - постричь место на коже, протереть его алкоголем и, наконец, произвести впрыскиваниндифферентной жидкости. . Чем больше предшествовало впрыскиваний морфия, тем меньше требовалось перечисленных приемов, чтобы воспроизвести картину отравления. Крылов с легкостью демонстрировал этот факт в нашей лаборатории и здесь несколькими формами опыта установил тождество своего факта с нашими условными рефлексами.
Описанный опыт без труда может быть проделан и перед вами. Вот собака, подвергавшаяся несколько раз впрыскиванию морфия. Ее держит на столе незнакомый ей человек. Она остается спокойной. Теперь перед ней появляется экспериментатор, который производил впрыскивания. Собака начинает беспокоиться и облизываться. Когда он приступил к протиранию кожи, у собаки обильно полилась слюна изо рта появились рвотные движения.
Этот опыт делает понятным давно известный факт, что, например, собаки, лишенные паратиреоидных желез, или собаки с экковской фистулой и перевязанной vena portae, раз поев мяса, затем больше к нему не прикасаются. Очевидно, теперь вид и запах мяса уже вызывают у них в известной степени те патологические раздражения, которые наступали у них при отравлении мясом в указанных условиях, и обусловливают этим отрицательную реакцию на мясо.
После всех приведенных опытов возникает вопрос: как, каким процессом происходит новая нервная связь, нервное замыкание? Ответ с чисто фактической стороны не представляется трудным. Безусловный или хорошо выработанный условный раздражитель, конечно, вызывает деятельное состояние определенного отдела головного мозга. Будем называть этот отдел по общепринятой терминологии центром, не связывая, однако, с этим словом представления о точной анатомической обособленности. Очевидно, что к этому центру направляются раздражения, производимые в то же время внешними агентами в клетках коры больших полушарий; путь для этих раздражений именно к этому центру является особенно облегченным, а после нескольких совпадений - проторенным. Таков явный смысл фактов. Исходя из такого понимания дела, мы в свое время поставили вышеописанный опыт с апоморфином, потом так хорошо подтвержденный опытом с морфием. Если к рефлекторно раздраженному центру направляются раздражения из клеток коры больших полушарий, то то же самое должно было произойти и в отношении автоматически, внутренними агентами, составом и свойствами крови, раздражаемого центра, что подтвердилось. В уже переданном вам фактическом материале заключается относящаяся сюда еще важная подробность. Внешние раздражения, даже со дня рождения животного направлявшиеся в известный центр, могут быть отклонены от него в направлении к другому центру и связаться с ним, если этот последний центр физиологически сильнее первого.
Таким образом это слияние, соединение раздражений, возникающих в разных пунктах головного мозга, это нервное замыкание и есть первый нервный механизм, с которым мы встречаемся при изучении деятельности коры больших полушарий. Конечно, остается дальнейшим вопросом: где именно происходит это замыкание - в коре ли больших полушарий исключительно или при участии и нижележащих частей головного мозга? Мыслимы и та и другая возможности. Возможно, что раздражения из клеток коры больших полушарий направляются прямо в центры, находящиеся вне коры. Но вероятно и другое. Имеющиеся в коре клетки афферентных волокон от всех рабочих органов, от всех пунктов организма, и представляющие собой корковые рецепторные центры всего организма, становясь деятельными во время работы органов, под влиянием безусловных, условных и автоматических раздражителей концентрируют на себя раздражения других клеток коры, производимые агентами внешнего мира. А еще вероятнее, что при целом мозге все раздражения, вызывающие в бодром состоянии животного безусловные рефлексы, прежде всего попадают в определенные клетки больших полушарий, и тогда эти клетки являются пунктами, куда направляются различные раздражения, которые и служат для образования условных рефлексов.
Затем мы можем по порядку перейти к вопросу о том: что же может сделаться условным раздражителем? Этот вопрос не такой простой, как это может представляться с первого взгляда. В общей форме, конечно, ответить легко. Условным раздражителем может сделаться всякий агент природы, для которого только имеется рецепторный аппарат у данного организма. Но это общее положение должно быть затем, с одной стороны, расчленено и дополнено, а с другой - ограничено. Самая первая классификация внешних раздражителей - это по составу раздражителей. Отдельным раздражителем может быть чрезвычайно дробный элемент внешнего агента, например очень малая часть одного и того же тона, определенная, хотя бы и ничтожно разнящаяся, интенсивность света и т. д. Таким образом через одно это почти неизмеримо увеличивается количество возможных раздражителей. Конечно, и здесь предел определяется совершенством, тонкостью рецепторных аппаратов. С другой стороны, природа действует на животное и совокупностью нескольких и часто очень многих элементов, комплексными раздражителями. Когда мы отличаем лицо одного человека от лица другого, мы принимаем одновременно в расчет и формы, и краски, и тени, и размеры; или, когда речь идет об ориентировке в местности, и т. д. Количество этих комплексных раздражителей можно было бы назвать безграничным. Ведь сколько может быть соединений из огромного ряда элементарных раздражителей, как это только что указано выше! Но и здесь имеется предел, определяемый конструкцией больших полушарий. Сейчас я хочу дать только приблизительное понятие о возможном количестве отдельных условных раздражителей. Весь же этот предмет огромной важности будет изложен подробно в позднейших лекциях.
Значит, в условный раздражитель может превратиться возникающее перед организмом явление природы. Но условным раздражителем может сделаться и прекращение явления. Пусть в экспериментальной комнате, например, раздаются удары метронома; собака вводится в комнату при этих ударах, и они все время продолжаются. Теперь, если при такой обстановке мы будем прерывать удары метронома и сейчас же после перерыва будем производить безусловный рефлекс, в нашем случае - подкармливание или вливание кислоты, то после нескольких таких совпадений перерыв ударов окажется условным возбудителем этих безусловных реакций (опыты Г. П. Зеленого и И. С. Маковского).
Условным раздражителем может быть не только прекращение явления, но и его ослабление, происходящее, однако, с известной быстротой. Если условный рефлекс был выработан на прекращение какого-нибудь явления, а затем это явление ослабляется медленно и постепенно, то оно не будет сопровождаться узловным действием. Вот один из опытов Зеленого. Тон ре2 духового камертона известной интенсивности, прекращенный сразу, произвел условное действие в размере 32 капель слюны за минуту. Тот же тон, постепенно в течение 12 минут ослабляемый до полного прекращения, не имел ни малейшего действия.
Следовательно, возбудителем теперь будет ие появление действующего агента, а его исчезание, прекращение или более или менее быстрое ослабление. И таких условных раздражителей можно, конечно, получить множество.
После этогопрежнее положение об агентах, из которых могут образоваться условные раздражители, мы должны видоизменить так, что колебание внешней среды как в положительную, так и в отрицательную сторону может сделаться условным раздражителем.
Следующая группа условных раздражителей, представляющая некоторую вариацию предшествующей, такая. У нас условным раздражителем является не наличный раздражитель, как он сейчас действует на животное, а остаток его действия в нервной системе по его прекращении. Опыт производится таким образом. Мы берем какой-нибудь внешний агент, положим, какой-либо звук, производим его 0.5-1 минуту, затем прекращаем и спустя 1-3 минуты присоединяем подкармливание или вливание кислоты. Если мы повторим такую комбинацию несколько раз, то произойдет следующее. Наш агент сам по себе не будет вызывать реакцию, но, после того как он прекратится, получится соответственно пищевая или кислотная реакция. Условным раздражителем станет не наличный раздражитель, не наш примененный звук, а след его в центральной нервной системе. На этом основании мы различаем рефлексы наличные и следовые.
Представляю протокол опыта со следовым рефлексом из работы Ф. С. Гроссмана.
Механическое раздражение кожи - условный следовой кислотный раздражитель. Это раздражение всякий раз продолжалось минуту, затем следовала пауза тоже в минуту, и потом вливалась в рот кислота.
 Что касается следовых рефлексов, то здесь мы различаем случаи, когда рефлекс образован на свежем следе, спустя одну или несколько секунд по прекращении агента, - это будет короткий следовой рефлекс. Если же мы ждем после прекращения значительное время, минуту и больше, прежде чем присоединить безусловный раздражитель, тогда мы говорим, что это длинный, поздний следовой рефлекс.
Мы должны различать эти случаи потому, что между наличными условными рефлексами и поздними следовыми есть довольно существенная разница в свойствах, как это будет показано в своем месте.
Наконец я перехожу к последнему и особому агенту, который, в отличие от всех предшествующих, постоянно и как бы сам собой может делаться условным раздражителем. Что касается следовых раздражений, то здесь места недоумению нет. От всякого раздражения в нервной системе остается некоторое время след. во всех отделах нервной системы мы встречаемся с явлением так называемого последействия. Но наш новый агент, хотя он отнюдь не менее реален, чем все предшествующие, представляет, однако, некоторые затруднения для ясного понимания его натуры. Я начну с описания общего вида опыта. Мы берем собаку и постоянно через определенный промежуток времени ее подкармливаем, а другой собаке, положим, вливаем через такой же промежуток в рот кислоту. Если мы это повторим несколько раз, то получим то, что ровно через этот промежуток без всякой еды и кислоты у первой обнаруживается сама собой пищевая реакция, а у второй - кислотная.
Привожу пример из работы Ю. П. Феокритовой.
Собака на станке подкармливается каждые полчаса. В разных опытах после 1-3 подкармливаний одно подкармливание пропускается. Тогда около тридцатой минуты после предшествующего подкармливания начинается слюноотделение и вместе двигательная пищевая реакция. Это иногда совершенно точно наступает в тридцатую минуту, иногда запаздывает на 1-2 минуты. В промежутках, если опыт проделан достаточное число раз, ни малейшей реакции.
Как же толковать результат опыта? Приходится сказать, что в данном случае время является условным раздражителем.
Только что приведенный опыт можно сделать и в несколько другой форме. Мы можем подкармливать животное через каждые полчаса и вместе присоединять действие какого-нибудь агента, т. e. мы каждые полчаса предваряем подкармливание за несколько секунд каким-нибудь агентом. Тогда образуется суммарный условный раздражитель. Он будет состоять из этого агента и из момента времени, т. e. тридцатой минуты. Если мы попробуем наш агент через 5-8 минут, то он не окажет никакого действия. Если попробуем его позже, действие уже будет, но небольшое. Через 20 минут оно будет больше, через 25 минут еще больше, а через 30 минут будет полный эффект. Если этот агент систематически не подкрепляется в другие моменты, кроме тридцатой минуты, то он перестает действовать даже в двадцать девятую минуту, а только в тридцатую обнаруживает свой полный эффект.
О пыт. Вот опыт тоже из работы Феокритовой.
За 30 секунд до подкармливания применяются удары метронома. Промежуток между подкармливаниями 30 минут.
Что касается следовых рефлексов, то здесь мы различаем случаи, когда рефлекс образован на свежем следе, спустя одну или несколько секунд по прекращении агента, - это будет короткий следовой рефлекс. Если же мы ждем после прекращения значительное время, минуту и больше, прежде чем присоединить безусловный раздражитель, тогда мы говорим, что это длинный, поздний следовой рефлекс.
Мы должны различать эти случаи потому, что между наличными условными рефлексами и поздними следовыми есть довольно существенная разница в свойствах, как это будет показано в своем месте.
Наконец я перехожу к последнему и особому агенту, который, в отличие от всех предшествующих, постоянно и как бы сам собой может делаться условным раздражителем. Что касается следовых раздражений, то здесь места недоумению нет. От всякого раздражения в нервной системе остается некоторое время след. во всех отделах нервной системы мы встречаемся с явлением так называемого последействия. Но наш новый агент, хотя он отнюдь не менее реален, чем все предшествующие, представляет, однако, некоторые затруднения для ясного понимания его натуры. Я начну с описания общего вида опыта. Мы берем собаку и постоянно через определенный промежуток времени ее подкармливаем, а другой собаке, положим, вливаем через такой же промежуток в рот кислоту. Если мы это повторим несколько раз, то получим то, что ровно через этот промежуток без всякой еды и кислоты у первой обнаруживается сама собой пищевая реакция, а у второй - кислотная.
Привожу пример из работы Ю. П. Феокритовой.
Собака на станке подкармливается каждые полчаса. В разных опытах после 1-3 подкармливаний одно подкармливание пропускается. Тогда около тридцатой минуты после предшествующего подкармливания начинается слюноотделение и вместе двигательная пищевая реакция. Это иногда совершенно точно наступает в тридцатую минуту, иногда запаздывает на 1-2 минуты. В промежутках, если опыт проделан достаточное число раз, ни малейшей реакции.
Как же толковать результат опыта? Приходится сказать, что в данном случае время является условным раздражителем.
Только что приведенный опыт можно сделать и в несколько другой форме. Мы можем подкармливать животное через каждые полчаса и вместе присоединять действие какого-нибудь агента, т. e. мы каждые полчаса предваряем подкармливание за несколько секунд каким-нибудь агентом. Тогда образуется суммарный условный раздражитель. Он будет состоять из этого агента и из момента времени, т. e. тридцатой минуты. Если мы попробуем наш агент через 5-8 минут, то он не окажет никакого действия. Если попробуем его позже, действие уже будет, но небольшое. Через 20 минут оно будет больше, через 25 минут еще больше, а через 30 минут будет полный эффект. Если этот агент систематически не подкрепляется в другие моменты, кроме тридцатой минуты, то он перестает действовать даже в двадцать девятую минуту, а только в тридцатую обнаруживает свой полный эффект.
О пыт. Вот опыт тоже из работы Феокритовой.
За 30 секунд до подкармливания применяются удары метронома. Промежуток между подкармливаниями 30 минут.
 Эти промежутки для образования рефлекса на время, конечно, можно брать любые. Мы не брали больше 30 минут.
Как понимать физиологически время в качестве условного раздражителя? На это, конечно, точного, определенного ответа пока дать нельзя. Но к известному пониманию этого подойти можно. Как мы вообще отмечаем время? Мы делаем это при помощи разных циклических явлений, захода и восхода солнца, движения стрелок по циферблату часов, и т. д. Но ведь у нас в теле этих циклических явлений тоже немало. Головной мозг за день получает раздражения, утомляется, затем восстанавливается. Пищеварительный канал периодически то занят пищей, то освобождается от нее, и т. д. И так как каждое состояние органа может отражаться на больших полушариях, то вот и основание, чтобы отличать один момент времени от другого. Возьмем короткие промежутки времени. Когда раздражение только что нанесено, оно чувствуется очень резко. Когда мы входим в комнату с каким-либо запахом, то мы сначала ощущаем его очень сильно, а затем все меньше и меньше. Состояние нервной клетки под влиянием раздражения испытывает ряд изменений. Точно так же и в обратном случае. Когда раздражитель прекращается, то сначала он чувствуется еще очень резко, а затем все бледнее и бледнее, и, наконец, мы совсем его не замечаем. Значит, опять имеется ряд различных состояний нервной клетки. С этой точки зрения можно понять как случаи рефлексов на перерыв раздражителя и следовых рефлексов, так и случаи рефлекса на время. В приведенном опыте животное периодически подкармливалось, ряд органов в связи с этим проделывал определенную деятельность, т. е. переживал ряд определенных последовательных изменений. Все это давало себя знать в больших полушариях, рецептировалось ими, и условным раздражителем делался определенный момент этих изменений.
В заключение положение о возможных условиях раздражителях может быть еще раз видоизменено и расширено следующим образом: бесчисленные колебания как внешней, так и внутренней среды организма, отражаясь каждое в определенных состояниях нервных клеток коры больших полушарий, могут сделаться отдельнловными раздражителями.
Затем я перехожу к следующему обширному отделу фактов До сих пор речь шла о рефлексах положительного характера, т. е. о таких рефлексах, которые дают в окончательном результате положительное действие: движения и деятельность желез, а в отношении самой нервной системы представляет собой процесс возбуждения. Но мы знаем другую половину нервной деятельности, нисколько не уступающую первой по физиологической, жизненной важности, - тормозной процесс. Следовательно, нужно ожидать, что при изучении работы больших полушарий как в высшей степени сложной мы должны будем встретиться и с явлениями торможения, постоянно и чрезвычайно сложно переплетающимися с явлениями раздражения. Но прежде чем заняться ими, считаю надобным остановиться коротенько на центральном задерживании при безусловных рефлексах.
Данные физиологии позволяют при нормальной деятельности отличать два рода центрального торможения, которые можно было бы назвать прямое и косвенное, или внутреннее и внешнее. С одной стороны, мы в разных иннервационных приборах: скелетного движения, кровообращения, дыхания и др., знаем прямое тормозное действие известных афферентных нервов или известных агентов крови на определенные центро. С другой стороны, центральная нервная деятельность переполнена случаями косвенного торможения. Это случаи торможения данного центра в силу того, что одновременно с его наличной работой, раздражениями других афферентных нервов или посредством других автоматических раздражений приводится в деятельное состояние другой центр. То же и в сложнылексах, называемых инстинктами. Например многие насекомые специально в стадии гусениц при малейшем прикосновении к ним моментально обездвиживаются и падают. Очевидно прямое торможение всего нервного двигательного аппарата. С другой стороны, возьмем случай цыпленка, только что вылупившегося из яйца. Он сейчас же обнаруживает пищевой хватательный рефлекс при зрительном раздражении маленьким предметом, лежащим на полу. Но если этот предмет сильно раздражает полость рта, то хватательный рефлекс тормозится и заменяется оборонительным выбрасывательным рефлексом.
Итак, одно торможение есть непосредственный эффект раздражений, приходящих в данный центр, - внутреннее торможение, а другой - результат взаимодействия различных деятельных центров - внешнее торможение.
При условных рефлексах мы также встречаемся с этими двумя родами центрального торможения. Так как внешнее торможение условных рефлексов вполне, без малейшего отличия. отвечает внешнему торможению безусловных рефлексов, то я и начну с него.
Самый простой и обыденный случай такой. Вы находитесь с животным в отдельной экспериментальной комнате, где вся обстановка некоторое время остается без колебаний, а затем вдруг происходит какое-нибудь изменение ее: проник посторонний звук, изменилось быстро освещение (зашло за облака или выступило из-за облаков солнце), проникла из-за двери струя воздуха, да еще с каким-нибудь новых запахом. Все это непременно ведет к большему или меньшему ослаблению или (смотря по силе колебаний) и к совершенному уничтожению условного рефлекса, если он как раз в это время должен был начаться. Объяснение здесь очень простое, трудностей не представляющее. Всякий такой новый раздражитель сейчас же ведет к появлению исследовательского рефлекса, т. e. соответствующий рецептор устанавливается в направлении к новому раздражителю, собака прислушивается, всматривается, принюхивается, и этот исследовательский рефлекс тормозит наш условный рефлекс. Вот почему при изучении условных рефлексов важно иметь то специальное здание, о котором я говорил в предшествующей лекции. Доступ случайных раздражителей туда или исключен, или по крайней мере чрезвычайно ограничен.
Понятное дело, что здесь надо учитывать и следующее обстоятельство. Мы знаем, что всякий агент, хотя бы и быстро исчезающий, действует не только тогда, когда он существует, но и некоторое время по прекращении его, - в нервной системе остается так называемое последействие. Поэтому если мы нашим условным раздражителем будем действовать вскоре после этого агента, то рефлекс будет все-таки более или менее заторможен. Надо прибавить кроме того, что различные посторонние как случайные, так нарочито применяемые раздражители, так сказать, затухают в нервной системе с различной скоростью: один через 2-3 минуты, другой через 10 минут, а есть и такие, которые держатся целые дни. Последнее специально относится ко вкусовым и особенно пищевым раздражителям, и с этим надо серьезно считаться.
Действие постороннего рефлекса будет, конечно, чрезвычайно различным, смотря по тому, с каким условным рефлексом мы имеем дело - с молодым или же с хорошо упроченным, старым. Естественно, что молодой рефлекс легче затормаживается, чем упрощенный. Вот почему в лаборатории раньше, когда экспериментатор сидел в одной комнате с животным, постоянно повторялся забавный факт. Сотрудник, выработав новый условный рефлекс и желая его демонстрировать, приглашал меня и, однако, ничего показать не мог. Стоило мне появиться в комнате, как рефлекс исчезал. Факт простой. Я являлся для собаки новым раздражителем, собака на меня смотрела, обнюхивала, - и этого было достаточно, чтобы молодой, только что образовавшийся рефлекс был заторможен. Или такой случай. Один экспериментатор выработал у собаки хорошие, стойкие рефлексы и много экспериментировал с ними. Но стоило передать эту собаку для работы другому экспериментатору, и у него эти рефлексы некоторое время отсутствовали. То же часто бывает, когда собака переводится из одной рабочей комнаты в другую, особенно из одной лаборатории в другую лабораторию.
Конечно, тем более тормозящим образом действуют по типу внешнего торможения специальные раздражители, например вид птицы для охотничьих собак, вид кошки для многих собак, шорох под полом для некоторых собак и т. п., и вообще очень сильные и чрезвычайные, необычные раздражители. Что касается последних, то здесь дело значительно усложняется. По отношению к сильным и чрезвычайным раздражителям все собаки распадаются на две группы. Одни реагируют на них положительно, агрессивно: рвутся на борьбу с ними, отчаянно лают. Другие обнаруживают пассивно-оборонительную реакцию: то стремятся сорваться со станка и убежать, то делаются как бы о деревнелыми, не делая ни малейшего движения, то дрожат и припадают к станку, то даже мочатся, чего с ними при обычных условиях никогда в станке не случается. Следовательно, у этих собак преобладает тормозной рефлекс, причем торможение простирается и на наш условный рефлекс. Так как здесь торможение первично исходит из других отделов мозга и только отсюда распространяется на пункт условного рефлекса, то позволительно и этот случай торможения отнести к виду внешнего торможения условных рефлексов.
Но все эти случаи, которые я только что перечислил, имеют временный характер, почему мы действующие в этих случаях агенты назвали гаснущими, или временными тормозами. Если они действуют на животное повторно, не сопровождаясь для животного никаким дальнейшим существенным результатом, то рано или поздно они делаются для животного индифферентными и тормозящее их влияние на условные рефлексы прекращается.
Относящийся сюда опыт, т. е. опыт гаснущим тормозом, прошел и перед вашими глазами. Собака, на которой я демонстрировал опыт с метрономом, стояла в нашей аудитории во время лекции накануне, и один из моих сотрудников повторял несколько раз опыт, причем я не привлекал вашего внимания к этому опыту. Дело в том, что опыт вначале и не удавался условный рефлекс был заторможен. Только постепенно животное освободилось от тормозящего влияния новой и особенной обстановки. Несколько лет тому назад я также читал курс об условных рефлексах, и тогда я поступил следующим образом. С самого начала курса я выставил в аудитории нужное число собак, с которыми мои сотрудники проделывали там предварительно и несколько раз соответствующие опыты, независимо от хода моих лекций. А затем я их утилизировал в надлежащий момент изложения - и без малейшей неудачи на протяжении всего курса. К сожалению, сейчас, в силу некоторых внешних обстоятельств, я лишен этой возможности и демонстрировать буду только изредка, главным образом ограничиваясь приведением протоколов опытов из наших напечатанных работ или из производимых сейчас в лаборатории исследований
Но, кроме этой подгруппы, в группе внешнего торможения имеется подгруппа постоянных тормозов, которые с повторением не теряют своего действия, а держатся постоянно. Например, вы имеете кислотный условный рефлекс. Если вы перед этим кислотным рефлексом собаку покормите, т. еозбудите у ней тот отдел мозга, тот центр, который работает, когда животное ест, то после этого на значительный срок кислотный рефлекс будет совершенно или в известной степени заторможен. И так будет всегда, сколько бы вы такой опыт ни повторяли. Значит, это надо считать постоянным тормозом. И таких постоянных тормозов немало. Если мы делаем условный рефлекс из вливания кислоты и по неосторожности (вливая недостаточно разбавленный раствор или вливая слишком часто и много) причиним животному стоматит, то условный рефлекс на кислоту будет задержан до тех пор, пока не будет устранено это патологическое состояние рта. Или, например, может случиться, что собака имеет ранение на коже и в станке лямками раздражается, и тогда присоединяется оборонительный рефлекс, а условные рефлексы, в особенности кислотный, будут заторможены. В этом отношении, конечно, может быть много и других случаев, например следующий. Сначала опыт идет хорошо, а затем вдруг рефлексы падают и, наконец, исчезают. Что такое? Но достаточно свести собаку на двор, где она помочится, она снова дает нормальные рефлексы. Очевидно, раздражение центра мочеиспускательного рефлекса затормозило наши условные рефлексы. Возьмем пору, когда самки находятся в периоде течки. Если самцы перед опытом находятся поблизости самок, то у первых также условные рефлексы оказываются заторможенными в большей или меньшей степени. Здесь тормозит, очевиздражение полового центра, и т. д.
Итак, вы видите, что условий, тормозящих наши рефлексы, очень много, и недаром эти рефлексы названы нами условными. Но, однако, все эти условия легко иметь в руках, их устранять, их не допускать. Это будут все случаи внешнего торможения, все они характеризуются, повторяю еще раз, тем, что, как только в центральной нервной системе возникнет другая, посторонняя нервная деятельность, она сейчас же дает себя знать на уменьшении или исчезании условных рефлексов, но временно, пока существует обусловивший ее раздражитель или его последействие.
Эти промежутки для образования рефлекса на время, конечно, можно брать любые. Мы не брали больше 30 минут.
Как понимать физиологически время в качестве условного раздражителя? На это, конечно, точного, определенного ответа пока дать нельзя. Но к известному пониманию этого подойти можно. Как мы вообще отмечаем время? Мы делаем это при помощи разных циклических явлений, захода и восхода солнца, движения стрелок по циферблату часов, и т. д. Но ведь у нас в теле этих циклических явлений тоже немало. Головной мозг за день получает раздражения, утомляется, затем восстанавливается. Пищеварительный канал периодически то занят пищей, то освобождается от нее, и т. д. И так как каждое состояние органа может отражаться на больших полушариях, то вот и основание, чтобы отличать один момент времени от другого. Возьмем короткие промежутки времени. Когда раздражение только что нанесено, оно чувствуется очень резко. Когда мы входим в комнату с каким-либо запахом, то мы сначала ощущаем его очень сильно, а затем все меньше и меньше. Состояние нервной клетки под влиянием раздражения испытывает ряд изменений. Точно так же и в обратном случае. Когда раздражитель прекращается, то сначала он чувствуется еще очень резко, а затем все бледнее и бледнее, и, наконец, мы совсем его не замечаем. Значит, опять имеется ряд различных состояний нервной клетки. С этой точки зрения можно понять как случаи рефлексов на перерыв раздражителя и следовых рефлексов, так и случаи рефлекса на время. В приведенном опыте животное периодически подкармливалось, ряд органов в связи с этим проделывал определенную деятельность, т. е. переживал ряд определенных последовательных изменений. Все это давало себя знать в больших полушариях, рецептировалось ими, и условным раздражителем делался определенный момент этих изменений.
В заключение положение о возможных условиях раздражителях может быть еще раз видоизменено и расширено следующим образом: бесчисленные колебания как внешней, так и внутренней среды организма, отражаясь каждое в определенных состояниях нервных клеток коры больших полушарий, могут сделаться отдельнловными раздражителями.
Затем я перехожу к следующему обширному отделу фактов До сих пор речь шла о рефлексах положительного характера, т. е. о таких рефлексах, которые дают в окончательном результате положительное действие: движения и деятельность желез, а в отношении самой нервной системы представляет собой процесс возбуждения. Но мы знаем другую половину нервной деятельности, нисколько не уступающую первой по физиологической, жизненной важности, - тормозной процесс. Следовательно, нужно ожидать, что при изучении работы больших полушарий как в высшей степени сложной мы должны будем встретиться и с явлениями торможения, постоянно и чрезвычайно сложно переплетающимися с явлениями раздражения. Но прежде чем заняться ими, считаю надобным остановиться коротенько на центральном задерживании при безусловных рефлексах.
Данные физиологии позволяют при нормальной деятельности отличать два рода центрального торможения, которые можно было бы назвать прямое и косвенное, или внутреннее и внешнее. С одной стороны, мы в разных иннервационных приборах: скелетного движения, кровообращения, дыхания и др., знаем прямое тормозное действие известных афферентных нервов или известных агентов крови на определенные центро. С другой стороны, центральная нервная деятельность переполнена случаями косвенного торможения. Это случаи торможения данного центра в силу того, что одновременно с его наличной работой, раздражениями других афферентных нервов или посредством других автоматических раздражений приводится в деятельное состояние другой центр. То же и в сложнылексах, называемых инстинктами. Например многие насекомые специально в стадии гусениц при малейшем прикосновении к ним моментально обездвиживаются и падают. Очевидно прямое торможение всего нервного двигательного аппарата. С другой стороны, возьмем случай цыпленка, только что вылупившегося из яйца. Он сейчас же обнаруживает пищевой хватательный рефлекс при зрительном раздражении маленьким предметом, лежащим на полу. Но если этот предмет сильно раздражает полость рта, то хватательный рефлекс тормозится и заменяется оборонительным выбрасывательным рефлексом.
Итак, одно торможение есть непосредственный эффект раздражений, приходящих в данный центр, - внутреннее торможение, а другой - результат взаимодействия различных деятельных центров - внешнее торможение.
При условных рефлексах мы также встречаемся с этими двумя родами центрального торможения. Так как внешнее торможение условных рефлексов вполне, без малейшего отличия. отвечает внешнему торможению безусловных рефлексов, то я и начну с него.
Самый простой и обыденный случай такой. Вы находитесь с животным в отдельной экспериментальной комнате, где вся обстановка некоторое время остается без колебаний, а затем вдруг происходит какое-нибудь изменение ее: проник посторонний звук, изменилось быстро освещение (зашло за облака или выступило из-за облаков солнце), проникла из-за двери струя воздуха, да еще с каким-нибудь новых запахом. Все это непременно ведет к большему или меньшему ослаблению или (смотря по силе колебаний) и к совершенному уничтожению условного рефлекса, если он как раз в это время должен был начаться. Объяснение здесь очень простое, трудностей не представляющее. Всякий такой новый раздражитель сейчас же ведет к появлению исследовательского рефлекса, т. e. соответствующий рецептор устанавливается в направлении к новому раздражителю, собака прислушивается, всматривается, принюхивается, и этот исследовательский рефлекс тормозит наш условный рефлекс. Вот почему при изучении условных рефлексов важно иметь то специальное здание, о котором я говорил в предшествующей лекции. Доступ случайных раздражителей туда или исключен, или по крайней мере чрезвычайно ограничен.
Понятное дело, что здесь надо учитывать и следующее обстоятельство. Мы знаем, что всякий агент, хотя бы и быстро исчезающий, действует не только тогда, когда он существует, но и некоторое время по прекращении его, - в нервной системе остается так называемое последействие. Поэтому если мы нашим условным раздражителем будем действовать вскоре после этого агента, то рефлекс будет все-таки более или менее заторможен. Надо прибавить кроме того, что различные посторонние как случайные, так нарочито применяемые раздражители, так сказать, затухают в нервной системе с различной скоростью: один через 2-3 минуты, другой через 10 минут, а есть и такие, которые держатся целые дни. Последнее специально относится ко вкусовым и особенно пищевым раздражителям, и с этим надо серьезно считаться.
Действие постороннего рефлекса будет, конечно, чрезвычайно различным, смотря по тому, с каким условным рефлексом мы имеем дело - с молодым или же с хорошо упроченным, старым. Естественно, что молодой рефлекс легче затормаживается, чем упрощенный. Вот почему в лаборатории раньше, когда экспериментатор сидел в одной комнате с животным, постоянно повторялся забавный факт. Сотрудник, выработав новый условный рефлекс и желая его демонстрировать, приглашал меня и, однако, ничего показать не мог. Стоило мне появиться в комнате, как рефлекс исчезал. Факт простой. Я являлся для собаки новым раздражителем, собака на меня смотрела, обнюхивала, - и этого было достаточно, чтобы молодой, только что образовавшийся рефлекс был заторможен. Или такой случай. Один экспериментатор выработал у собаки хорошие, стойкие рефлексы и много экспериментировал с ними. Но стоило передать эту собаку для работы другому экспериментатору, и у него эти рефлексы некоторое время отсутствовали. То же часто бывает, когда собака переводится из одной рабочей комнаты в другую, особенно из одной лаборатории в другую лабораторию.
Конечно, тем более тормозящим образом действуют по типу внешнего торможения специальные раздражители, например вид птицы для охотничьих собак, вид кошки для многих собак, шорох под полом для некоторых собак и т. п., и вообще очень сильные и чрезвычайные, необычные раздражители. Что касается последних, то здесь дело значительно усложняется. По отношению к сильным и чрезвычайным раздражителям все собаки распадаются на две группы. Одни реагируют на них положительно, агрессивно: рвутся на борьбу с ними, отчаянно лают. Другие обнаруживают пассивно-оборонительную реакцию: то стремятся сорваться со станка и убежать, то делаются как бы о деревнелыми, не делая ни малейшего движения, то дрожат и припадают к станку, то даже мочатся, чего с ними при обычных условиях никогда в станке не случается. Следовательно, у этих собак преобладает тормозной рефлекс, причем торможение простирается и на наш условный рефлекс. Так как здесь торможение первично исходит из других отделов мозга и только отсюда распространяется на пункт условного рефлекса, то позволительно и этот случай торможения отнести к виду внешнего торможения условных рефлексов.
Но все эти случаи, которые я только что перечислил, имеют временный характер, почему мы действующие в этих случаях агенты назвали гаснущими, или временными тормозами. Если они действуют на животное повторно, не сопровождаясь для животного никаким дальнейшим существенным результатом, то рано или поздно они делаются для животного индифферентными и тормозящее их влияние на условные рефлексы прекращается.
Относящийся сюда опыт, т. е. опыт гаснущим тормозом, прошел и перед вашими глазами. Собака, на которой я демонстрировал опыт с метрономом, стояла в нашей аудитории во время лекции накануне, и один из моих сотрудников повторял несколько раз опыт, причем я не привлекал вашего внимания к этому опыту. Дело в том, что опыт вначале и не удавался условный рефлекс был заторможен. Только постепенно животное освободилось от тормозящего влияния новой и особенной обстановки. Несколько лет тому назад я также читал курс об условных рефлексах, и тогда я поступил следующим образом. С самого начала курса я выставил в аудитории нужное число собак, с которыми мои сотрудники проделывали там предварительно и несколько раз соответствующие опыты, независимо от хода моих лекций. А затем я их утилизировал в надлежащий момент изложения - и без малейшей неудачи на протяжении всего курса. К сожалению, сейчас, в силу некоторых внешних обстоятельств, я лишен этой возможности и демонстрировать буду только изредка, главным образом ограничиваясь приведением протоколов опытов из наших напечатанных работ или из производимых сейчас в лаборатории исследований
Но, кроме этой подгруппы, в группе внешнего торможения имеется подгруппа постоянных тормозов, которые с повторением не теряют своего действия, а держатся постоянно. Например, вы имеете кислотный условный рефлекс. Если вы перед этим кислотным рефлексом собаку покормите, т. еозбудите у ней тот отдел мозга, тот центр, который работает, когда животное ест, то после этого на значительный срок кислотный рефлекс будет совершенно или в известной степени заторможен. И так будет всегда, сколько бы вы такой опыт ни повторяли. Значит, это надо считать постоянным тормозом. И таких постоянных тормозов немало. Если мы делаем условный рефлекс из вливания кислоты и по неосторожности (вливая недостаточно разбавленный раствор или вливая слишком часто и много) причиним животному стоматит, то условный рефлекс на кислоту будет задержан до тех пор, пока не будет устранено это патологическое состояние рта. Или, например, может случиться, что собака имеет ранение на коже и в станке лямками раздражается, и тогда присоединяется оборонительный рефлекс, а условные рефлексы, в особенности кислотный, будут заторможены. В этом отношении, конечно, может быть много и других случаев, например следующий. Сначала опыт идет хорошо, а затем вдруг рефлексы падают и, наконец, исчезают. Что такое? Но достаточно свести собаку на двор, где она помочится, она снова дает нормальные рефлексы. Очевидно, раздражение центра мочеиспускательного рефлекса затормозило наши условные рефлексы. Возьмем пору, когда самки находятся в периоде течки. Если самцы перед опытом находятся поблизости самок, то у первых также условные рефлексы оказываются заторможенными в большей или меньшей степени. Здесь тормозит, очевиздражение полового центра, и т. д.
Итак, вы видите, что условий, тормозящих наши рефлексы, очень много, и недаром эти рефлексы названы нами условными. Но, однако, все эти условия легко иметь в руках, их устранять, их не допускать. Это будут все случаи внешнего торможения, все они характеризуются, повторяю еще раз, тем, что, как только в центральной нервной системе возникнет другая, посторонняя нервная деятельность, она сейчас же дает себя знать на уменьшении или исчезании условных рефлексов, но временно, пока существует обусловивший ее раздражитель или его последействие.
Лекция четвертая. 2) внутреннее торможение условных рефлексов: a) угасание условных рефлексов
Мм. гг.! В конце предшествующей лекции познакомил вас с так называемым нами внешним торможением условных рефлексов, т. е. многочисленными случаями временного столкновения их с другими посторонними раздражениями мозга, причем наши рефлексы на это время слабнут в большей или меньшей степени или даже совершенно исчезают. Теперь мы приступаем к другому роду торможения, называемого нами внутренним, когда положительный условный раздражитель при определенных условиях сам превращается в тормозной, отрицательный, т. е. когда он в клетках больших полушарий производит не процесс возбуждения, а процесс торможения. Таким образом мы получаем рядом с положительными условными рефлексами отрицательные условные рефлексы. Явное, бросающееся в глаза различие между внешним и внутренним торможением состоит в том, что внешнее торможение возникает сейчас же при указанных выше условиях, внутреннее же торможение непременно развивается постепенно, а иногда вырабатывается очень медленно и даже с трудов. Я начну с того случая внутреннего торможения, с которым мы при изучении условных рефлексов встретились прежде всего, и опишу его, что касается понимания его нами, так сказать, до известной степени исторически, так как понимание это, конечно, складывалось постепенно. Перед вами та же собака, и действовать на нее мы будем тем же условным раздражителем - ударами метронома. Мой помощник будет громко считать число капель слюны за 30 секунд действия метронома и отмечать промежуток в секундах между началом раздражения и началом слюноотделения. Этот промежуток мы привыкли называть латентным периодом, хотя в нашем случае было бы рациональнее изобрести для обозначения его другой термин, как увидим это дальше. На этот раз за раздражением метрономом не последует, как обыкновенно, подкармливание животного, мы не подкрепим условного рефлекса (наше обычное выражение), а через 2 минуты повторим то же самое, и так несколько раз. Мы получим следующее: На этом мы опыт временно остановим, чтобы в сегодняшней же лекции иметь возможность прибавить к нему важную подробность. Ясно, что с повторением при данном условии условный рефлекс слабеет. Если бы мы опыт продолжали дальше, дело кончилось бы полным отсутствием слюноотделения.
Это Это явление быстрой и более или менее постепенной потери условным раздражителем своего действия, раз он не сопровождается соответствующим безусловным, мы, не предрешая вопроса об его натуре, назвали угасанием и собрали относительно него очень большой материал, к передаче которого я теперь и перейду.
Только - предварительно несколько слов еще о нашей терминологии. Мы ранее часто различали прилагательными - натуральный и искусственный - условный рефлекс, который образовался или образовывался, так сказать, сам собой на пищу на расстоянии и на подробности процедуры вливания кислоты, от рефлекса на агенты, не имеющие обыкновенно никакого отношения ни к еде, ни к введению в рот отвергаемых веществ. В свойствах этих рефлексов нет ни малейшей разницы. Бесчисленные посторонние агенты, из которых мы делали и постоянно делаем условные раздражители, были важны для нас не только потому, что их легко было сделать вполне однообразными, точными и удобно регулируемыми в отношении интенсивности, но и потому, что они открывали для нас огромный горизонт исследования, как увидим это дальше. Таким образом то, что сначала было применено нами только для проверки правильности нашего понимания механизма образования натуральных рефлексов, впоследствии сделалось главным материалом нашего исследования. Я упоминаю об этом сейчас потому, что на первых порах нашей работы было сделано много опытов именно с натуральными условными рефлексами, и сейчас я буду приводить немало протоколов таких опытов.
Чт. Что касается хода угасания, его постепенности и правильности, то здесь часто наблюдаются колебания. Эти колебания зависят от двоякого рода условии. Во-первых, от внешних. Для правильного падения условных повторно неподкрепляемых рефлексов требуются как однообразие и постоянство условного раздражителя, так и отсутствие даже малейших изменений в обстановке животного. При натуральных условных рефлексах нахождение пищи перед животным то вблизи, то несколько дальше, то неподвижное, то перемещающееся хотя бы и незначительно, обусловливает очень большие колебания (то вверх, то вниз) угасающего рефлекса. При искусственных агентах, конечно, легко достигается абсолютное постоянство раздражения, и этот повод колебаний исключается вполне. Резкие изменения обстановки, как случай внешнего торможения, понятно, быстро снижают рефлекс, с тем, чтобы по миновании этих изменений он опять несколько поднялся. Но особенно интересно влияние незначительных изменений обстановки. Они временно ослабляют или устраняют угасание. Такой случай произошел в нашем сегодняшнем опыте при пятом повторении рефлекса. Повышение рефлекса с 5 капель до 7 отчетливо совпало с некоторым движением среди слушателей в этот момент. Это очень важный пункт в физиологии больших полушарий, и к нему мы вернемся еще раз несколько позже в сегодняшней же лекции. Но и при постоянстве как раздражителя, так и обстановки все же иногда наблюдаются колебания, и притом ритмического характера. И это, очевидно, от условий второго рода, внутренних условий происходящего при угасании нервного процесса. С этим явлением впоследствии мы будем встречаться много раз.
Быстрота угасания, т. е. как скоро эффект повторяемого без подкрепления условного раздражителя доходит до нуля, также определяется многими условиями.
Прежде всего надо упомянуть об индивидуальности животных. У одних животных условные рефлексы, при прочих равных условиях, угасают быстро, у других очень медленно. Это отчетливо связано с общим характером нервной системы животного. У живых, возбудимых собак рефлексы угасают большей частью медленно, у спокойных, так сказать, солидных, угасают быстро.
Затем понятным образом имеет значение степень выработанности, прочности рефлекса. Чем он моложе, менее укреплен, тем скорее угасает, и наоборот.
Резкое влияние на быстроту угасания оказывает интенсивность безусловного рефлекса, при помощи которого образован условный.
Вот относящиеся сюда опыты из работы Б. П. Бабкина на одной и той же собаке.
Определенное количество 1% 0/ раствора extr. Quassiae в среднем из 10 опытов дает 1.71 куб. см слюны безусловного рефлекса и условный рефлекс в размере 0.3 куб. см за минуту раздражения. Определенное количество 0.1% раствора соляной кислоты в среднем из 5 опытов дает 5.2 куб. см слюны безусловного рефлекса и 0.9 куб. см условного за то же время. В обоих случаях дело идет о натуральном условном рефлексе, т. е. о действии веществ на расстоянии. Опыты ведутся при прочих равных условиях. Цифры представляют ход угасания.
На этом мы опыт временно остановим, чтобы в сегодняшней же лекции иметь возможность прибавить к нему важную подробность. Ясно, что с повторением при данном условии условный рефлекс слабеет. Если бы мы опыт продолжали дальше, дело кончилось бы полным отсутствием слюноотделения.
Это Это явление быстрой и более или менее постепенной потери условным раздражителем своего действия, раз он не сопровождается соответствующим безусловным, мы, не предрешая вопроса об его натуре, назвали угасанием и собрали относительно него очень большой материал, к передаче которого я теперь и перейду.
Только - предварительно несколько слов еще о нашей терминологии. Мы ранее часто различали прилагательными - натуральный и искусственный - условный рефлекс, который образовался или образовывался, так сказать, сам собой на пищу на расстоянии и на подробности процедуры вливания кислоты, от рефлекса на агенты, не имеющие обыкновенно никакого отношения ни к еде, ни к введению в рот отвергаемых веществ. В свойствах этих рефлексов нет ни малейшей разницы. Бесчисленные посторонние агенты, из которых мы делали и постоянно делаем условные раздражители, были важны для нас не только потому, что их легко было сделать вполне однообразными, точными и удобно регулируемыми в отношении интенсивности, но и потому, что они открывали для нас огромный горизонт исследования, как увидим это дальше. Таким образом то, что сначала было применено нами только для проверки правильности нашего понимания механизма образования натуральных рефлексов, впоследствии сделалось главным материалом нашего исследования. Я упоминаю об этом сейчас потому, что на первых порах нашей работы было сделано много опытов именно с натуральными условными рефлексами, и сейчас я буду приводить немало протоколов таких опытов.
Чт. Что касается хода угасания, его постепенности и правильности, то здесь часто наблюдаются колебания. Эти колебания зависят от двоякого рода условии. Во-первых, от внешних. Для правильного падения условных повторно неподкрепляемых рефлексов требуются как однообразие и постоянство условного раздражителя, так и отсутствие даже малейших изменений в обстановке животного. При натуральных условных рефлексах нахождение пищи перед животным то вблизи, то несколько дальше, то неподвижное, то перемещающееся хотя бы и незначительно, обусловливает очень большие колебания (то вверх, то вниз) угасающего рефлекса. При искусственных агентах, конечно, легко достигается абсолютное постоянство раздражения, и этот повод колебаний исключается вполне. Резкие изменения обстановки, как случай внешнего торможения, понятно, быстро снижают рефлекс, с тем, чтобы по миновании этих изменений он опять несколько поднялся. Но особенно интересно влияние незначительных изменений обстановки. Они временно ослабляют или устраняют угасание. Такой случай произошел в нашем сегодняшнем опыте при пятом повторении рефлекса. Повышение рефлекса с 5 капель до 7 отчетливо совпало с некоторым движением среди слушателей в этот момент. Это очень важный пункт в физиологии больших полушарий, и к нему мы вернемся еще раз несколько позже в сегодняшней же лекции. Но и при постоянстве как раздражителя, так и обстановки все же иногда наблюдаются колебания, и притом ритмического характера. И это, очевидно, от условий второго рода, внутренних условий происходящего при угасании нервного процесса. С этим явлением впоследствии мы будем встречаться много раз.
Быстрота угасания, т. е. как скоро эффект повторяемого без подкрепления условного раздражителя доходит до нуля, также определяется многими условиями.
Прежде всего надо упомянуть об индивидуальности животных. У одних животных условные рефлексы, при прочих равных условиях, угасают быстро, у других очень медленно. Это отчетливо связано с общим характером нервной системы животного. У живых, возбудимых собак рефлексы угасают большей частью медленно, у спокойных, так сказать, солидных, угасают быстро.
Затем понятным образом имеет значение степень выработанности, прочности рефлекса. Чем он моложе, менее укреплен, тем скорее угасает, и наоборот.
Резкое влияние на быстроту угасания оказывает интенсивность безусловного рефлекса, при помощи которого образован условный.
Вот относящиеся сюда опыты из работы Б. П. Бабкина на одной и той же собаке.
Определенное количество 1% 0/ раствора extr. Quassiae в среднем из 10 опытов дает 1.71 куб. см слюны безусловного рефлекса и условный рефлекс в размере 0.3 куб. см за минуту раздражения. Определенное количество 0.1% раствора соляной кислоты в среднем из 5 опытов дает 5.2 куб. см слюны безусловного рефлекса и 0.9 куб. см условного за то же время. В обоих случаях дело идет о натуральном условном рефлексе, т. е. о действии веществ на расстоянии. Опыты ведутся при прочих равных условиях. Цифры представляют ход угасания.
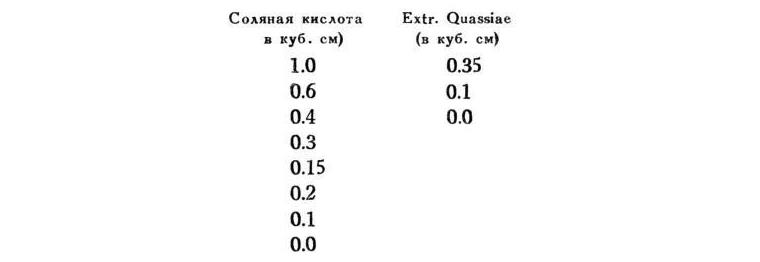 Общий срок угасания отчетливо связан с великиной промежутка между повторениями угашаемого рефлекса. Чем меньше промежуток, тем в меньший период времени достигается окончательное угасание рефлекса, при этом большей частью при меньшем числе повторений, и наоборот.
Представляю опыт из работы Бабкина для иллюстрации этого отношения.
Раздражение животного через равные промежутки времени в продолжение минуты мясным порошком на расстоянии. Следующие пять рядов угасания все произведены в один и тот же день. В промежутках между этими рядами собака получала отдых и подкармливалась мясным порошком.
Общий срок угасания отчетливо связан с великиной промежутка между повторениями угашаемого рефлекса. Чем меньше промежуток, тем в меньший период времени достигается окончательное угасание рефлекса, при этом большей частью при меньшем числе повторений, и наоборот.
Представляю опыт из работы Бабкина для иллюстрации этого отношения.
Раздражение животного через равные промежутки времени в продолжение минуты мясным порошком на расстоянии. Следующие пять рядов угасания все произведены в один и тот же день. В промежутках между этими рядами собака получала отдых и подкармливалась мясным порошком.

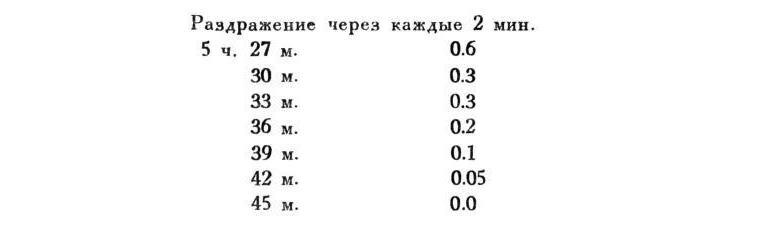 Таким образом рефлекс при промежутке:
Таким образом рефлекс при промежутке:
 Последнее условие - это просто повторение опытов с угасанием на одном и том же животном. По мере повторения этих опытов число раздражений до окончательного угасания, при прочих равных условиях, становится все меньше и меньше, пока, наконец, полное угасание не наступает уже после одного неподкрепления условного рефлекса, по крайней мере у части животных.
Исключительный интерес в факте угасания привязывается к следующему обстоятельству. Падение рефлекса при угасании обнаруживается не только на том условном рефлексе, который непосредственно подвергся процедуре угашения (первично угашейный рефлекс), но и на других, не тронутых, так сказать, прямо этой процедурой (вторично угашенные рефлексы), и не только на других, связанных с одним и тем же безусловным раздражителем (однородных, по нашей терминологии), но и на выработанных при помощи других безусловных раздражителей (разнородных), и, наконец, при высших степенях угасания, даже на самих безусловных. Так, у одной собаки определенное количество соляной кислоты давало в среднем около 6 куб. см слюны. То же количество соляной кислоты после значительного угашения условного кожно-механического кислотного рефлекса дало только 3.8 куб. см (опыт И. Я. Перельцвейга). Наиболее резкий факт - это вторичное угасание в большей или меньшей степени однородных условных рефлексов; он у нас и обследован более подробно.
Для всех случаев вторичного угасания имеет первенствующее значение степень угашения. Очень сильное угашение многое сравнивает, стирает мелкие отличия. Пмеренном же угашении выступают многие подробности. Принимая остальные условия равными, можно сказать, что последующий эффект угашения одного условного рефлекса на другие, однородные с ним, определяется относительной физиологической силой этих последних. А эта сила в свою очередь зависит от возраста рефлекса, от постоянных или редких, или даже прекратившихся подкреплений его, от частоты предшествовавших угощений и, наконец, от предварительного подкрепления в данный экспериментальный сеанс. Чем Чем больше физиологическая сила данного рефлекса сравнительно с силой первично угашаемого или угашенного рефлекса, тем он менее подвергается вторичному угасанию, и, наоборот, при первичном угашении сильнейшего рефлекса вторично вполне гаснут все слабые.
Вот опыт из работ Бабкина.
У собаки три условных раздражителя кислотного рефлекса: звонок, удары метронома и механическое раздражение кожи.
Условное раздражение продолжается 30 секунд.
Опыт, произведенный накануне и показывающий относительную силу раздражителей.
Последнее условие - это просто повторение опытов с угасанием на одном и том же животном. По мере повторения этих опытов число раздражений до окончательного угасания, при прочих равных условиях, становится все меньше и меньше, пока, наконец, полное угасание не наступает уже после одного неподкрепления условного рефлекса, по крайней мере у части животных.
Исключительный интерес в факте угасания привязывается к следующему обстоятельству. Падение рефлекса при угасании обнаруживается не только на том условном рефлексе, который непосредственно подвергся процедуре угашения (первично угашейный рефлекс), но и на других, не тронутых, так сказать, прямо этой процедурой (вторично угашенные рефлексы), и не только на других, связанных с одним и тем же безусловным раздражителем (однородных, по нашей терминологии), но и на выработанных при помощи других безусловных раздражителей (разнородных), и, наконец, при высших степенях угасания, даже на самих безусловных. Так, у одной собаки определенное количество соляной кислоты давало в среднем около 6 куб. см слюны. То же количество соляной кислоты после значительного угашения условного кожно-механического кислотного рефлекса дало только 3.8 куб. см (опыт И. Я. Перельцвейга). Наиболее резкий факт - это вторичное угасание в большей или меньшей степени однородных условных рефлексов; он у нас и обследован более подробно.
Для всех случаев вторичного угасания имеет первенствующее значение степень угашения. Очень сильное угашение многое сравнивает, стирает мелкие отличия. Пмеренном же угашении выступают многие подробности. Принимая остальные условия равными, можно сказать, что последующий эффект угашения одного условного рефлекса на другие, однородные с ним, определяется относительной физиологической силой этих последних. А эта сила в свою очередь зависит от возраста рефлекса, от постоянных или редких, или даже прекратившихся подкреплений его, от частоты предшествовавших угощений и, наконец, от предварительного подкрепления в данный экспериментальный сеанс. Чем Чем больше физиологическая сила данного рефлекса сравнительно с силой первично угашаемого или угашенного рефлекса, тем он менее подвергается вторичному угасанию, и, наоборот, при первичном угашении сильнейшего рефлекса вторично вполне гаснут все слабые.
Вот опыт из работ Бабкина.
У собаки три условных раздражителя кислотного рефлекса: звонок, удары метронома и механическое раздражение кожи.
Условное раздражение продолжается 30 секунд.
Опыт, произведенный накануне и показывающий относительную силу раздражителей.

 Пр. При угашении условного раздражителя средней силы более слабый оказался также совершенно угашениым, а более сильный еще обнаружил некоторое действие.
То же самое выступает и при угашении комплексных (т. е. состоящих из нескольких различных агентов) условных раздражителей и их составляющих. При угашении комплексного раздражителя угашаются и его составляющие. При двух равных составляющих при отдельном угашении одного из них угасает и другой, но суммарный еще несколько действует. При угашении сильнейшего слабейший при отдельной пробе оказывается вполне недействующим, и, наоборот, - при угощении слабейшего сильнейший сохраняет еще свое действие, хотя и ослабленно. При угашении сильнейшего, вполне маскирующего в отдельной пробе слабейший (об отношении агентов в условных комплексных раздражителях будет особая речь в одной из последующих лекций), угасает вполне и суммарный.
Для понимания процесса угасания очень важно следующее особенное отношение. Мы имеем слабейший составляющий раздражитель, вполне замаскированный сильнейшим, т. е. при отдельной пробе не действующий, и тем не менее, когда его повторяют отдельно, не сопровождая безусловным раздражителем, и повторяют несколько раз, оказывается угашенным не только сильнейший составляющий, но и суммарный.
Вот один из таких опытов из работы Перельцвейга.
Раздражение условными кислотными раздражителями: комплексным (механическое раздражение кожи одновременно с температурным в и отдельными составляющими в продолжение минуты.
Пр. При угашении условного раздражителя средней силы более слабый оказался также совершенно угашениым, а более сильный еще обнаружил некоторое действие.
То же самое выступает и при угашении комплексных (т. е. состоящих из нескольких различных агентов) условных раздражителей и их составляющих. При угашении комплексного раздражителя угашаются и его составляющие. При двух равных составляющих при отдельном угашении одного из них угасает и другой, но суммарный еще несколько действует. При угашении сильнейшего слабейший при отдельной пробе оказывается вполне недействующим, и, наоборот, - при угощении слабейшего сильнейший сохраняет еще свое действие, хотя и ослабленно. При угашении сильнейшего, вполне маскирующего в отдельной пробе слабейший (об отношении агентов в условных комплексных раздражителях будет особая речь в одной из последующих лекций), угасает вполне и суммарный.
Для понимания процесса угасания очень важно следующее особенное отношение. Мы имеем слабейший составляющий раздражитель, вполне замаскированный сильнейшим, т. е. при отдельной пробе не действующий, и тем не менее, когда его повторяют отдельно, не сопровождая безусловным раздражителем, и повторяют несколько раз, оказывается угашенным не только сильнейший составляющий, но и суммарный.
Вот один из таких опытов из работы Перельцвейга.
Раздражение условными кислотными раздражителями: комплексным (механическое раздражение кожи одновременно с температурным в и отдельными составляющими в продолжение минуты.
 До сих пор я говорил о слабом или значительном учащении, о степени угашения, но этот пункт требует особого добавления. Интенсивность угашения определяется не только степенью уменьшения условного рефлекса, на которой мы остановились, не только окончательным нулем эффекта, она может нарастать и дальше; мы имеем, так сказать, дальнейшее невидимое угашение. Повторяя сделавшийся недействительным при угашении условный раздражитель, мы углубляем, усиливаем угасание.
После констатирования этого факта исчезает странность вышеприведенного опыта, где недействительный условный раздражитель, повторенный изолированно несколько раз, оказал влияние на следующие за ним раздражители. Таким образом при опытах с угощением необходимо обращать внимание на глубину угасания. Как она определяется за нулем, будет показано в связи со следующим пунктом, к которому мы теперь переходим.
Что делается дальше с угашенным рефлексом? Как он опять получает свое потерянное действие?
Угашенный рефлекс, предоставленный сам себе, без малейшего воздействия со стороны экспериментатора, спустя то или другое время снова приобретает свое действие.
Это, конечно, не относится к только что образующимся условным рефлексам, еще очень слабым и непостоянным. Эти последние в некоторых случаях после угасания для своего восстановления будут нуждаться в новых подкреплениях, т. е. в комбинировании их с безусловным рефлексом. Все же выработанные рефлексы непременно после угасания снова сами собой делаются действительными. И одним из показателей глубины угасания и будет тот период времени, который при всех прочих равных условиях будет требоваться для возвращения угашенному рефлексу его прежнего эффекта. Самопроизвольное восстановление угашенного рефлекса происходит с очень различной быстротой - от нескольких минут до нескольких часов.
Представляю примеры.
Раздражение мясным порошком на расстоянии через каждые 3 минуты без подкрепления (из работы Бабкина).
До сих пор я говорил о слабом или значительном учащении, о степени угашения, но этот пункт требует особого добавления. Интенсивность угашения определяется не только степенью уменьшения условного рефлекса, на которой мы остановились, не только окончательным нулем эффекта, она может нарастать и дальше; мы имеем, так сказать, дальнейшее невидимое угашение. Повторяя сделавшийся недействительным при угашении условный раздражитель, мы углубляем, усиливаем угасание.
После констатирования этого факта исчезает странность вышеприведенного опыта, где недействительный условный раздражитель, повторенный изолированно несколько раз, оказал влияние на следующие за ним раздражители. Таким образом при опытах с угощением необходимо обращать внимание на глубину угасания. Как она определяется за нулем, будет показано в связи со следующим пунктом, к которому мы теперь переходим.
Что делается дальше с угашенным рефлексом? Как он опять получает свое потерянное действие?
Угашенный рефлекс, предоставленный сам себе, без малейшего воздействия со стороны экспериментатора, спустя то или другое время снова приобретает свое действие.
Это, конечно, не относится к только что образующимся условным рефлексам, еще очень слабым и непостоянным. Эти последние в некоторых случаях после угасания для своего восстановления будут нуждаться в новых подкреплениях, т. е. в комбинировании их с безусловным рефлексом. Все же выработанные рефлексы непременно после угасания снова сами собой делаются действительными. И одним из показателей глубины угасания и будет тот период времени, который при всех прочих равных условиях будет требоваться для возвращения угашенному рефлексу его прежнего эффекта. Самопроизвольное восстановление угашенного рефлекса происходит с очень различной быстротой - от нескольких минут до нескольких часов.
Представляю примеры.
Раздражение мясным порошком на расстоянии через каждые 3 минуты без подкрепления (из работы Бабкина).
 Другой опыт (из работ М. И. Эльяссона). Раздражение мясным порошком на расстоянии через каждые 10 минут без подкрепления.
Другой опыт (из работ М. И. Эльяссона). Раздражение мясным порошком на расстоянии через каждые 10 минут без подкрепления.
 Наш сегодняшний опыт c угасанием установили 23 минуты тому назад. Пробуем опять действовать ударами метронома. Мы получаем:
Наш сегодняшний опыт c угасанием установили 23 минуты тому назад. Пробуем опять действовать ударами метронома. Мы получаем:
 Значит, произошло самопроизвольное восстановление в известной степени.
Столь различная скорость восстановленишенного рефлекса зависит от нескольких условий. Прежде всего и чаще всего ее определяет степень, глубина угасания. Затем очень большое значение имеет индивидуальность животных, тип их нервной системы. Заметное действие оказывает интенсивность условного рефлекса. И, наконец, значительное влияние имеет и то, как часто повторяются опыты с угасанием.
Возврат угашенного рефлекса к обычному действию может быть ускорен. Для этого нужно применить тот безусловный рефлекс, при помощи которого образован данный условный рефлекс или один, или совместно с угашенным условным раздражителем, т. е. подкрепить последний. Этот прием тем скорее достигает цели, чем менее глубоко угашен условный рефлекс. При слабом угашении достаточно однократного применения приема, при очень глубоком требуется многократное. Это будет вторым способом определения глубины угасания.
Вопрос о том: скорее ли или одинаково быстро возвращается действие угашенного условного раздражителя в случае применения безусловного рефлекса одного или совместно с условным раздражителем, - оставался до сих пор без точного исследования. Мы занялись им серьезно только теперь.
Весь переданный фактический материал об угасании, может быть несколько обременивший ваше внимание как не объединенный общей точкой зрения, постепенно решал для нас важнейший вопрос: что же такое то явление, которое мы назвали угасанием? Мы должны были признать его за особый вид торможения, так как другие возможные толкованиявления нашими фактами решительно устранялись.
Чт Что угасание не есть разрушение условного рефлекса, разрыв образовавшейся нервной связи, бесспорно доказывается фактом самопроизвольного восстановления угашеного рефлекса по истечении того или другого срока.
После этого оставалась еще возможность думать об утомлении той или другой части нервно-секреторного аппарата, который участвует в наших рефлексах. Но точно так же, как предположение о разрушении рефлекса, и всякая мысль об утомлении при угасании должна быть отброшена ввиду наших фактов. Что ни о каком утомлении собственно секреторных элементов не может быть речи, явствует из всего нашего фактического материала, так как постоянно подкрепляемые условные рефлексы дают огромные количества слюны, сколько бы раз они ни повторялись; да и восстановление угашенного рефлекса достигается лишним разом секреторной деятельности при применении безусловного раздражителя, притом же секреторная работа угашаемого рефлекса часто совершенно ничтожна. Но угасание не может быть утомлением и нервных элементов. Прошу вспомнить опыт, когда один из компонентов условного раздражителя кожи, именно слабейший - температурный, сам по себе в отдельности не производивший ни малейшего положительного эффекта, после многократного повторения, не сопровождаемого безусловным раздражителем, т. е. при процедуре угашения, повел к угасанию более сильного, механического, компонента и даже самого комплексного раздражителя. Не было никакой положительной деятельности. Следовательно, что же могло утомиться? К тому же повторно раздражался температурный нервный аппарат кожи со всеми его составными элементами, а утомился бы (если признать здесь утомление) неработавший механический.
Таким образом путем исключения мы приходим к заключе нию, что то, что мы назвали угасанием, есть тормозной процесс, торможение. С этой точки зрения легко понимаются почти все факты, сообщенные выше относительно угасания.
Наблюдаемые иногда при угасании самопроизвольные колебания ритмического характера легко понимаются как явление борьбы, уравновешивания между раздражительным и тормозным процессами. Также понятно и влияние индивидуальности, так как мы из нашей жизни хорошо знаем, как сильно колеблется тормозная способность нервной системы у разных людей, и чему массу примеров на наших животных мы увидим в дальнейшем изложении. Ясно, что чем сильнее условный рефлекс, т. е. чем интенсивнее раздражительный процесс, тем большее напряжение тормозного процесса, тем большее время потребуется, чтобы преодолеть первый процесс. Так как с повторением неподкрепляемого условного раздражителя торможение должно суммироваться, то естественно, что чем через меньшие промежутки времени происходит повторение, тем в более короткий срок достигается максимальное торможение. Не представляет трудности понимание и того, что при повторении опытов с угасанием угасание происходит все скорее, потому что повторение, практика, усиливает тормозной процесс, что мы опять знаем из ежедневных наблюдений над нами самими и с чем мы часто будем встречаться при дальнейшем изучении условных рефлексов. Влияние угасания одного условного рефлекса на другие не только на однородные, но и на разнородные и даже на безусловный рефлекс должно найти свое объяснение в признании факта распространения тормозного процесса из исходного пункта по массе мозга, что займет наше внимание основательно в одной из следующих лекций.
Но все же остается еще один факт из вышеперечисленных относительно угасания, которым мы должны заняться подробно сейчас же, так как до сих пор он является совершенно темным для нас. Это - те взмахи на правильной линии угасания, которые появляются весьма часто в связи с случайными раздражениями, возникающими в окружающей среде. Стоит раздаться какомунибудь постороннему звуку, измениться общему освещению комнаты в ту или другую сторону, и т. д., и мы имеем резкое нарушение развивающегося угасания, быстрое усиление условного рефлекса. С тем же успехом, конечно, действует масса всех тех посторонних раздражении, которые применяем мы сами для проверки этого влияния. Образцы этих опытов будут приведены ниже. Сюда же относится один из фактов, который очень долго оставался загадочным для нашей лаборатории. Мы угощаем натуральный условный рефлекс на мясной порошок. Сам по себе он восстановится через 1 - час, как показывает контрольный опыт. Но мы сейчас же, как только получил нуль при угасании, вливаем собаке в рот раствор кислоты и по прекращении кислотного слюноотделения (всего спустя 5 минут после нуля угасания) снова применяем мясной порошок на расстоянии как условный пищевой раздражитель. Перед нами почти полный натуральный условный рефлекс. Как это могло произойти, когда условные рефлексы носят отчетливый специфический характер, т. е. определенный раздражитель обусловливает только определенную реакцию? Здесь же угашенный условный пищевой рефлекс возвращает свой эффект благодаря применению агента совершенно другого, кислотного рефлекса. Кислотный рефлекс резко отличается от пищевого и по составу слюны и по двигательной реакции. Итак, что это такое? Фактически --это устранение тормозного процесса. Во всех только что приведенных случаях это устранение носит одну общую черту - оно временное, существует только до тех пор, пока существует вызвавший его раздражитель или продолжается его видимое либо невидимое последействие. Вот вам относящийся сюда интересный эпизод из жизни нашей лаборатории. Члены нашей лаборатории резко разошлись в суждениях относительно восстанавливающего действия кислотного безусловного рефлекса на угашенный пищевой. Одни убежденно удостоверяли этот факт, другие так же решительно его отрицали. Что же верно? Как часто случается в жизни и в науке, оказались правы обе стороны. Опыты И. В. Завадского вполне разъяснили дело. Основание для различных результатов в руках у разных экспериментаторов лежало в просматриваемой ими разнице условий их опытов; одни испытывали угашенный условный пищевой рефлекс сейчас же или в ближайшие минуты, после того как прекратилось слюноотделение от вливания кислоты, другие же - спустя более значительный срок. Принимая это во внимание, Завадский в одном и том же опыте получил и те и другие факты.
Вот один из таких опытов.
Значит, произошло самопроизвольное восстановление в известной степени.
Столь различная скорость восстановленишенного рефлекса зависит от нескольких условий. Прежде всего и чаще всего ее определяет степень, глубина угасания. Затем очень большое значение имеет индивидуальность животных, тип их нервной системы. Заметное действие оказывает интенсивность условного рефлекса. И, наконец, значительное влияние имеет и то, как часто повторяются опыты с угасанием.
Возврат угашенного рефлекса к обычному действию может быть ускорен. Для этого нужно применить тот безусловный рефлекс, при помощи которого образован данный условный рефлекс или один, или совместно с угашенным условным раздражителем, т. е. подкрепить последний. Этот прием тем скорее достигает цели, чем менее глубоко угашен условный рефлекс. При слабом угашении достаточно однократного применения приема, при очень глубоком требуется многократное. Это будет вторым способом определения глубины угасания.
Вопрос о том: скорее ли или одинаково быстро возвращается действие угашенного условного раздражителя в случае применения безусловного рефлекса одного или совместно с условным раздражителем, - оставался до сих пор без точного исследования. Мы занялись им серьезно только теперь.
Весь переданный фактический материал об угасании, может быть несколько обременивший ваше внимание как не объединенный общей точкой зрения, постепенно решал для нас важнейший вопрос: что же такое то явление, которое мы назвали угасанием? Мы должны были признать его за особый вид торможения, так как другие возможные толкованиявления нашими фактами решительно устранялись.
Чт Что угасание не есть разрушение условного рефлекса, разрыв образовавшейся нервной связи, бесспорно доказывается фактом самопроизвольного восстановления угашеного рефлекса по истечении того или другого срока.
После этого оставалась еще возможность думать об утомлении той или другой части нервно-секреторного аппарата, который участвует в наших рефлексах. Но точно так же, как предположение о разрушении рефлекса, и всякая мысль об утомлении при угасании должна быть отброшена ввиду наших фактов. Что ни о каком утомлении собственно секреторных элементов не может быть речи, явствует из всего нашего фактического материала, так как постоянно подкрепляемые условные рефлексы дают огромные количества слюны, сколько бы раз они ни повторялись; да и восстановление угашенного рефлекса достигается лишним разом секреторной деятельности при применении безусловного раздражителя, притом же секреторная работа угашаемого рефлекса часто совершенно ничтожна. Но угасание не может быть утомлением и нервных элементов. Прошу вспомнить опыт, когда один из компонентов условного раздражителя кожи, именно слабейший - температурный, сам по себе в отдельности не производивший ни малейшего положительного эффекта, после многократного повторения, не сопровождаемого безусловным раздражителем, т. е. при процедуре угашения, повел к угасанию более сильного, механического, компонента и даже самого комплексного раздражителя. Не было никакой положительной деятельности. Следовательно, что же могло утомиться? К тому же повторно раздражался температурный нервный аппарат кожи со всеми его составными элементами, а утомился бы (если признать здесь утомление) неработавший механический.
Таким образом путем исключения мы приходим к заключе нию, что то, что мы назвали угасанием, есть тормозной процесс, торможение. С этой точки зрения легко понимаются почти все факты, сообщенные выше относительно угасания.
Наблюдаемые иногда при угасании самопроизвольные колебания ритмического характера легко понимаются как явление борьбы, уравновешивания между раздражительным и тормозным процессами. Также понятно и влияние индивидуальности, так как мы из нашей жизни хорошо знаем, как сильно колеблется тормозная способность нервной системы у разных людей, и чему массу примеров на наших животных мы увидим в дальнейшем изложении. Ясно, что чем сильнее условный рефлекс, т. е. чем интенсивнее раздражительный процесс, тем большее напряжение тормозного процесса, тем большее время потребуется, чтобы преодолеть первый процесс. Так как с повторением неподкрепляемого условного раздражителя торможение должно суммироваться, то естественно, что чем через меньшие промежутки времени происходит повторение, тем в более короткий срок достигается максимальное торможение. Не представляет трудности понимание и того, что при повторении опытов с угасанием угасание происходит все скорее, потому что повторение, практика, усиливает тормозной процесс, что мы опять знаем из ежедневных наблюдений над нами самими и с чем мы часто будем встречаться при дальнейшем изучении условных рефлексов. Влияние угасания одного условного рефлекса на другие не только на однородные, но и на разнородные и даже на безусловный рефлекс должно найти свое объяснение в признании факта распространения тормозного процесса из исходного пункта по массе мозга, что займет наше внимание основательно в одной из следующих лекций.
Но все же остается еще один факт из вышеперечисленных относительно угасания, которым мы должны заняться подробно сейчас же, так как до сих пор он является совершенно темным для нас. Это - те взмахи на правильной линии угасания, которые появляются весьма часто в связи с случайными раздражениями, возникающими в окружающей среде. Стоит раздаться какомунибудь постороннему звуку, измениться общему освещению комнаты в ту или другую сторону, и т. д., и мы имеем резкое нарушение развивающегося угасания, быстрое усиление условного рефлекса. С тем же успехом, конечно, действует масса всех тех посторонних раздражении, которые применяем мы сами для проверки этого влияния. Образцы этих опытов будут приведены ниже. Сюда же относится один из фактов, который очень долго оставался загадочным для нашей лаборатории. Мы угощаем натуральный условный рефлекс на мясной порошок. Сам по себе он восстановится через 1 - час, как показывает контрольный опыт. Но мы сейчас же, как только получил нуль при угасании, вливаем собаке в рот раствор кислоты и по прекращении кислотного слюноотделения (всего спустя 5 минут после нуля угасания) снова применяем мясной порошок на расстоянии как условный пищевой раздражитель. Перед нами почти полный натуральный условный рефлекс. Как это могло произойти, когда условные рефлексы носят отчетливый специфический характер, т. е. определенный раздражитель обусловливает только определенную реакцию? Здесь же угашенный условный пищевой рефлекс возвращает свой эффект благодаря применению агента совершенно другого, кислотного рефлекса. Кислотный рефлекс резко отличается от пищевого и по составу слюны и по двигательной реакции. Итак, что это такое? Фактически --это устранение тормозного процесса. Во всех только что приведенных случаях это устранение носит одну общую черту - оно временное, существует только до тех пор, пока существует вызвавший его раздражитель или продолжается его видимое либо невидимое последействие. Вот вам относящийся сюда интересный эпизод из жизни нашей лаборатории. Члены нашей лаборатории резко разошлись в суждениях относительно восстанавливающего действия кислотного безусловного рефлекса на угашенный пищевой. Одни убежденно удостоверяли этот факт, другие так же решительно его отрицали. Что же верно? Как часто случается в жизни и в науке, оказались правы обе стороны. Опыты И. В. Завадского вполне разъяснили дело. Основание для различных результатов в руках у разных экспериментаторов лежало в просматриваемой ими разнице условий их опытов; одни испытывали угашенный условный пищевой рефлекс сейчас же или в ближайшие минуты, после того как прекратилось слюноотделение от вливания кислоты, другие же - спустя более значительный срок. Принимая это во внимание, Завадский в одном и том же опыте получил и те и другие факты.
Вот один из таких опытов.
 Как видите, 7 минут спустя по окончании кислотного слюнаотделения восстановление минимальное и только на одной железе; спустя же только 40 секунд восстановление очень значительное и на обеих железах.
Временный характер восстановления обнаруживается и при других раздражителях, о которых у нас сейчас идет речь и которые обусловливают неправильные взмахивая обычно правильно опускающейся линии угасания или уже при полном угасании.
Вот другой опыт Завадского на другой собаке.
Как видите, 7 минут спустя по окончании кислотного слюнаотделения восстановление минимальное и только на одной железе; спустя же только 40 секунд восстановление очень значительное и на обеих железах.
Временный характер восстановления обнаруживается и при других раздражителях, о которых у нас сейчас идет речь и которые обусловливают неправильные взмахивая обычно правильно опускающейся линии угасания или уже при полном угасании.
Вот другой опыт Завадского на другой собаке.
 Как видно, и здесь восстановляют угашенный рефлекс как наличные раздражители (первые два раздражителя), так и латентное последействие раздражителя (последний раздражитель).
В только что приведенных опытах восстанавливающее действие сводится на немногие минуты, соответственно краткости действия наличных раздражителей и их последействия. Но при некоторых специальных посторонних рефлексах, упомянутых в отделе о внешнем торможении и имеющих затяжной характер, соответственно и восстановляющее действие дает себя знать во время всего опыта и, таким образом, становится невозможным получить правильную линию угасания и вообще полное и прочное угасание.
Далее надо остановиться на следующем важном обстоятельстве, касающемся того же пункта. На всем протяжении работы мы много раз наблюдали одновременное существование нескольких различных рефлексов; при этом, конечно, выступает и их взаимодействие, то дающее перевес одному или другому из них, то обусловливающее их взаимную нейтрализацию. Например, когда делают из механического раздражения кожи условный раздражитель, то часто встречаются при этом с противодействием безусловных кожных рефлексов в роде чесательного движения животного, отряхивательного и тому подобных движений, вследствие чего у некоторых животных, правда редких, становится невозможным достигнуть постоянного ровного условного кожно-механического рефлекса. То же самое получается и при некоторых высоких тонах, вызывающих у некоторых собак резкую отрицательную двигательную реакцию. А это значит, что безусловные раздражители, как агенты из группы внешнего торможения, постоянно более или менее тормозят условный рефлекс. Но тогда нужно ожидать, что такие упорствующие безусловные рефлексы тем больше, тем чаще будут нарушать правильный ход угасания, так как процесс угасания гораздо лабильнее процесса раздражения. Как примеры таких случаев, так и их более подробный экспериметальный анализ будут представлены в одной изследующих лекций.
Все приведенные данные позволяют нам называть изучаемое явление - временное восстановление угашаетого или угашенного рефлекса, временное устранение торможения - растормаживанием, каковым термином мы отныне и будем пользоваться постоянно.
Но теперь возникает серьезный вопрос: какая же разница между восстановлением угашенного условного рефлекса при помощи его безусловного и тем, что сейчас мы назвали растормаживанием? Фактическую разницу нетрудно видеть. В одном случае восстановление стойкое, в другом временное, в одном оно наступает при помощи специального агента, с которым предварительно связан условный агент, в другом это происходит под влиянием совершенно посторонних раздражений. Что касается существенного основания этого различия, то о нем составить себе отчетливое представление при настоящем фактическом материале трудно или даже прямо невозможно. Ведь и в первом случае дело идет только об устранении торможения, так как всякая мысль о разрушении условного рефлекса при угасании исключена фактом рано или поздно, но непременно наступающего самопроизвольного восстановления угашенного условного рефлекса. Таким образом приходится признать какую-то разницу в самом механизме устранения тормозного процесса в обоих этих случаях.
Как представлять себе внутренний механизм растормаживания?
Конечно, раз мы не знаем, что такое торможение, так же как и само раздражение и их взаимное отношение, не может быть и речи о фундаментальном объяснении растормаживания. Мы находим только нужным здесь обратить внимание на следующее сопоставление фактов. Те самые агенты, которые вызываемыми ими посторонними рефлексами тормозят положительные условные рефлексы, - когда эти посторонние рефлекты слабы с самого начала или слабеют при повторении, - производят растормаживание угашенных условных рефлексов. (Относящиеся сюда факты будут приведены в следующей лекции). В таком случае получается, так сказать, схематическое, словесное основание считать и называть растормаживание торможением торможения, как мы ранее и выражались. Но это, конечно, не объяснение.
Итак, главнейший результат всего переданного вам в этой лекции экспериментального материала тот, что положительный условный раздражитель довольно скоро, остро, но все же постепенно может быть превращен в условный отрицательный, тормозной. Мы должны, таким образом, постоянно иметь дело как с положительными условными, так и с отрицательными условными рефлексами.
Как видно, и здесь восстановляют угашенный рефлекс как наличные раздражители (первые два раздражителя), так и латентное последействие раздражителя (последний раздражитель).
В только что приведенных опытах восстанавливающее действие сводится на немногие минуты, соответственно краткости действия наличных раздражителей и их последействия. Но при некоторых специальных посторонних рефлексах, упомянутых в отделе о внешнем торможении и имеющих затяжной характер, соответственно и восстановляющее действие дает себя знать во время всего опыта и, таким образом, становится невозможным получить правильную линию угасания и вообще полное и прочное угасание.
Далее надо остановиться на следующем важном обстоятельстве, касающемся того же пункта. На всем протяжении работы мы много раз наблюдали одновременное существование нескольких различных рефлексов; при этом, конечно, выступает и их взаимодействие, то дающее перевес одному или другому из них, то обусловливающее их взаимную нейтрализацию. Например, когда делают из механического раздражения кожи условный раздражитель, то часто встречаются при этом с противодействием безусловных кожных рефлексов в роде чесательного движения животного, отряхивательного и тому подобных движений, вследствие чего у некоторых животных, правда редких, становится невозможным достигнуть постоянного ровного условного кожно-механического рефлекса. То же самое получается и при некоторых высоких тонах, вызывающих у некоторых собак резкую отрицательную двигательную реакцию. А это значит, что безусловные раздражители, как агенты из группы внешнего торможения, постоянно более или менее тормозят условный рефлекс. Но тогда нужно ожидать, что такие упорствующие безусловные рефлексы тем больше, тем чаще будут нарушать правильный ход угасания, так как процесс угасания гораздо лабильнее процесса раздражения. Как примеры таких случаев, так и их более подробный экспериметальный анализ будут представлены в одной изследующих лекций.
Все приведенные данные позволяют нам называть изучаемое явление - временное восстановление угашаетого или угашенного рефлекса, временное устранение торможения - растормаживанием, каковым термином мы отныне и будем пользоваться постоянно.
Но теперь возникает серьезный вопрос: какая же разница между восстановлением угашенного условного рефлекса при помощи его безусловного и тем, что сейчас мы назвали растормаживанием? Фактическую разницу нетрудно видеть. В одном случае восстановление стойкое, в другом временное, в одном оно наступает при помощи специального агента, с которым предварительно связан условный агент, в другом это происходит под влиянием совершенно посторонних раздражений. Что касается существенного основания этого различия, то о нем составить себе отчетливое представление при настоящем фактическом материале трудно или даже прямо невозможно. Ведь и в первом случае дело идет только об устранении торможения, так как всякая мысль о разрушении условного рефлекса при угасании исключена фактом рано или поздно, но непременно наступающего самопроизвольного восстановления угашенного условного рефлекса. Таким образом приходится признать какую-то разницу в самом механизме устранения тормозного процесса в обоих этих случаях.
Как представлять себе внутренний механизм растормаживания?
Конечно, раз мы не знаем, что такое торможение, так же как и само раздражение и их взаимное отношение, не может быть и речи о фундаментальном объяснении растормаживания. Мы находим только нужным здесь обратить внимание на следующее сопоставление фактов. Те самые агенты, которые вызываемыми ими посторонними рефлексами тормозят положительные условные рефлексы, - когда эти посторонние рефлекты слабы с самого начала или слабеют при повторении, - производят растормаживание угашенных условных рефлексов. (Относящиеся сюда факты будут приведены в следующей лекции). В таком случае получается, так сказать, схематическое, словесное основание считать и называть растормаживание торможением торможения, как мы ранее и выражались. Но это, конечно, не объяснение.
Итак, главнейший результат всего переданного вам в этой лекции экспериментального материала тот, что положительный условный раздражитель довольно скоро, остро, но все же постепенно может быть превращен в условный отрицательный, тормозной. Мы должны, таким образом, постоянно иметь дело как с положительными условными, так и с отрицательными условными рефлексами.
Лекция пятая. 2) внутреннее торможение: 6) условное торможение
Мм. гг.! Вся предшествующая лекции была заполнена изучением первого случая внутреннего торможения - угасания. Это кратковременное превращение положительного условного раздражителя (раз он через небольшие, минутные, промежутки повторялся без сопровождения его безусловным рефлексом) в отрицательный, тормозной раздражитель. Теперь я перехожу ко второму случаю внутреннего торможения, так же подробно нами изученному. Вот основной опыт. Мы имеем прочно выработанный положительный условный раздражитель. Затем мы прибавляем к нему новый агент и эту комбинацию повторно воспроизводим, хотя бы и с большими промежутками, через часы и даже дни, не сопровождая безусловным раздражителем. Тогда эта комбинация постепенно делается недействительной, т. е. наш условный раздражитель в соединении с новым агентом постепенно теряет свое положительное действие, хотя применяемый в то же время отдельно и всегда подкрепляемый остается в полной силе. Это явление до сих пор в наших исследованиях называется условным торможением. Обозначение это нельзя назвать удачным, потому что и угасание есть тоже условное торможение, так как возникает, вырабатывается при определенном условии. Оправданием для такого названия служит история изучения факта этого торможения. Так как здесь дело идет о соучастии прибавочного агента, то на первых порах нами это явление было смешано с внешним торможением, и только потом, когда стала отчетливой натура этого явления как внутреннего торможения, оно было отличено от внешнего прилагательным «условное». Как видим дальше, для него более соответствующим обозначением был бы термин «дифференцировочное торможение". Выработка условного торможения представляет особенный интерес в том отношении, что, иллюстрируя многостороннюю сложность явлений, с которой мы здесь имеем дело, она, однако, свидетельствует вместе с тем об экспериментальной возможности удовлетворительно разобраться в этой сложности. Я поэтому считаю полезным остановиться на этом пункте несколько подробнее. При выработке условного торможения прежде всего выступоют очень своеобразные отношения в связи с моментом встречи положительного условного раздражителя и прибавочного агента. Если этот агент пускается в ход за немного секунд (3-5) до начала условного раздражителя (обыкновенный у нас случай) или одновременно с ним, или даже несколько секунд после начала последнего, а затем при всех этих вариациях продолжается вместе с ним, то условное торможение развивается более или менее легко. Если прибавочный агент прекращается, как только начинается условный раздражитель, то в некоторых случаях условное торможение наступает уже с заметной трудностью для нервной системы животного, что явствует из беспокойства животного и разных его оборонительных реакций. Если же вставляется промежуток в несколько секунд между прибавочным агентом и условным раздражителем, незаметно никакого эффекта. Когда промежуток доходит до 10 секунд (наиболее обыкновенный случай), прибавочный агент сам становится условным раздражителем, как это было приведено выше при описании процедуры образования условных рефлексов (вторичный условный рефлекс). Только если берется исключительно сильный прибавочный агент, например автомобильный гудок, условное торможение может происходить еще и промежутке в 20 секунд. Вот относящийся сюда опыт из работы Фролова. Автомобильный гудок, действовавший 10 секунд и отделенный от условного пищевого раздражителя ударов метронома паузой в 10 секунд, при первом его применении нисколько не повлиял на величину условного рефлекса. Но затем, отделенный паузой даже в 20 секунд, он, при повторении в такой комбинации с неподкрепленным условным раздражителем, постепенно повел к уменьшению величины этого последнего. О п ы т 28 XII 1924. Автомобильный гудок применяется только во второй раз. О п ы т 21 I 1925. Автомобильный гудок применяется в три- инадцатый раз.
О п ы т 21 I 1925. Автомобильный гудок применяется в три- инадцатый раз.
 Вообще вышеуказанные промежутки несколько колеблются в связи с интенсивностью прибавочных агентов.
Таким образом в данном случае мы имеем очень интересную встречу процессов раздражения и торможения. Как понимать столь различный ход явлений при такой, повидимому незначительной, разнице в условиях?
Мы остановились на следующем понимании приведенных фактов, опираясь на соответствие его с другими нам известными фактами. Когда прибавочный агент и условный раздражитель совпадают во времени (точно или приблизительно, т. е. в виде свежих следов прибавочного агента), они вместе образуют как бы новый особый раздражитель, отчасти сходный с условным раздражителем, отчасти от него отличный. А мы увидим в седьмой лекции, как подобные раздражители, например близкие тоны или разные места кожи и т. п., когда один из них при помощи соответственной процедуры становится условным раздражителем, являются сперва сами собой тоже условно действующими, а потом, при систематическом повторении их без подкрепления, теряют это свое действие, превращаясь тормозные раздражители. Это было бы в нашем объясняемом случае вазой образования условного торможения. Когда же прибавочный агент более или менее отодвигается во времени от условного раздражителя, т. е. когда слитие их в одно раздражение затруднено или совсем не осуществляется, то имеет место обычный процесс образования из прибавочного агента нового условного раздражителя, причем старый условный раздражитель имеетеперь совершенно то же значение, как безусловный раздражитель при обыкновенном образовании главной массы условных рефлексов. При таком взгляде легко понимать, почему промежуток между прибавочным агентом и условным раздражителем при развитии условного торможения может быть тем длиннее, чем сильнее прибавочной агент. Сильный агент оставляет после себя более длинное последействие, которое и при более значительном промежутке может сливаться с условным раздражителем, образуя с ним общий и особый нервный акт. Во всяком случае, правильно или неправильно наше объяснение, сами явления представляют ободыяющий экспериментатора пример уловимой закономерности в столь сложном случае центральной нервной деятельности.
Но рядом с этим почти постоянным положением дела надо упомянуть об очень редких случаях как на нетронутых животных, так и на оперированных в области больших полушарий, когда при очевидно повышенной возбудимости нервной системы и при полном совпадении во времени прибавочного агента и условного раздражителя резко и длительно выступает не условное торможение, а образование вторичного условного рефлекса, а затем может даже существовать одновременно и то и другое.
О п ы т Н. А. Кашерининовой. Механическое раздражение кожи - условный кислотный раздражитель, удары метронома прибавочный агент в тормозной комбинации. При двадцать пятом повторении тормозной комбинации она дала только 3 капли слюны за минуту, при эффекте отдельно применяемого условного раздражителя в 29 капель за минуту. После тридцать четвертого повторения тормозной комбинации удары метронома, испробованные отдельно, вызывали отделен до участия в тормозной комбинации они совершенно не имели слюногонного действия.
Может быть, в очень слабой и мимолетной форме то же встречается и чаще.
Самый ход развития условного торможения также очень разнообразен. Один раз, когда к положительному условному раздражителю впервые прибавляется новый агент, происходит сейчас же уменьшение или даже совершенное исчезание условного действия, которое при повторении комбинации потом восстанавливается, а затем вновь и постепенно идет к нулю. При другом прибавочном агенте, наоборот, дело начинается с более значительного положительного эффекта комбинации сравнительно с обычным действием условного раздражителя, а затем только начинается постепенное падение эффекта комбинации до нуля. При следующей вариации может происходить смена первоначального уменьшения последующим увеличением эффекта комбинации сравнительно с нормой условного раздражителя, и лишь потом наступает постепенное уменьшение до прочного нуля. Что это значит и с чем связаны эти вариации? Это связано с интенсивностью постороннего рефлекса, вызываемого прибавочным агентом. Первоначальное и сейчас же наступающее уменьшение эффекта комбинации есть бесспорно процесс внешнего торможения. Один прибавочный агент, положим, вызывает сильный исследовательский рефлекс, он будет с места тормозить положительный условный рефлекс. Если другой прибавочный агент сопровождается только слабым исследовательским рефлексом, то дело начнется с увеличенного эффекта комбинации, и это бесспорно есть процесс растормаживания, так как при наших обычных условных рефлексах перед началом действия условного раздражителя есть фаза внутреннего торможения, которая и устраняется слабым исследовательским рефлексом. С этой фазой мы познакомимся в следующей лекции при изучении третьего случая внутреннего торможения. Смена первоначального уменьшения эффекта комбинации увеличением его имеет свое основание в постепенном ослаблении постороннего рефлекса при повторном применении прибавочного агента; рефлекс этот сперва тормозит Условный рефлекс, а при своем ослаблении растормаживает его, как это мы увидим в конце сегодняшней же лекции.
Всевозможные агенты внешнего мира могут служить в качестве прибавочных для развития условного торможения, но, конечно, только те, для которых есть воспринимающие поверхности у данного животного. Примеров этих агентов мы будем видеть немало в протоколах нижеприводимых опытов.
Прибавочные агенты, как уже упомянуто выше, могут служить для условного торможения не только как совпадающие во времени с условным раздражителем, но и в виде следов, обыкновенно возможно ранних, т. е. при непосредственном переходе от прекращенного прибавочного агента к условному раздражителю; лишь при очень сильных агентах и на более поздних следах возможна выработка условного торможения, как я только что указал.
Но когда тормозная комбинация вполне выработалась, тогда возможно и очень большое отставление во времени прибавочного агента от условного раздражителя, даже до минуты, и все же тормозное значение комбинации сохраняется.
Имеются опыты и с временем, как агентом внутреннего торможения. Вот относящийся сюда опыт К. Н. Краышковского.
У собаки условное торможение кислотного рефлекса (тон условный кислотный раздражитель, механическое раздражение кожи - прибавочный агент) постоянно с некоторой целью применялось через 19-20 минут после последнего вливания кислоты. Тогда образовалось такое положение дела: в девятнадцатуюдвадцатую минуту после вливания кислоты условный раздражитель - тон - и без присоединения к нему механического раздражения кожи давал минимальное отделение слюны или никакого.
Вообще вышеуказанные промежутки несколько колеблются в связи с интенсивностью прибавочных агентов.
Таким образом в данном случае мы имеем очень интересную встречу процессов раздражения и торможения. Как понимать столь различный ход явлений при такой, повидимому незначительной, разнице в условиях?
Мы остановились на следующем понимании приведенных фактов, опираясь на соответствие его с другими нам известными фактами. Когда прибавочный агент и условный раздражитель совпадают во времени (точно или приблизительно, т. е. в виде свежих следов прибавочного агента), они вместе образуют как бы новый особый раздражитель, отчасти сходный с условным раздражителем, отчасти от него отличный. А мы увидим в седьмой лекции, как подобные раздражители, например близкие тоны или разные места кожи и т. п., когда один из них при помощи соответственной процедуры становится условным раздражителем, являются сперва сами собой тоже условно действующими, а потом, при систематическом повторении их без подкрепления, теряют это свое действие, превращаясь тормозные раздражители. Это было бы в нашем объясняемом случае вазой образования условного торможения. Когда же прибавочный агент более или менее отодвигается во времени от условного раздражителя, т. е. когда слитие их в одно раздражение затруднено или совсем не осуществляется, то имеет место обычный процесс образования из прибавочного агента нового условного раздражителя, причем старый условный раздражитель имеетеперь совершенно то же значение, как безусловный раздражитель при обыкновенном образовании главной массы условных рефлексов. При таком взгляде легко понимать, почему промежуток между прибавочным агентом и условным раздражителем при развитии условного торможения может быть тем длиннее, чем сильнее прибавочной агент. Сильный агент оставляет после себя более длинное последействие, которое и при более значительном промежутке может сливаться с условным раздражителем, образуя с ним общий и особый нервный акт. Во всяком случае, правильно или неправильно наше объяснение, сами явления представляют ободыяющий экспериментатора пример уловимой закономерности в столь сложном случае центральной нервной деятельности.
Но рядом с этим почти постоянным положением дела надо упомянуть об очень редких случаях как на нетронутых животных, так и на оперированных в области больших полушарий, когда при очевидно повышенной возбудимости нервной системы и при полном совпадении во времени прибавочного агента и условного раздражителя резко и длительно выступает не условное торможение, а образование вторичного условного рефлекса, а затем может даже существовать одновременно и то и другое.
О п ы т Н. А. Кашерининовой. Механическое раздражение кожи - условный кислотный раздражитель, удары метронома прибавочный агент в тормозной комбинации. При двадцать пятом повторении тормозной комбинации она дала только 3 капли слюны за минуту, при эффекте отдельно применяемого условного раздражителя в 29 капель за минуту. После тридцать четвертого повторения тормозной комбинации удары метронома, испробованные отдельно, вызывали отделен до участия в тормозной комбинации они совершенно не имели слюногонного действия.
Может быть, в очень слабой и мимолетной форме то же встречается и чаще.
Самый ход развития условного торможения также очень разнообразен. Один раз, когда к положительному условному раздражителю впервые прибавляется новый агент, происходит сейчас же уменьшение или даже совершенное исчезание условного действия, которое при повторении комбинации потом восстанавливается, а затем вновь и постепенно идет к нулю. При другом прибавочном агенте, наоборот, дело начинается с более значительного положительного эффекта комбинации сравнительно с обычным действием условного раздражителя, а затем только начинается постепенное падение эффекта комбинации до нуля. При следующей вариации может происходить смена первоначального уменьшения последующим увеличением эффекта комбинации сравнительно с нормой условного раздражителя, и лишь потом наступает постепенное уменьшение до прочного нуля. Что это значит и с чем связаны эти вариации? Это связано с интенсивностью постороннего рефлекса, вызываемого прибавочным агентом. Первоначальное и сейчас же наступающее уменьшение эффекта комбинации есть бесспорно процесс внешнего торможения. Один прибавочный агент, положим, вызывает сильный исследовательский рефлекс, он будет с места тормозить положительный условный рефлекс. Если другой прибавочный агент сопровождается только слабым исследовательским рефлексом, то дело начнется с увеличенного эффекта комбинации, и это бесспорно есть процесс растормаживания, так как при наших обычных условных рефлексах перед началом действия условного раздражителя есть фаза внутреннего торможения, которая и устраняется слабым исследовательским рефлексом. С этой фазой мы познакомимся в следующей лекции при изучении третьего случая внутреннего торможения. Смена первоначального уменьшения эффекта комбинации увеличением его имеет свое основание в постепенном ослаблении постороннего рефлекса при повторном применении прибавочного агента; рефлекс этот сперва тормозит Условный рефлекс, а при своем ослаблении растормаживает его, как это мы увидим в конце сегодняшней же лекции.
Всевозможные агенты внешнего мира могут служить в качестве прибавочных для развития условного торможения, но, конечно, только те, для которых есть воспринимающие поверхности у данного животного. Примеров этих агентов мы будем видеть немало в протоколах нижеприводимых опытов.
Прибавочные агенты, как уже упомянуто выше, могут служить для условного торможения не только как совпадающие во времени с условным раздражителем, но и в виде следов, обыкновенно возможно ранних, т. е. при непосредственном переходе от прекращенного прибавочного агента к условному раздражителю; лишь при очень сильных агентах и на более поздних следах возможна выработка условного торможения, как я только что указал.
Но когда тормозная комбинация вполне выработалась, тогда возможно и очень большое отставление во времени прибавочного агента от условного раздражителя, даже до минуты, и все же тормозное значение комбинации сохраняется.
Имеются опыты и с временем, как агентом внутреннего торможения. Вот относящийся сюда опыт К. Н. Краышковского.
У собаки условное торможение кислотного рефлекса (тон условный кислотный раздражитель, механическое раздражение кожи - прибавочный агент) постоянно с некоторой целью применялось через 19-20 минут после последнего вливания кислоты. Тогда образовалось такое положение дела: в девятнадцатуюдвадцатую минуту после вливания кислоты условный раздражитель - тон - и без присоединения к нему механического раздражения кожи давал минимальное отделение слюны или никакого.
 Скорость выработки условного торможения, а также и полнота его (абсолютное или относительное торможение) зависят от нескольких условий.
На первом месте здесь надо поставить индивидуальность животного, тип его нервной системы: раздражительный, уравновешенный или легко тормозимый. У некоторых собак развитие торможения затягивается на очень большой срок, и оно никогда не делается полным, у других, наоборот, после очень немногих повторений тормозной комбинации наступает прочное и полное торможение.
Затем отчетливое значение имеет интенсивность прибавочного агента. Так, у собаки (опыт Г. В. Миштовта), имевшей условный кислотный раздражитель из ударов метронома, приложение к коже температуры в 4-5 to для условного торможения начало сопровождаться торможением только с тридцатого раза, а полного торможения не было достигнуто и после 145 раз. Температура в 1Р, примененная с той же целью после четырехмесячного перерыва, дала полное торможение уже после 12 повторений. Но тот или другой результат определяется и отношением интенсивностей прибавочного агента и агента, являющегося условным раздражителем. Так, температура в 45°, приложенная к коже, не образует абсолютной тормозной комбинации с условным раздражителем из ударов метронома, а это достигается легко на том же животном при световом условном раздражителе (опыт Фурсикова).
Наконец обращает на себя внимание факт, что при всех прочих равных условиях первый случай развития условного торможения у данной собаки требует обыкновенно гораздо большего срока, чем последующие.
До сих пор я говорил об условном торможении, не представив доказательств, что это действительно торможение, а не просто исчезание эффекта при систематическом неподкреплении комбинации безусловным раздражителем. Эти доказательства будут приводиться по мере того, как будут передаваться наши материалы, собранные относительно этого предмета. Вот первое.
Чем является тот прибавочный агент, при участии которого происходит условное торможение, какая его окончательная функция? Конечно, вопрос должен быть решен пробой этого агента в других формах опыта. Сам по себе, испытанный одиночно, когда полностью уже развилось условное торможение, он не производит никакого видимого эффекта. Но в комбинации с другими условными раздражителями, с которыми он раньше никогда не применялся, его роль обнаруживается вполне ярко. Эти другие условные раздражители при присоединении к ним прибавочного агента тормозной комбинации сразу испытывают резкое уменьшение эффекта сравнительно с обычным. Это действие выступает не только на однородных рефлексах, т. е. связанных с одним и тем же безусловным, но и на разнородных и даже на безусловном.
Вот соответствующие опыты.
У собаки образованы три условных пищевых рефлекса: на вспыхивание лампочек перед глазами собаки, на вертящийся предмет и на тон cis духового камертона. На условном рефлексе с вертящимся предметом выработано полное условное торможение отдельно: при помощи механического раздражения кожи и ударов метронома (опыт Н. И. Лепорского).
Скорость выработки условного торможения, а также и полнота его (абсолютное или относительное торможение) зависят от нескольких условий.
На первом месте здесь надо поставить индивидуальность животного, тип его нервной системы: раздражительный, уравновешенный или легко тормозимый. У некоторых собак развитие торможения затягивается на очень большой срок, и оно никогда не делается полным, у других, наоборот, после очень немногих повторений тормозной комбинации наступает прочное и полное торможение.
Затем отчетливое значение имеет интенсивность прибавочного агента. Так, у собаки (опыт Г. В. Миштовта), имевшей условный кислотный раздражитель из ударов метронома, приложение к коже температуры в 4-5 to для условного торможения начало сопровождаться торможением только с тридцатого раза, а полного торможения не было достигнуто и после 145 раз. Температура в 1Р, примененная с той же целью после четырехмесячного перерыва, дала полное торможение уже после 12 повторений. Но тот или другой результат определяется и отношением интенсивностей прибавочного агента и агента, являющегося условным раздражителем. Так, температура в 45°, приложенная к коже, не образует абсолютной тормозной комбинации с условным раздражителем из ударов метронома, а это достигается легко на том же животном при световом условном раздражителе (опыт Фурсикова).
Наконец обращает на себя внимание факт, что при всех прочих равных условиях первый случай развития условного торможения у данной собаки требует обыкновенно гораздо большего срока, чем последующие.
До сих пор я говорил об условном торможении, не представив доказательств, что это действительно торможение, а не просто исчезание эффекта при систематическом неподкреплении комбинации безусловным раздражителем. Эти доказательства будут приводиться по мере того, как будут передаваться наши материалы, собранные относительно этого предмета. Вот первое.
Чем является тот прибавочный агент, при участии которого происходит условное торможение, какая его окончательная функция? Конечно, вопрос должен быть решен пробой этого агента в других формах опыта. Сам по себе, испытанный одиночно, когда полностью уже развилось условное торможение, он не производит никакого видимого эффекта. Но в комбинации с другими условными раздражителями, с которыми он раньше никогда не применялся, его роль обнаруживается вполне ярко. Эти другие условные раздражители при присоединении к ним прибавочного агента тормозной комбинации сразу испытывают резкое уменьшение эффекта сравнительно с обычным. Это действие выступает не только на однородных рефлексах, т. е. связанных с одним и тем же безусловным, но и на разнородных и даже на безусловном.
Вот соответствующие опыты.
У собаки образованы три условных пищевых рефлекса: на вспыхивание лампочек перед глазами собаки, на вертящийся предмет и на тон cis духового камертона. На условном рефлексе с вертящимся предметом выработано полное условное торможение отдельно: при помощи механического раздражения кожи и ударов метронома (опыт Н. И. Лепорского).

 B первом опыте к вспыхиванию лампочек впервые было присоединено механическое раздражение кожи, и получилось огромное торможение, хотя и не до нуля. Во втором -- то же сделали удары метронома с тоном cis, впервые с ним встретившиеся.
Итак, агент, служивший для условного торможения одного условного рефлекса, тормозит при первом же применении и другие условные рефлексы, связанные с одним и тем же безусловным рефлексом.
Но то же проявляется и при первом действии этого агента совместно с условными разнородными агентами, т. е. основанными на разных безусловных рефлексах.
У собаки удары метронома - условный пищевой возбудитель, свист - прибавочный агент тормозной комбинации с ударами метронома, механическое раздражение кожи - условный кислотный раздражитель (опыты Бабкина).
B первом опыте к вспыхиванию лампочек впервые было присоединено механическое раздражение кожи, и получилось огромное торможение, хотя и не до нуля. Во втором -- то же сделали удары метронома с тоном cis, впервые с ним встретившиеся.
Итак, агент, служивший для условного торможения одного условного рефлекса, тормозит при первом же применении и другие условные рефлексы, связанные с одним и тем же безусловным рефлексом.
Но то же проявляется и при первом действии этого агента совместно с условными разнородными агентами, т. е. основанными на разных безусловных рефлексах.
У собаки удары метронома - условный пищевой возбудитель, свист - прибавочный агент тормозной комбинации с ударами метронома, механическое раздражение кожи - условный кислотный раздражитель (опыты Бабкина).
 Что касается степени условного торможения на комбинациях рефлексов, то она зависит от интенсивности комплексных рефлексов, которые подвергаются этому торможению.
Вот относящийся сюда опыт из работ Лепорского.
У собаки имелось три отдельных условных пищевых рефлекса: на вертящийся предмет, вспыхивание лампочек и тон. Ко всем им в отдельности был выработан условный тормоз из механического раздражения кожи, причем тормозная комбинация во всех трех случаях не давала ни следа слюны. Действуя все вместе одновременно, эти условные раздражители обусловливали гораздо больший слюноотделительный эффект, чем самый сильный из них в отдельности.
Что касается степени условного торможения на комбинациях рефлексов, то она зависит от интенсивности комплексных рефлексов, которые подвергаются этому торможению.
Вот относящийся сюда опыт из работ Лепорского.
У собаки имелось три отдельных условных пищевых рефлекса: на вертящийся предмет, вспыхивание лампочек и тон. Ко всем им в отдельности был выработан условный тормоз из механического раздражения кожи, причем тормозная комбинация во всех трех случаях не давала ни следа слюны. Действуя все вместе одновременно, эти условные раздражители обусловливали гораздо больший слюноотделительный эффект, чем самый сильный из них в отдельности.
 Условный тормоз, тормозивший до нуля в отдельности каждый условный раздражитель, комплекс из них, как более сильный раздражитель, затормозил только частично
Из приведенных опытов с полным правом следует заключить, что прибавочный агент в случае полного развития условного торможения обусловливает сам по себе тормозящее действие, становится настоящим условным тормозом, каковым термином мы и будем его теперь обозначать.
Понятно, что такое же тормозящее действие имеет и обнаруживает и сама тормозная комбинаичем это действие долго дает себя знать и после того, как прекратилось само раздражение. Это последовательное торможение продолжается минуты и десятки минут и обнаруживается не только на отдельно примененном условном раздражителе тормозной комбинации, но и на всех других условных раздражителях, даже и на разнородных.
Вот относящиеся сюда опыты.
I
У собаки вертящийся предмет - условный пищевой раздражитель, тон в 30 000 колебаний гальтоновского свистка условный тормоз (опыты П. Н. Николаева).
Условный тормоз, тормозивший до нуля в отдельности каждый условный раздражитель, комплекс из них, как более сильный раздражитель, затормозил только частично
Из приведенных опытов с полным правом следует заключить, что прибавочный агент в случае полного развития условного торможения обусловливает сам по себе тормозящее действие, становится настоящим условным тормозом, каковым термином мы и будем его теперь обозначать.
Понятно, что такое же тормозящее действие имеет и обнаруживает и сама тормозная комбинаичем это действие долго дает себя знать и после того, как прекратилось само раздражение. Это последовательное торможение продолжается минуты и десятки минут и обнаруживается не только на отдельно примененном условном раздражителе тормозной комбинации, но и на всех других условных раздражителях, даже и на разнородных.
Вот относящиеся сюда опыты.
I
У собаки вертящийся предмет - условный пищевой раздражитель, тон в 30 000 колебаний гальтоновского свистка условный тормоз (опыты П. Н. Николаева).
 По прекращении тормозной комбинации условный раздражитель из этой комбинации оказался ослабленным десятки минут.
II
У собаки вертящийся предмет - условный кислотный раздражитель, тон - условный пищевой, механическое раздражение кожи - условный тормоз к последнему (опыты Н. П. Понизовского).
По прекращении тормозной комбинации условный раздражитель из этой комбинации оказался ослабленным десятки минут.
II
У собаки вертящийся предмет - условный кислотный раздражитель, тон - условный пищевой, механическое раздражение кожи - условный тормоз к последнему (опыты Н. П. Понизовского).
 Здесь оказался последовательно заторможенным разнородный условный рефлекс.
Так же, как тормозная комбинация, и отдельно примененный условный тормоз обусловливает последовательное торможение.
Теперь еще две подробности.
Последовательное торможение суммируется. Если мы тормозную комбинацию применим не один раз, а последовательно несколько раз, то ее последовательное тормозящее действие становится интенсивнее и тем значительнее, чем более раз мы ее повторяем.
О п ы т. Удары метронома - условный пищевой раздражитель, вертящийся предмет - условный тормоз к нему (опыты
О. М. Чеботаревой).
Здесь оказался последовательно заторможенным разнородный условный рефлекс.
Так же, как тормозная комбинация, и отдельно примененный условный тормоз обусловливает последовательное торможение.
Теперь еще две подробности.
Последовательное торможение суммируется. Если мы тормозную комбинацию применим не один раз, а последовательно несколько раз, то ее последовательное тормозящее действие становится интенсивнее и тем значительнее, чем более раз мы ее повторяем.
О п ы т. Удары метронома - условный пищевой раздражитель, вертящийся предмет - условный тормоз к нему (опыты
О. М. Чеботаревой).
 О пыт на той же собаке на другой день.
О пыт на той же собаке на другой день.
 В первом опыте при меньшей исходной величине условный рефлекс, испробованный через минуты, уменьшился в два раза (3 вместо 6 капель), во втором после двукратного применения тормозной комбинации еще через 542 минут промежутка он был в 4 раза меньше (2 вместо 8 капель).
Второй важный, относящийся к последовательному торможению, факт - это то, что при повторении тормозной комбинации в ряде опытов ее последовательное тормозящее действие становится все короче. Начинаясь с десятков минут, оно постепенно сходит на минуты и даже секунды.
О п ы т. Вертящийся предмет - условный пищевой раздражитель, тон - условный тормоз к нему (опыты Николаева 2 VI 1909).
В первом опыте при меньшей исходной величине условный рефлекс, испробованный через минуты, уменьшился в два раза (3 вместо 6 капель), во втором после двукратного применения тормозной комбинации еще через 542 минут промежутка он был в 4 раза меньше (2 вместо 8 капель).
Второй важный, относящийся к последовательному торможению, факт - это то, что при повторении тормозной комбинации в ряде опытов ее последовательное тормозящее действие становится все короче. Начинаясь с десятков минут, оно постепенно сходит на минуты и даже секунды.
О п ы т. Вертящийся предмет - условный пищевой раздражитель, тон - условный тормоз к нему (опыты Николаева 2 VI 1909).
 О пыт на той же собаке 10 I 1910.
О пыт на той же собаке 10 I 1910.
 Что касается нарочитого разрушения действия тормозной комбинации и условного тормоза, то явление это в целом довольно сложное и до сих пор нами не выясненное с фактической стороны во всех его подробностях. Я упомяну сейчас только о наиболее установленных отношениях. Более быстрым способом полного устранения тормозной функции комбинации является, естественно, прием, совершенно обратный процедуре ее образования, т. е. сопровождение тормозной комбинации безусловным раздражителем данного рефлекса.
О п ы т. Тон духового камертона - условный кислотный раздражитель, механическое раздражение кожи - условный тормоз к нему (опыты Кржышковского).
Что касается нарочитого разрушения действия тормозной комбинации и условного тормоза, то явление это в целом довольно сложное и до сих пор нами не выясненное с фактической стороны во всех его подробностях. Я упомяну сейчас только о наиболее установленных отношениях. Более быстрым способом полного устранения тормозной функции комбинации является, естественно, прием, совершенно обратный процедуре ее образования, т. е. сопровождение тормозной комбинации безусловным раздражителем данного рефлекса.
О п ы т. Тон духового камертона - условный кислотный раздражитель, механическое раздражение кожи - условный тормоз к нему (опыты Кржышковского).

 В высшей степени интересно, что точное чередование приведенной процедуры разрушения с применением условного раздражителя данной комбинации с его обычным подкреплением чрезвычайно замедляет ход разрушения. Факт этот впоследствии будет подробно обследован и анализирован.
Кроме этого, во всяком случае постепенного ослабления тормозной комбинации, имеется быстрое нарушение тормозной комбинации, сейчас же наступающее и так же быстро исчезающее. F. сли во время действия тормозной комбинации на животное упадут раздражители из группы так называемых нами внешних гаснущих тормозов средней интенсивности, то тормозная комбинация обнаруживает в большей или меньшей степени положительное действие, свойственное ее условному раздражителю, применяемому отдельно. Следовательно, гаснущий тормоз устранил тормозное действие комбинации, т. e. мы встречаемся и здесь с кем же, что изучили при угасании, т. е. с растормаживанием.
Вот опыты из работ Николаева, демонстрирующие эти отношения.
Вертящийся предмет - условный пищевой раздражитель, тон - условный к нему тормоз, удары метронома, механическое и температурное раздражение кожи - посторонние агенты.
В высшей степени интересно, что точное чередование приведенной процедуры разрушения с применением условного раздражителя данной комбинации с его обычным подкреплением чрезвычайно замедляет ход разрушения. Факт этот впоследствии будет подробно обследован и анализирован.
Кроме этого, во всяком случае постепенного ослабления тормозной комбинации, имеется быстрое нарушение тормозной комбинации, сейчас же наступающее и так же быстро исчезающее. F. сли во время действия тормозной комбинации на животное упадут раздражители из группы так называемых нами внешних гаснущих тормозов средней интенсивности, то тормозная комбинация обнаруживает в большей или меньшей степени положительное действие, свойственное ее условному раздражителю, применяемому отдельно. Следовательно, гаснущий тормоз устранил тормозное действие комбинации, т. e. мы встречаемся и здесь с кем же, что изучили при угасании, т. е. с растормаживанием.
Вот опыты из работ Николаева, демонстрирующие эти отношения.
Вертящийся предмет - условный пищевой раздражитель, тон - условный к нему тормоз, удары метронома, механическое и температурное раздражение кожи - посторонние агенты.

 Таким образом, при посторонних раздражителях: ударах метронома, механическом раздражении кожи и ее нагревании, на время их действия устранялась тормозная функция тормозной комбинации, так что комбинация приобретала первоначальное положительное действие в большей или меньшей мере.
Очень интересен на той же собаке следующий опыт. Ради применения нового растормаживателя - запаха - она была переведена в новую комнату с запаховым прибором. Этот прибор, помимо действия как нового предмета перед глазами животного, действовал на него еще постояннным дуновением, обвиванием и стуком электрического мотора, значит, целым комплексом новых посторонних раздражителей.
Этот опыт, поставленный на следующий день после предшествующего, - 23 1909.
Таким образом, при посторонних раздражителях: ударах метронома, механическом раздражении кожи и ее нагревании, на время их действия устранялась тормозная функция тормозной комбинации, так что комбинация приобретала первоначальное положительное действие в большей или меньшей мере.
Очень интересен на той же собаке следующий опыт. Ради применения нового растормаживателя - запаха - она была переведена в новую комнату с запаховым прибором. Этот прибор, помимо действия как нового предмета перед глазами животного, действовал на него еще постояннным дуновением, обвиванием и стуком электрического мотора, значит, целым комплексом новых посторонних раздражителей.
Этот опыт, поставленный на следующий день после предшествующего, - 23 1909.
 Новый комплекс посторонних раздражителей растормаживал тормозную комбинацию, но действие посторонних раздражителей на животное все падало и падало с течением времени, а через 1'/2 часа после начала опыта и совершенно исчезло, как и происходит это часто с посторонними раздражителями, действующими в качестве внешних тормозов. Надо сказать, что в данном случае мы имели дело с собакой, давно находившейся в лаборатории, подвергавшейся многочисленным влияниям, на которую всякие новые перемены почти не оказывали никакого действия или очень быстро исчезающее; она скоро делалась в отношении их совершенно индифферентной. Вот почему у нее в новой обстановке и началось прямо с растормаживания. То же видно и при раздражении запахом камфоры. Он прямо только с первого раза растормозил комбинацию, а со второго сделался уже индифферентным для животного.
Совсем иначе шел опыт у другой собаки Николаева, которая была новичком в лаборатории, а может быть, обладала другим типом нервной системы - была более легко тормозимой.
О пыт 15 II 1910. Вертящийся предмет - условный пищевой раздражитель, тон - условный тормоз к нему, метроном посторонний агент.
Новый комплекс посторонних раздражителей растормаживал тормозную комбинацию, но действие посторонних раздражителей на животное все падало и падало с течением времени, а через 1'/2 часа после начала опыта и совершенно исчезло, как и происходит это часто с посторонними раздражителями, действующими в качестве внешних тормозов. Надо сказать, что в данном случае мы имели дело с собакой, давно находившейся в лаборатории, подвергавшейся многочисленным влияниям, на которую всякие новые перемены почти не оказывали никакого действия или очень быстро исчезающее; она скоро делалась в отношении их совершенно индифферентной. Вот почему у нее в новой обстановке и началось прямо с растормаживания. То же видно и при раздражении запахом камфоры. Он прямо только с первого раза растормозил комбинацию, а со второго сделался уже индифферентным для животного.
Совсем иначе шел опыт у другой собаки Николаева, которая была новичком в лаборатории, а может быть, обладала другим типом нервной системы - была более легко тормозимой.
О пыт 15 II 1910. Вертящийся предмет - условный пищевой раздражитель, тон - условный тормоз к нему, метроном посторонний агент.
 Опыт с тем же результатом повторился на следующий день, а еще через день (17 II 1910) в начале опыта пускали один метроном в течение минуты, а дальше опыт протекал следующим образом:
Опыт с тем же результатом повторился на следующий день, а еще через день (17 II 1910) в начале опыта пускали один метроном в течение минуты, а дальше опыт протекал следующим образом:
 В опыте 15 II удары метронома как постороннего раздражителя при первом применении, повидимому, совершенно не растормозили тормозной комбинации, но дело в том, что тот же метроном, присоединенный к одному условному раздражителю, сам его совершенно затормозил. Следовательно, и нуль рефлекса при первой комбинации (вертящийся предмет - тон + удары метронома) был, так сказать, не нулем оставшегося нетронутым внутреннего торможения, а нулем внешнего торможения вследствие очень сильного действия метронома при первом его применении как возбудителя постороннего рефлекса. Когда же после нескольких повторений метронома в комбинации 16 П и в отдельности в начале опыта 17 II значение его как возбудителя постороннего рефлекса упало и вместе тормозящее действие на положительный условный раздражитель почти или совсем исчезло, его растормаживающее действие на тормозную комбинацию выступает совершенно отчетливо, как это видно в опыте 17 II.
Конечно, способ выражения при истолковании последних опытов может казаться произвольным и искусственным, но этого требует неизбежная на первых порах схематизация сложных явлений, когда их внутренний механизм остается еще недоступным для анализа. Примененной формой выражения должны констатироваться только фактическое положение дела и последовательность явлений.
Растормаживание тормозной комбинации получается и с раздражителями из группы простых, постоянных тормозов. Вот относящийся сюда опыт из работы Николаева.
Вертящийся предмет - условный пищевой раздражитель, тон - условный тормоз к нему.
В опыте 15 II удары метронома как постороннего раздражителя при первом применении, повидимому, совершенно не растормозили тормозной комбинации, но дело в том, что тот же метроном, присоединенный к одному условному раздражителю, сам его совершенно затормозил. Следовательно, и нуль рефлекса при первой комбинации (вертящийся предмет - тон + удары метронома) был, так сказать, не нулем оставшегося нетронутым внутреннего торможения, а нулем внешнего торможения вследствие очень сильного действия метронома при первом его применении как возбудителя постороннего рефлекса. Когда же после нескольких повторений метронома в комбинации 16 П и в отдельности в начале опыта 17 II значение его как возбудителя постороннего рефлекса упало и вместе тормозящее действие на положительный условный раздражитель почти или совсем исчезло, его растормаживающее действие на тормозную комбинацию выступает совершенно отчетливо, как это видно в опыте 17 II.
Конечно, способ выражения при истолковании последних опытов может казаться произвольным и искусственным, но этого требует неизбежная на первых порах схематизация сложных явлений, когда их внутренний механизм остается еще недоступным для анализа. Примененной формой выражения должны констатироваться только фактическое положение дела и последовательность явлений.
Растормаживание тормозной комбинации получается и с раздражителями из группы простых, постоянных тормозов. Вот относящийся сюда опыт из работы Николаева.
Вертящийся предмет - условный пищевой раздражитель, тон - условный тормоз к нему.
 Латентное последействие соды, присоединившись к тормозной комбинации, растормозило ее.
Когда к тормозной комбинации присоединяется латентное последействие очень сильного агента, например сильного раствора хинина, то сначала наблюдается отсутствие растормаживания в силу тех же отношений, с которыми мы встретились в предшествующем опыте при применении метронома как растормаживателя. Но и здесь, при хинине, стоит только испробовать тормозную комбинацию на большем расстоянии от момента вливания его в рот, когда латентноего последействие несколько ослабнет, - растормаживающее его действие обнаруживается отчетливо, как и в случае с метрономом.
Все переданное относительно условного торможения делает совершенно очевидным, что процесс, происходящий при условном торможении, совершенно тождествен с процессом, который наступает при угасании. Основное условие их возникновения одно и то же. Это - отсутствие соучастия безусловного раздражителя. А затем и там и здесь процесс развивается постепенно, повторением усиливается, последующее тормозящее действие не ограничивается положительным условным раздражителем, который подвергается процедуре угашения или условного торможения, а простирается и на другие условные раздражители, даже и на разнородные, и, наконец, быстро и временно нарушается под влиянием посторонних раздражителей. Разница только в том, что при угасании положительный условный раздражитель один теряет свое действие, а при условном торможении - в соединении с посторонним агентом, являясь теперь, так сказать, как бы другим.
Латентное последействие соды, присоединившись к тормозной комбинации, растормозило ее.
Когда к тормозной комбинации присоединяется латентное последействие очень сильного агента, например сильного раствора хинина, то сначала наблюдается отсутствие растормаживания в силу тех же отношений, с которыми мы встретились в предшествующем опыте при применении метронома как растормаживателя. Но и здесь, при хинине, стоит только испробовать тормозную комбинацию на большем расстоянии от момента вливания его в рот, когда латентноего последействие несколько ослабнет, - растормаживающее его действие обнаруживается отчетливо, как и в случае с метрономом.
Все переданное относительно условного торможения делает совершенно очевидным, что процесс, происходящий при условном торможении, совершенно тождествен с процессом, который наступает при угасании. Основное условие их возникновения одно и то же. Это - отсутствие соучастия безусловного раздражителя. А затем и там и здесь процесс развивается постепенно, повторением усиливается, последующее тормозящее действие не ограничивается положительным условным раздражителем, который подвергается процедуре угашения или условного торможения, а простирается и на другие условные раздражители, даже и на разнородные, и, наконец, быстро и временно нарушается под влиянием посторонних раздражителей. Разница только в том, что при угасании положительный условный раздражитель один теряет свое действие, а при условном торможении - в соединении с посторонним агентом, являясь теперь, так сказать, как бы другим.
Лекция шестая. 2) внутреннее торможение: в) запаздывание
Мм. гг.! Перехожу к нашему третьему случаю внутреннего торможения, называемому у нас запаздыванием. Понятно, что комбинирование условного и безусловного раздражителей при выработке условного рефлекса можно очень разнообразить в отношении промежутка между началом условного раздражителя и моментом присоединения к нему безусловного. Промежутки эти могут быть короткие, 1-- секунд, может быть, даже и части секунды (но должно, как это уже было указано, непременно начинать с условного раздражителя), а также и более длинные, до нескольких минут. Таким образом происходят разные условные рефлексы, которыеМы отличаем разными названиями: то совпадающие, точнее говоря - почти совпадающие, то отставленные, или запаздывающие. Промежутки играют очень важную роль, так как ими, во-первых, обусловливается то торможение, которым мы теперь будем заниматься; во-вторых, они же определяют окончательную судьбу каждого условного рефлекса, как увидим это позже. Если промежуток постоянно берется малый, 1-5 секунд, то при выработке условного рефлекса и слюноотделение тоже начинается быстро, сейчас же после начала условного раздражителя. При увеличении промежутка слюноотделение все более и более запаздывает, отодвигается от начала условного раздражителя. Это запаздывание соответственно доходит до минут. Запаздывания можно достигать разными приемами. Один раз мы начинаем опыт с почти совпадающего рефлекса. Условная реакция наступает быстро - через 1--3 секунды. Затем мы постепенно увеличиваем промежуток, положим, каждый день на 5 секунд. Соответственно и условный эффект постепенно запаздывает, пока мы не остановимся на определенном промежутке и, таким образом, на определенном запаздывании. В другой раз мы сразу переходим с почти совпадающего рефлекса на очень отставленный. Тогда выработанная реакция сначала совершенно исчезает и на большой срок происходит длинный период нулей, как выражались некоторые мои сотрудники в своих работах. А кончается тем, что поблизости момента присоединения безусловного раздражителя появляется слюноотделение, которое с повторением так расположенного опыта все растет, подаваясь постепенно назад, т. е. к началу условного раздражителя, и останавливаясь, наконец, на значительном от него расстоянии. Начинать прямо с очень значительного отставления едва ли возможно, по крайней мере у огромного большинства собак, потому что собаки в таких случаях скоро погружаются в сонливое состояние, и тогда получить у них условный рефлекс не удается. Из-за указанной трудности этот последний прием не был у нас достаточно исследован. Вот образец запаздывающего рефлекса из работы Завадского, которому принадлежит заслуга особенно тщательного обследования данного случая внутреннего торможения. У собаки свисток - условный кислотный раздражитель. Начало условного раздражителя от начала безусловного отделено промежутком в 3 минуты. Скорость развития запаздывания чрезвычайно разнообразна. Прежде всего она находится в очень резкой зависимости от индивидуальности животного, характера его нервной системы. У одних собак запаздывание происходит очень быстро, у других, наоборот, очень медленно: слюноотделение упорно не отходит от начала условного раздражителя. У одних оно обозначается в течение одного опыта при повторении нескольких более отставленных рефлексов, у других для этого мало месяца и более. У первых запаздывание часто довольно скоро переходит в сонное состояние, и поэтому у некоторых из этих собак приходится при систематической работе над ними ограничиваться коротко отставленными рефлексами, т. e. довольствоваться меньшей величиной слюноотделения, чем у других.
Затем отчетливо дает себя знать род условных раздражителей. Механические и термические раздражения кожи и световые раздражения скорее ведут к развитию запаздывания, чем звуковые. При прочих равных условиях это идет рядом вообще с меньшим условным эффектом первых раздражений сравнительно со вторыми. Вот опыты из работы В. В. Яковлевой, иллюстрирующие эти отношения.
У собаки три условных пищевых раздражителя: удары метронома, механическое раздражение кожи и вспыхивание лампочки перед глазами.
О п ы т 9 IV 1924. Безусловный раздражитель отставлен от условного на 30 секунд.
Скорость развития запаздывания чрезвычайно разнообразна. Прежде всего она находится в очень резкой зависимости от индивидуальности животного, характера его нервной системы. У одних собак запаздывание происходит очень быстро, у других, наоборот, очень медленно: слюноотделение упорно не отходит от начала условного раздражителя. У одних оно обозначается в течение одного опыта при повторении нескольких более отставленных рефлексов, у других для этого мало месяца и более. У первых запаздывание часто довольно скоро переходит в сонное состояние, и поэтому у некоторых из этих собак приходится при систематической работе над ними ограничиваться коротко отставленными рефлексами, т. e. довольствоваться меньшей величиной слюноотделения, чем у других.
Затем отчетливо дает себя знать род условных раздражителей. Механические и термические раздражения кожи и световые раздражения скорее ведут к развитию запаздывания, чем звуковые. При прочих равных условиях это идет рядом вообще с меньшим условным эффектом первых раздражений сравнительно со вторыми. Вот опыты из работы В. В. Яковлевой, иллюстрирующие эти отношения.
У собаки три условных пищевых раздражителя: удары метронома, механическое раздражение кожи и вспыхивание лампочки перед глазами.
О п ы т 9 IV 1924. Безусловный раздражитель отставлен от условного на 30 секунд.
 Опыт 24 IV 1925. Рефлексы, многократно применявшиеся для других целей за годовой промежуток между этим и предшествующим опытом, затем были сделаны более отставленными: сперва на минуту, а потом на две. Всефлексы повторялись отставленными одинаковое число раз.
Опыт 24 IV 1925. Рефлексы, многократно применявшиеся для других целей за годовой промежуток между этим и предшествующим опытом, затем были сделаны более отставленными: сперва на минуту, а потом на две. Всефлексы повторялись отставленными одинаковое число раз.
 Большое влияние имеет длительная практика короткого отставления, которое иногда упорно не допускает развития более значительного запаздывания при большем отставлении без словного рефлекса от начала условного.
Наконец, повидимому, при раздражениях одного рода, например звуковых, запаздывание развивается скорее или медленнее в связи с тем, является ли раздражение сплошным или прерывистым. В случае сплошного оно наступает скорее.
Итак, при большом отставлении во времени безусловного раздражителя от начала условного эффект условного состоит из двух фаз, часто одинаковых по продолжительности: начальной недеятельной и второй деятельной. Что значит недеятельная фаза? Нарастает ли, постепенно суммируется [ли] раздражительный процесс, чтобы через очень большой период проявиться в видимом эффекте, или он, достаточный во силе, существует гораздо раньше, но до поры до времени благодаря чему-то маскируется, заслоняется? Уже обычные приемы при получении запаздывающих рефлексов говорят против первого предположения. При первоначальном коротком отставьении мы уже имеем вполне проявившийся эффект условного раздражения. Почему же тот же раздражитель при большей его продолжительности требовал бы большего суммационного времени? Если бы это было утомлением, то оно должно было бы вести к постепенному уменьшению эффекта. А этого нет. Эффект, хотя и запаздывая, при первом приеме выработки запаздывания (прием приведен в начале лекции) все увеличивается, а при втором - сначала исчезает, потом появляется вновь и, долго возрастая, наконец, становится постоянным на максимуме. Ввиду этого остается второе предположение, что здесь имеет место замаскирование раздражения, его временное задерживание. И это доказывается фактом, что раздражительный процесс в первой фазе можно быстро выявить.
Если мы в недеятельной фазе подействуем на наше животное каким-нибудь посторонним агентом, не имеющим до сих пор никакого отношения к работе слюнных желез, то мы получим сейчас же слюноотделение, и часто обильное, вместе с соответствуюшей данному условному раздражителю двигательной реакцией, т. е. условный рефлекс выступает целиком.
Вот опыты Завадского.
Механическое раздражение кожи условный кислотный раздражитель. Начало безусловного раздражителя от начала продолжающегося все время условного раздражителя отделено промежутком в минуты. Удары метронома никакого отношения к кислотному рефлюксу доселе не имели.
Большое влияние имеет длительная практика короткого отставления, которое иногда упорно не допускает развития более значительного запаздывания при большем отставлении без словного рефлекса от начала условного.
Наконец, повидимому, при раздражениях одного рода, например звуковых, запаздывание развивается скорее или медленнее в связи с тем, является ли раздражение сплошным или прерывистым. В случае сплошного оно наступает скорее.
Итак, при большом отставлении во времени безусловного раздражителя от начала условного эффект условного состоит из двух фаз, часто одинаковых по продолжительности: начальной недеятельной и второй деятельной. Что значит недеятельная фаза? Нарастает ли, постепенно суммируется [ли] раздражительный процесс, чтобы через очень большой период проявиться в видимом эффекте, или он, достаточный во силе, существует гораздо раньше, но до поры до времени благодаря чему-то маскируется, заслоняется? Уже обычные приемы при получении запаздывающих рефлексов говорят против первого предположения. При первоначальном коротком отставьении мы уже имеем вполне проявившийся эффект условного раздражения. Почему же тот же раздражитель при большей его продолжительности требовал бы большего суммационного времени? Если бы это было утомлением, то оно должно было бы вести к постепенному уменьшению эффекта. А этого нет. Эффект, хотя и запаздывая, при первом приеме выработки запаздывания (прием приведен в начале лекции) все увеличивается, а при втором - сначала исчезает, потом появляется вновь и, долго возрастая, наконец, становится постоянным на максимуме. Ввиду этого остается второе предположение, что здесь имеет место замаскирование раздражения, его временное задерживание. И это доказывается фактом, что раздражительный процесс в первой фазе можно быстро выявить.
Если мы в недеятельной фазе подействуем на наше животное каким-нибудь посторонним агентом, не имеющим до сих пор никакого отношения к работе слюнных желез, то мы получим сейчас же слюноотделение, и часто обильное, вместе с соответствуюшей данному условному раздражителю двигательной реакцией, т. е. условный рефлекс выступает целиком.
Вот опыты Завадского.
Механическое раздражение кожи условный кислотный раздражитель. Начало безусловного раздражителя от начала продолжающегося все время условного раздражителя отделено промежутком в минуты. Удары метронома никакого отношения к кислотному рефлюксу доселе не имели.
 Происходит очень интересное и неожиданное явление: условный выработанный раздражитель один не действует 1-11/2 минуты, а индифферентный агент, впервые к нему в это время прибавленный, тотчас же обусловливает заканомерную реакцию данного условного раздражителя.
Очевидно, что мы здесь имеем дело с тем же явлением, с которым познакомились раньше и которое было названо растормаживанием. Так как в нашей лаборатории впервые оно наиболее систематически было исследовано Завадоким, то я и остановлюсь более подробно на его опытах.
Как вы могли заметить, я в трех лекциях подряд говорю о растормаживании и привожу относительно его много протокольного материала. Оправданием для этого, как мне кажется, должна служить относительная физиологическая своеобразность явления, хотя и для него, конечно, имеются аналогии в теперешней нервной физиологии, и затем полная темнота, что касается понимания его внутреннего механизма. Можно надеяться, что изобилие фактических подробностей может когда-нибудь пособить в понимании этого механизма.
Обращаю ваше внимание на первый из только что приведенных двух опытов. Мы видим в нем отчетливо, что присоединенные к механическому раздражению кожи удары метронома не только вызвали слюноотделение в недеятельной фазе, но вместе с тем резко уменьшили слюноотделительный эффект во второй, деятельной фазе. Во всех случаях одного механического раздражения кожи за 1/2 минуты выделилось слюны от 29 до 46 капель; при раздражении кожи совместно с ударами метронома 17 капель. Таким образом мы наблюдаем двойное действие постороннего агента, растормаживающее в недеятельной фазе и тормозящее в деятельной.
Если присоединять к условному раздражителю в запаздывающем рефлексе различные посторонние агенты из группы внешних тормозов, то получается очень разнообразная картина изменений в течение запаздывающего рефлекса, в которой, однако, легко открывается определенная закономерность. Все посторонние агенты, применяемые в этих наших опытах, могут быть расположены в следующий ряд по их влиянию на запаздывающие рефлексы.
I. Температура 5° С и 44° С, приложенная к коже, и слабый запах камфоры.
II. Температура 0.5° С и 50° С.
III. Бесшумно вертящийся перед глазами собаки предмет, удары метронома, механическое раздражение необычных мест (если механическое раздражение одного места кожи есть условное раздражение), свисток умеренной силы, запах уксусного амила.
IV. Интенсивный запах камфоры, сильные свистки и звонки.
Первый ряд раздражителей не оказывает никакого влияния на ход запаздывающего рефлекса.
Второй ряд обнаруживает свое действие только на первой фазе рефлекса, вызывая во время ее отделение слюны.
Третий ряд нарушает обе фазы, в первой вызывая слюноотделение, во второй уменьшая его сравнительно с нормой.
Наконец четвертый ряд почти или совсем не изменяет первую фазу, но вполне или почти подавляет слюноотделение во второй.
Нужно прибавить, что эти опыты при соблюдении определенных предосторожностей идут часто с поражающей точностью.
Вот ряд опытов, иллюстрирующих указанные отношения.
Механическое раздражение кожи - условный кислотный раздражитель. Начало условного раздражителя от начала безусловного отделено промежутком в 3 минуты. Посторонними раздражителями служат температура в 44 С, приложенная к коже, температура в 0.5° С, запах уксусного амила и звонок.
Происходит очень интересное и неожиданное явление: условный выработанный раздражитель один не действует 1-11/2 минуты, а индифферентный агент, впервые к нему в это время прибавленный, тотчас же обусловливает заканомерную реакцию данного условного раздражителя.
Очевидно, что мы здесь имеем дело с тем же явлением, с которым познакомились раньше и которое было названо растормаживанием. Так как в нашей лаборатории впервые оно наиболее систематически было исследовано Завадоким, то я и остановлюсь более подробно на его опытах.
Как вы могли заметить, я в трех лекциях подряд говорю о растормаживании и привожу относительно его много протокольного материала. Оправданием для этого, как мне кажется, должна служить относительная физиологическая своеобразность явления, хотя и для него, конечно, имеются аналогии в теперешней нервной физиологии, и затем полная темнота, что касается понимания его внутреннего механизма. Можно надеяться, что изобилие фактических подробностей может когда-нибудь пособить в понимании этого механизма.
Обращаю ваше внимание на первый из только что приведенных двух опытов. Мы видим в нем отчетливо, что присоединенные к механическому раздражению кожи удары метронома не только вызвали слюноотделение в недеятельной фазе, но вместе с тем резко уменьшили слюноотделительный эффект во второй, деятельной фазе. Во всех случаях одного механического раздражения кожи за 1/2 минуты выделилось слюны от 29 до 46 капель; при раздражении кожи совместно с ударами метронома 17 капель. Таким образом мы наблюдаем двойное действие постороннего агента, растормаживающее в недеятельной фазе и тормозящее в деятельной.
Если присоединять к условному раздражителю в запаздывающем рефлексе различные посторонние агенты из группы внешних тормозов, то получается очень разнообразная картина изменений в течение запаздывающего рефлекса, в которой, однако, легко открывается определенная закономерность. Все посторонние агенты, применяемые в этих наших опытах, могут быть расположены в следующий ряд по их влиянию на запаздывающие рефлексы.
I. Температура 5° С и 44° С, приложенная к коже, и слабый запах камфоры.
II. Температура 0.5° С и 50° С.
III. Бесшумно вертящийся перед глазами собаки предмет, удары метронома, механическое раздражение необычных мест (если механическое раздражение одного места кожи есть условное раздражение), свисток умеренной силы, запах уксусного амила.
IV. Интенсивный запах камфоры, сильные свистки и звонки.
Первый ряд раздражителей не оказывает никакого влияния на ход запаздывающего рефлекса.
Второй ряд обнаруживает свое действие только на первой фазе рефлекса, вызывая во время ее отделение слюны.
Третий ряд нарушает обе фазы, в первой вызывая слюноотделение, во второй уменьшая его сравнительно с нормой.
Наконец четвертый ряд почти или совсем не изменяет первую фазу, но вполне или почти подавляет слюноотделение во второй.
Нужно прибавить, что эти опыты при соблюдении определенных предосторожностей идут часто с поражающей точностью.
Вот ряд опытов, иллюстрирующих указанные отношения.
Механическое раздражение кожи - условный кислотный раздражитель. Начало условного раздражителя от начала безусловного отделено промежутком в 3 минуты. Посторонними раздражителями служат температура в 44 С, приложенная к коже, температура в 0.5° С, запах уксусного амила и звонок.

 Совершенно те же в существенном результаты получаются, если посторонние раздражители присоединяются к условному раздражителю не на всем его протяжении от его начала до присоединения безусловного раздражителя, а отдельно то в недеятельную фазу (первые 17/2 минуты), то в деятельную (вторые 1'/2 минуты).
Вот примеры.
То же животное и те же отношения
Совершенно те же в существенном результаты получаются, если посторонние раздражители присоединяются к условному раздражителю не на всем его протяжении от его начала до присоединения безусловного раздражителя, а отдельно то в недеятельную фазу (первые 17/2 минуты), то в деятельную (вторые 1'/2 минуты).
Вот примеры.
То же животное и те же отношения
 Посторонний раздражитель из третьей группы отдельно вызвал слюноотделение в первой фазе и уменьшил его во второй; в первом опыте некоторое уменьшение его во второй фазе, конечно, произошло от следов раздражения.
Теперь поднимается вопрос: что это за серия посторонних раздражителей? Что лежит в ее основании? По многим данным становится ясным, что здесь идет дело об относительной физиологической силе посторонних раздражителей, о величине реакций на них животного. Приведенный ряд есть ряд все более и более увеличивающихся по силе действия раздражителей. В некоторых случаях это прямо видно в таблице, когда применяются разные интенсивности одних и тех же агентов и когда, стало быть, соответственно изменяются и реакции на них животного. А затем и во всех случаях это обнаруживалось вигательной реакции животного. При первых членах ряда часто не замечалось ни малейшей реакции. При дальнейших членах ряда реакция становилась постепенно все энергичнее и продолжительнее. Наконец применяемые нами теперь посторонние раздражители суть агенты так называемого нами внешнего торможения, и на деятельной фазе анализируемых рефлексов вполне отчетливо и тем резче сказывается их тормозящее действие, чем более вниз мы подвигаемся по приведенной таблице, кончая полным торможением. Следовательно, наш ряд посторонних раздражителей есть действительно ряд по нарастающему их действию на организм, начиная с легкого исследовательского рефлекса.
Что это понимание фактов правильно, вполне подтверждается и следующей вариацией опытов. Если посторонний раздражитель повторно присоединять к запаздывающему рефлексу, то его гормозящее действие на деятельную фазу постепенно раз от разу падает. А это есть совершенно понятное свойство посторонних раздражителей как гаснущих тормозов.
Bor Вот опыты с тем же животным, что выше.
Посторонний раздражитель из третьей группы отдельно вызвал слюноотделение в первой фазе и уменьшил его во второй; в первом опыте некоторое уменьшение его во второй фазе, конечно, произошло от следов раздражения.
Теперь поднимается вопрос: что это за серия посторонних раздражителей? Что лежит в ее основании? По многим данным становится ясным, что здесь идет дело об относительной физиологической силе посторонних раздражителей, о величине реакций на них животного. Приведенный ряд есть ряд все более и более увеличивающихся по силе действия раздражителей. В некоторых случаях это прямо видно в таблице, когда применяются разные интенсивности одних и тех же агентов и когда, стало быть, соответственно изменяются и реакции на них животного. А затем и во всех случаях это обнаруживалось вигательной реакции животного. При первых членах ряда часто не замечалось ни малейшей реакции. При дальнейших членах ряда реакция становилась постепенно все энергичнее и продолжительнее. Наконец применяемые нами теперь посторонние раздражители суть агенты так называемого нами внешнего торможения, и на деятельной фазе анализируемых рефлексов вполне отчетливо и тем резче сказывается их тормозящее действие, чем более вниз мы подвигаемся по приведенной таблице, кончая полным торможением. Следовательно, наш ряд посторонних раздражителей есть действительно ряд по нарастающему их действию на организм, начиная с легкого исследовательского рефлекса.
Что это понимание фактов правильно, вполне подтверждается и следующей вариацией опытов. Если посторонний раздражитель повторно присоединять к запаздывающему рефлексу, то его гормозящее действие на деятельную фазу постепенно раз от разу падает. А это есть совершенно понятное свойство посторонних раздражителей как гаснущих тормозов.
Bor Вот опыты с тем же животным, что выше.
 В этих опытах мы видим, как тормозящее действие постороннего раздражителя на деятельную фазу с повторением постепенно падает, особенно правильно во втором опыте.
Отношения, выступившие во всех приведенных опытах, проделанных на одном и том же животном, повторились и на других животных. Разница оказалась только в некотором изменение состава рядов посторонних раздражителей, члены которых располагались отчасти в другом порядке, что и понятно. Реакции животных на разные посторонние раздражители очень, даже чрезвычайно часто, колеблются в интенсивности в зависимости от индивидуального характера нервной системы и от прошлого животного, от его встреч в жизни с темиили другими раздражителями, как это каждый из нас знает хорошо по себе.
Таким образом мы должны признать, что указанный выше порядок изменений эффекта запаздывающего рефлекса под влиянием посторонних раздражителей действительно определяется различной физиологической силой этих раздражителей. При малой силе их незаметно никакого изменения на нашем рефлексе. При некотором увеличении ее терпит изменение только недеятельная фаза, превращаясь в деятельную. При еще более значительном увеличении одновременно изменяется и деятельная фаза, очень теряя в своем размере, часто сравниваясь, а иногда даже и уступая первой фазе, ставшей теперь деятельной. Наконец при высшей силе некоторых посторонних раздражителей запаздывающий рефлекс делается нулевым на всем своем протяжении.
В описанных изменениях запаздывающего рефлекса под влиянием посторонних раздражителей особенно резко подчеркиваются два важных факта, относящихся деятельности центральной нервной системы. Первый, что посторонний раздражитель, действуя на положительный условный раздражитель, его тормозит, а падая на отрицательный, тормозной, его растормаживает; иначе говоря, в обоих случаях изменяет наличный центральный нервный процесс в обратный. А второй, что тормозной процесс сравнительно с раздражительным процессом лабильнее, так как более легко, под влиянием более слабых посторонних раздражителей, испытывает изменение.
Относительно этого второго пункта у меня имеется следующий случайный, но поучительный опыт. Я читал публичные лекции об условных рефлексах в новом отдаленном месте (собак пришлось перевозить) и при вообще необытной для наших животных обстановке. Все опыты с положительными условными рефлексами в числе 5-6, демонстрированных на первой лекции, протекли вполне хорошо. Все опыты на тех же животных с торразными условными рефлексами (тоже в числе 5-6) на следующей лекции были неудачны, т. е. во всех них наступило растормаживание. Таким образом одна и та же обстановка нисколько не нарушила положительных условных рефлексов, но, даже повторенная, резко повлияла на тормозные условные рефлексы.
К обсуждению этих фактов мы вернемся в более поздней лекции в связи с многочисленными фактами, относящимися к тому же пункту взаимоотношения между раздражительным и тормозным процессами.
Так как в первой фазе запаздывающего рефлекса мы имеем совершенно тот же тормозной процесс, что при угасании и условном торможении, то и в подробностях относительно него мы должны ожидать аналогичных отношений.
При угасании и условном торможении мы видели, что торможение, достигнутое для определенного условного раздражителя, простирается само собой и на другие условные раздражители, но в мере, которая определяется относительной физиологической силой этих раздражителей. Если другие условные раздражители слабее того, на котором было развито или выработано торможение, то вторичное последовательное торможение этих других раздражителей полное; если же они сильнее, то торможение только частичное, т. е. вырабатывающееся торможение как бы точно отмеривается по размеру того условного раздражителя, на котором оно вырабатывалось. То же самое наблюдается и на тормозном процессе запаздывающего рефлекса. С изменением силы условного раздражителя этого рефлекса установившееся отношение между недеятельной и деятельной фазами резко нарушается то в одну, то в другую сторону, соответственно изменению силы раздражителя.
Вот относящиеся сюда опыты на том же животном.
В этих опытах мы видим, как тормозящее действие постороннего раздражителя на деятельную фазу с повторением постепенно падает, особенно правильно во втором опыте.
Отношения, выступившие во всех приведенных опытах, проделанных на одном и том же животном, повторились и на других животных. Разница оказалась только в некотором изменение состава рядов посторонних раздражителей, члены которых располагались отчасти в другом порядке, что и понятно. Реакции животных на разные посторонние раздражители очень, даже чрезвычайно часто, колеблются в интенсивности в зависимости от индивидуального характера нервной системы и от прошлого животного, от его встреч в жизни с темиили другими раздражителями, как это каждый из нас знает хорошо по себе.
Таким образом мы должны признать, что указанный выше порядок изменений эффекта запаздывающего рефлекса под влиянием посторонних раздражителей действительно определяется различной физиологической силой этих раздражителей. При малой силе их незаметно никакого изменения на нашем рефлексе. При некотором увеличении ее терпит изменение только недеятельная фаза, превращаясь в деятельную. При еще более значительном увеличении одновременно изменяется и деятельная фаза, очень теряя в своем размере, часто сравниваясь, а иногда даже и уступая первой фазе, ставшей теперь деятельной. Наконец при высшей силе некоторых посторонних раздражителей запаздывающий рефлекс делается нулевым на всем своем протяжении.
В описанных изменениях запаздывающего рефлекса под влиянием посторонних раздражителей особенно резко подчеркиваются два важных факта, относящихся деятельности центральной нервной системы. Первый, что посторонний раздражитель, действуя на положительный условный раздражитель, его тормозит, а падая на отрицательный, тормозной, его растормаживает; иначе говоря, в обоих случаях изменяет наличный центральный нервный процесс в обратный. А второй, что тормозной процесс сравнительно с раздражительным процессом лабильнее, так как более легко, под влиянием более слабых посторонних раздражителей, испытывает изменение.
Относительно этого второго пункта у меня имеется следующий случайный, но поучительный опыт. Я читал публичные лекции об условных рефлексах в новом отдаленном месте (собак пришлось перевозить) и при вообще необытной для наших животных обстановке. Все опыты с положительными условными рефлексами в числе 5-6, демонстрированных на первой лекции, протекли вполне хорошо. Все опыты на тех же животных с торразными условными рефлексами (тоже в числе 5-6) на следующей лекции были неудачны, т. е. во всех них наступило растормаживание. Таким образом одна и та же обстановка нисколько не нарушила положительных условных рефлексов, но, даже повторенная, резко повлияла на тормозные условные рефлексы.
К обсуждению этих фактов мы вернемся в более поздней лекции в связи с многочисленными фактами, относящимися к тому же пункту взаимоотношения между раздражительным и тормозным процессами.
Так как в первой фазе запаздывающего рефлекса мы имеем совершенно тот же тормозной процесс, что при угасании и условном торможении, то и в подробностях относительно него мы должны ожидать аналогичных отношений.
При угасании и условном торможении мы видели, что торможение, достигнутое для определенного условного раздражителя, простирается само собой и на другие условные раздражители, но в мере, которая определяется относительной физиологической силой этих раздражителей. Если другие условные раздражители слабее того, на котором было развито или выработано торможение, то вторичное последовательное торможение этих других раздражителей полное; если же они сильнее, то торможение только частичное, т. е. вырабатывающееся торможение как бы точно отмеривается по размеру того условного раздражителя, на котором оно вырабатывалось. То же самое наблюдается и на тормозном процессе запаздывающего рефлекса. С изменением силы условного раздражителя этого рефлекса установившееся отношение между недеятельной и деятельной фазами резко нарушается то в одну, то в другую сторону, соответственно изменению силы раздражителя.
Вот относящиеся сюда опыты на том же животном.

 В опытах, если не отчетливо увеличение деятельной фазы с усилением условного раздражителя, то резко уменьшение се с ослаблением раздражителя.
To Точно таким же образом на запаздывании сказывается и сила безусловного раздражителя. Это удобно демонстрируется на условном пищевом раздражителе. Стоит только сравнить распределение фаз запаздывающего рефлекса на обычно питаемом животном и подвергнутом некоторому голоданию.
О пы т. Другое животное. Свисток средней силы - условный пищевой раздражитель. Отставление -- 5инуты.
О пыт 13 XiI 1907 при обычном питании.
В опытах, если не отчетливо увеличение деятельной фазы с усилением условного раздражителя, то резко уменьшение се с ослаблением раздражителя.
To Точно таким же образом на запаздывании сказывается и сила безусловного раздражителя. Это удобно демонстрируется на условном пищевом раздражителе. Стоит только сравнить распределение фаз запаздывающего рефлекса на обычно питаемом животном и подвергнутом некоторому голоданию.
О пы т. Другое животное. Свисток средней силы - условный пищевой раздражитель. Отставление -- 5инуты.
О пыт 13 XiI 1907 при обычном питании.
 При голодном состоянии недеятельная фаза, тормозной процесс почти исчезли.
При запаздывании наблюдается такая же суммация торможения, какую мы видели при угасании и при условном торможении. Это явственно выступает на следующем опыте.
Механическое раздражение кожи - условный раздражитель. Отставление - 3 минуты.
При голодном состоянии недеятельная фаза, тормозной процесс почти исчезли.
При запаздывании наблюдается такая же суммация торможения, какую мы видели при угасании и при условном торможении. Это явственно выступает на следующем опыте.
Механическое раздражение кожи - условный раздражитель. Отставление - 3 минуты.
 Механическое раздражение кожи в первые 1/2 минуты, обусловливающее торможение, повторенное несколько раз без продолжения его во вторые 17/2 минуты, повело к такому усилению тормозного процесса, что после этого примененное раздражение кожи во вторые минуты в первом случае сильно потеряло в своем положительном эффекте, а при повторении этой процедуры еще раз и совсем лишилось своего действия.
Как понимать, что в запаздывающем рефлексе один и тот же раздражитель сначала действует тормозящим образом, а затем возбуждающим? Что обусловливает противоположное действие одного и того же раздражителя, повидимому, при совершенно одинаковых условиях? Можно ли привести это явление в согласие с уже нам известными фактами? Я думаю, что да. В третьей и пятой лекциях мы познакомились с агентами, из которых могут быть образованы как положительные, так и тормозные узловные раздражители. Между ними упоминалось время как реальный агент. Там было разъяснено, как его надо понимать физиологически, и были приведены опыты, иллюстрирующие его роль как раздражителя. Особенно прошу вспомнить вариацию опыта, когда был применен суммарный раздражитель, состоящий из внешнего агента и из определенного момента времени. Тогда один внешний агент без надлежащего момента времени не действовал, а его действие появлялось и постоянно возрастало лишь по мере того, как приближался надлежащий момент времени. В запаздывающем рефлексе совершенно то же самое. В приведенных опытах безусловный раздражитель присоединялся к внешнему агенту только по истечении третьей минуты, т. е. только конец третьей минуты вместе с внешним агентом составляли тот настоящий суммарный агент, за которым непосредственно следовал безусловный раздражитель, и он-то (этот суммарный агент) специально и должен был сделаться условным раздражителем. Внешний же агент в соединении с другими моментами времени, отдаленными от конца третьей минуты, был другим суммарным агентом, который систематически не сопровождался безусловным рефлексом, и на него должен был вырабатываться тормозной рефлекс, как на всякий несколько отличающийся агент, что мы уже и видели при случае условного торможения и с чем мы в еще более резких примерах познакомимся в следующей лекции на случае так называемого нами дифференцировочного торможения. В запаздывающем рефлексе значение момента времени как раздражителя может быть представлено еще конкретнее и проще. Мы продолжаем избранный для образования условного рефлекса внешчий агент известное время, а в каждый момент его продолжения он оказывается для организма другим. Мы знаем, как быстро мы привыкаем к запахам, звукам и освещению, т. е. ощущаем их все иначе. А это, конечно, значит, что нервные клетки, на которые падают раздражения, переживают различное последовательные состояния. И только определенное состояние, совпадающее с безусловным рефлексом, становится условным раздражителем. В следующей же лекции мы увидим, до какой степени простирается дифференцирование интенсивностей одного и того же агента, причем одна интенсивность становится положительно, а другая отрицательно действующим раздражителем. Таким образом мы имеем несколько несомненных фактов, которые делают понятным факт запаздывающего рефлекса.
Факт развития запаздывания имеет очень большое значение ь методическом отношении, и при разнообразном исследовании условных рефлексов с ним необходимо постоянно считаться. Конечно, для того чтобы иметь данные о размере условного рефлекса и его многочисленных и тонких вариациях при разных условиях, мы должны отодвигать на больший или меньший период времени момент присоединения безусловного раздражителя от начального момента условного. Но это влечет за собой в большей или меньшей мере сейчас же развитие запаздывания, т. е. при этом рядом с раздражительным процессом возникает тормозной. Естественно, это осложняет исследование, потому что приходится иметь дело с двумя процессами, хотя бы вопрос касался одного раздражительного. Указанное обстоятельство, например, совершенно исключает возможности при обыкновенной форме наших опытов с условными рефлексами определения истинного латентного периода этих рефлексов. То, что мы обыкновенно при наших исследованиях называем латентным периодом, и то, что вы видели при нашем первом опыте (помните, там прошло 8 секунд между первым ударом метронома и появлением первой капли слюны), есть запаздывание, вмешательство тормозного периода и заслуживало бы точного термина «предварительный тормозной период». Поэтому для цели определения истинного латентного периода условных рефлексов необходимо пользоваться совпадающими рефлексами, когда безусловный раздражитель возможно скоро, через секунду или даже части секунды, следует за началом условного. И только при таких рефлексах, главным образом пользуясь двигательным компонентом наших рефлексов, можно убедиться, что величина латентного периода условных рефлексов действительно падает в категорию величин обыкновенного рефлекторного времени. Этими определениями у нас занимаются только теперь. Мы не придавали фундаментального значения определениютинного латентного периода условных рефлексов при установлении их рефлекторной натуры, считая главным доказательством этого их непременность и закономерность при известных условиях, так как и при обыкновенных рефлексах латентныериоды значительно колеблются, находясь в зависимости от тех или других центральных связей, или путей. Можно с правом допустить, не угрожая рефлекторной натуре условных рефлексов, что в больших полушариях эти связи сложнее.
Решение многих других вопросотносительно условных рефлексов, например вопрос о подлинном, беспрепятственном течении при них раздражительного процесса с самого его начала и т. п., также более или менее затрудняется вмешательством запаздывания. В этом отношении много помогает делу характер нервной системы разных животных. У иных из них, как это было указано выше, запаздывание развивается с трудом, и тогда раздражительный процесс или совсем не нарушается, или очень мало. Но и в руках экспериментатора есть некоторое средство ослаблять влияние запаздывания, ограничиваясь возможно коротким отставлением безусловного раздражителя от начала условного, если величина условного рефлекса вообще велика, т. е. если и за короткий срок она достаточна для сравнения при различных вариациях опыта. Но, с другой стороны, есть случаи, когда предварительный тормозной процесс представляет некоторую выгоду, дает возможность для решения некоторых вопросов, как с этим мы встретимся в следующей лекции. Таким образом приходится приспособляться, соответственно пользоваться имеющимися отношениями.
Познакомившись в последних трех лекциях с основным фактическим материалом относительно внутреннего торможения условных рефлексов, нетрудно усмотреть его огромную биологическую ценность. Этим торможением постоянно корригируется и совершенствуется сигнализационная деятельность больших полушарий. Если в данное время сигнальный, условный раздражитель повторно не сопровождается его, так сказать, безусловно-деловым раздражителем, он как убыточный для организма, напрасно вызывающий трату энергии, на некоторое, как правило короткое, время лишается своего физиологического действия. Если условно действующий агент, постоянно встречаясь с посторонним агентом, при условии этой встречи, не сопровождается безусловным раздражителем, он также теряет свое условное значение, но только в этой комбинации, являясь в таком случае как бы другим агентом. Наконец если условный агент всегда только в определенный момент после своего возникновения совпадает с действием безусловного раздражителя, то он при более или менее отдаленно предшествующих этому раздражителю моментах делается недействительным, проявляясь все в большей и большей степени по мере приближения к моменту присоединения безусловного раздражителя. Таким образом постоянно обнаруживается высшее приспособление организма к окружающим условиям, или, иначе говоря, более тонкое и точное уравновешивание организма с внешней средой.
Как ясно из всего сообщенного в последних трех лекциях, мы при угасании, условном торможении и запаздывании имеем перед собой образование условных тормозных рефлексов. Но эти же рефлексы могут быть получены еще и при совсем иной процедуре. Если с условными тормозными раздражителями одновременно на короткие сроки, но повторно, применять индифферентные агенты, то эти последние приобретают такое тормозную функцию, т. е. и сами по себе вызывают в коре процесс торможения. Этот предмет специально разработан Ю. В. Фольбортом. Прежде всего он делал посторонние агенты индифферентными, повторяя их перед животным до тех пор, пока они не переставали тормозить положительных условных рефлексов и растормаживать тормозных. Затем он присоединял их повторно на короткие сроки то к угашенным условным рефлексам, то к тормозным комбинациям. И после этого испытывал их действие, комбинируя их с положительными условными раздражителями, прияем и обнаруживал теперь их роль как условных тормозов. Так как эти испытания естественно приходилось ради точности результата повторять не один раз, то Фольборт один раз сопровождал пробную комбинацию безусловным рефлексом, в другой - нет, делая это строго поочередно, чтобы при этих повторных испытаниях комбинация не получила благодаря им постоянного, определенного, то положительного, то отрицательного характера. Но, конечно, бесспорнее и убедительнее была та форма опыта, когда он при испытаниях всегда сопровождал пробную комбинацию безусловным рефлексом, а торможение, несмотря на это, при повторении опыта, все же резко выступало.
Вот эти опыты.
На собаке применялся натуральный условный пищевой рефлекс. Когда он угашался до нуля, к нему повторно присоединялись сделанные раньше индифферентными удары метронома, а затем при продолжении этого приема изредка пробовалась комбинация ударов метронома с положительным пищевым рефлексом, едой.
О п ы т 5 IX 1911 после 10 совпадений ударов метронома с угашенным условным рефлексом.
Механическое раздражение кожи в первые 1/2 минуты, обусловливающее торможение, повторенное несколько раз без продолжения его во вторые 17/2 минуты, повело к такому усилению тормозного процесса, что после этого примененное раздражение кожи во вторые минуты в первом случае сильно потеряло в своем положительном эффекте, а при повторении этой процедуры еще раз и совсем лишилось своего действия.
Как понимать, что в запаздывающем рефлексе один и тот же раздражитель сначала действует тормозящим образом, а затем возбуждающим? Что обусловливает противоположное действие одного и того же раздражителя, повидимому, при совершенно одинаковых условиях? Можно ли привести это явление в согласие с уже нам известными фактами? Я думаю, что да. В третьей и пятой лекциях мы познакомились с агентами, из которых могут быть образованы как положительные, так и тормозные узловные раздражители. Между ними упоминалось время как реальный агент. Там было разъяснено, как его надо понимать физиологически, и были приведены опыты, иллюстрирующие его роль как раздражителя. Особенно прошу вспомнить вариацию опыта, когда был применен суммарный раздражитель, состоящий из внешнего агента и из определенного момента времени. Тогда один внешний агент без надлежащего момента времени не действовал, а его действие появлялось и постоянно возрастало лишь по мере того, как приближался надлежащий момент времени. В запаздывающем рефлексе совершенно то же самое. В приведенных опытах безусловный раздражитель присоединялся к внешнему агенту только по истечении третьей минуты, т. е. только конец третьей минуты вместе с внешним агентом составляли тот настоящий суммарный агент, за которым непосредственно следовал безусловный раздражитель, и он-то (этот суммарный агент) специально и должен был сделаться условным раздражителем. Внешний же агент в соединении с другими моментами времени, отдаленными от конца третьей минуты, был другим суммарным агентом, который систематически не сопровождался безусловным рефлексом, и на него должен был вырабатываться тормозной рефлекс, как на всякий несколько отличающийся агент, что мы уже и видели при случае условного торможения и с чем мы в еще более резких примерах познакомимся в следующей лекции на случае так называемого нами дифференцировочного торможения. В запаздывающем рефлексе значение момента времени как раздражителя может быть представлено еще конкретнее и проще. Мы продолжаем избранный для образования условного рефлекса внешчий агент известное время, а в каждый момент его продолжения он оказывается для организма другим. Мы знаем, как быстро мы привыкаем к запахам, звукам и освещению, т. е. ощущаем их все иначе. А это, конечно, значит, что нервные клетки, на которые падают раздражения, переживают различное последовательные состояния. И только определенное состояние, совпадающее с безусловным рефлексом, становится условным раздражителем. В следующей же лекции мы увидим, до какой степени простирается дифференцирование интенсивностей одного и того же агента, причем одна интенсивность становится положительно, а другая отрицательно действующим раздражителем. Таким образом мы имеем несколько несомненных фактов, которые делают понятным факт запаздывающего рефлекса.
Факт развития запаздывания имеет очень большое значение ь методическом отношении, и при разнообразном исследовании условных рефлексов с ним необходимо постоянно считаться. Конечно, для того чтобы иметь данные о размере условного рефлекса и его многочисленных и тонких вариациях при разных условиях, мы должны отодвигать на больший или меньший период времени момент присоединения безусловного раздражителя от начального момента условного. Но это влечет за собой в большей или меньшей мере сейчас же развитие запаздывания, т. е. при этом рядом с раздражительным процессом возникает тормозной. Естественно, это осложняет исследование, потому что приходится иметь дело с двумя процессами, хотя бы вопрос касался одного раздражительного. Указанное обстоятельство, например, совершенно исключает возможности при обыкновенной форме наших опытов с условными рефлексами определения истинного латентного периода этих рефлексов. То, что мы обыкновенно при наших исследованиях называем латентным периодом, и то, что вы видели при нашем первом опыте (помните, там прошло 8 секунд между первым ударом метронома и появлением первой капли слюны), есть запаздывание, вмешательство тормозного периода и заслуживало бы точного термина «предварительный тормозной период». Поэтому для цели определения истинного латентного периода условных рефлексов необходимо пользоваться совпадающими рефлексами, когда безусловный раздражитель возможно скоро, через секунду или даже части секунды, следует за началом условного. И только при таких рефлексах, главным образом пользуясь двигательным компонентом наших рефлексов, можно убедиться, что величина латентного периода условных рефлексов действительно падает в категорию величин обыкновенного рефлекторного времени. Этими определениями у нас занимаются только теперь. Мы не придавали фундаментального значения определениютинного латентного периода условных рефлексов при установлении их рефлекторной натуры, считая главным доказательством этого их непременность и закономерность при известных условиях, так как и при обыкновенных рефлексах латентныериоды значительно колеблются, находясь в зависимости от тех или других центральных связей, или путей. Можно с правом допустить, не угрожая рефлекторной натуре условных рефлексов, что в больших полушариях эти связи сложнее.
Решение многих других вопросотносительно условных рефлексов, например вопрос о подлинном, беспрепятственном течении при них раздражительного процесса с самого его начала и т. п., также более или менее затрудняется вмешательством запаздывания. В этом отношении много помогает делу характер нервной системы разных животных. У иных из них, как это было указано выше, запаздывание развивается с трудом, и тогда раздражительный процесс или совсем не нарушается, или очень мало. Но и в руках экспериментатора есть некоторое средство ослаблять влияние запаздывания, ограничиваясь возможно коротким отставлением безусловного раздражителя от начала условного, если величина условного рефлекса вообще велика, т. е. если и за короткий срок она достаточна для сравнения при различных вариациях опыта. Но, с другой стороны, есть случаи, когда предварительный тормозной процесс представляет некоторую выгоду, дает возможность для решения некоторых вопросов, как с этим мы встретимся в следующей лекции. Таким образом приходится приспособляться, соответственно пользоваться имеющимися отношениями.
Познакомившись в последних трех лекциях с основным фактическим материалом относительно внутреннего торможения условных рефлексов, нетрудно усмотреть его огромную биологическую ценность. Этим торможением постоянно корригируется и совершенствуется сигнализационная деятельность больших полушарий. Если в данное время сигнальный, условный раздражитель повторно не сопровождается его, так сказать, безусловно-деловым раздражителем, он как убыточный для организма, напрасно вызывающий трату энергии, на некоторое, как правило короткое, время лишается своего физиологического действия. Если условно действующий агент, постоянно встречаясь с посторонним агентом, при условии этой встречи, не сопровождается безусловным раздражителем, он также теряет свое условное значение, но только в этой комбинации, являясь в таком случае как бы другим агентом. Наконец если условный агент всегда только в определенный момент после своего возникновения совпадает с действием безусловного раздражителя, то он при более или менее отдаленно предшествующих этому раздражителю моментах делается недействительным, проявляясь все в большей и большей степени по мере приближения к моменту присоединения безусловного раздражителя. Таким образом постоянно обнаруживается высшее приспособление организма к окружающим условиям, или, иначе говоря, более тонкое и точное уравновешивание организма с внешней средой.
Как ясно из всего сообщенного в последних трех лекциях, мы при угасании, условном торможении и запаздывании имеем перед собой образование условных тормозных рефлексов. Но эти же рефлексы могут быть получены еще и при совсем иной процедуре. Если с условными тормозными раздражителями одновременно на короткие сроки, но повторно, применять индифферентные агенты, то эти последние приобретают такое тормозную функцию, т. е. и сами по себе вызывают в коре процесс торможения. Этот предмет специально разработан Ю. В. Фольбортом. Прежде всего он делал посторонние агенты индифферентными, повторяя их перед животным до тех пор, пока они не переставали тормозить положительных условных рефлексов и растормаживать тормозных. Затем он присоединял их повторно на короткие сроки то к угашенным условным рефлексам, то к тормозным комбинациям. И после этого испытывал их действие, комбинируя их с положительными условными раздражителями, прияем и обнаруживал теперь их роль как условных тормозов. Так как эти испытания естественно приходилось ради точности результата повторять не один раз, то Фольборт один раз сопровождал пробную комбинацию безусловным рефлексом, в другой - нет, делая это строго поочередно, чтобы при этих повторных испытаниях комбинация не получила благодаря им постоянного, определенного, то положительного, то отрицательного характера. Но, конечно, бесспорнее и убедительнее была та форма опыта, когда он при испытаниях всегда сопровождал пробную комбинацию безусловным рефлексом, а торможение, несмотря на это, при повторении опыта, все же резко выступало.
Вот эти опыты.
На собаке применялся натуральный условный пищевой рефлекс. Когда он угашался до нуля, к нему повторно присоединялись сделанные раньше индифферентными удары метронома, а затем при продолжении этого приема изредка пробовалась комбинация ударов метронома с положительным пищевым рефлексом, едой.
О п ы т 5 IX 1911 после 10 совпадений ударов метронома с угашенным условным рефлексом.
 Опыт 18 XII 1911 после 26 совпадений ударов метронома с угашенным условным рефлексом.
Опыт 18 XII 1911 после 26 совпадений ударов метронома с угашенным условным рефлексом.
 Совершенно очевидно, что ранее индифферентные удары метронома после совместных применений с угашенным условным раздражителем стали тормозящим агентом. Эти новые, иначе полученные, условные тормозные раздражители оказались по своим свойствам вполне сходными с теми, которые мы изучили при угасании, условном торможении и запаздывании.
Новый условный тормоз, полученный с одним угашенным рефлексом, действует тормозящим образом и на другие условные рефлексы.
О п ы т 15 III 1911. Другая собака. Удары метронома совпадали с угашенным условным натуральным пищевым рефлексом, а затем применялись вместе с запахом камфоры, искусственным условным пищевым раздражителем.
Совершенно очевидно, что ранее индифферентные удары метронома после совместных применений с угашенным условным раздражителем стали тормозящим агентом. Эти новые, иначе полученные, условные тормозные раздражители оказались по своим свойствам вполне сходными с теми, которые мы изучили при угасании, условном торможении и запаздывании.
Новый условный тормоз, полученный с одним угашенным рефлексом, действует тормозящим образом и на другие условные рефлексы.
О п ы т 15 III 1911. Другая собака. Удары метронома совпадали с угашенным условным натуральным пищевым рефлексом, а затем применялись вместе с запахом камфоры, искусственным условным пищевым раздражителем.
 Новый условный тормоз обнаруживает свое действие не только при одновременном применении с положительными условными раздражителями, но и последовательно.
При его повторениях он также суммируется, и, наконец, его действие может растормаживаться.
Итак, оказывается: если индифферентные раздражения попадают в кору полушарий, когда там господствует тормозной процесс, то они хронически получаютормозную функцию, т. e. когда ими потом действуют на деятельные пункты коры, они производят в них тормозной процесс.
Однако не следует упускать из виду и то, что, как увидим это потом, посторонние раздражители, повторяемые без дальнейших последствий для животного, так же и сами по себе одни ведут к развитию в коре тормозного состояния. Следовательно, при только что сообщенных опытах должно быть специально проконтролировано, насколько главное участие в образовании новых условных тормозов принимают именно условные тормозные рефлексы, с которыми повторно сочетаются посторонние раздражители.
Новый условный тормоз обнаруживает свое действие не только при одновременном применении с положительными условными раздражителями, но и последовательно.
При его повторениях он также суммируется, и, наконец, его действие может растормаживаться.
Итак, оказывается: если индифферентные раздражения попадают в кору полушарий, когда там господствует тормозной процесс, то они хронически получаютормозную функцию, т. e. когда ими потом действуют на деятельные пункты коры, они производят в них тормозной процесс.
Однако не следует упускать из виду и то, что, как увидим это потом, посторонние раздражители, повторяемые без дальнейших последствий для животного, так же и сами по себе одни ведут к развитию в коре тормозного состояния. Следовательно, при только что сообщенных опытах должно быть специально проконтролировано, насколько главное участие в образовании новых условных тормозов принимают именно условные тормозные рефлексы, с которыми повторно сочетаются посторонние раздражители.
Лекция седьмая. Анализаторная и синтезирующая работа больших полушарии: а) первоначальный обобщенный характер условных раздражителей; 6) дифференцировочное торможение
Мм. гг.! Условнорефлекторные агенты, беспрерывно сигнализируя непосредственно благоприятствующие и разрушающие влияния окружающей среды на организм, представляют собой, в соответствии с бесконечным разнообразием и колебаниями этой среды, как мельчайшие ее элементы, так и то большие, то меньшие комплексы из них. А это делается возможным только благодаря тому, что нервная система обладает как механизмами, выделяющими для организма из сложности среды отдельные ее элементы, - анализаторными механизмами, так и механизмами. соединяющими, сливающими эти элементы для организма в те или другие комплексы, - механизмами синтезирующими. Следовательно, при изучении высшей нервной деятельности, деятельности коры больших полушарий, мы должны встретиться с явлениями как нервного анализа, так и нервного синтеза, к чему мы теперь и приступаем. Нервная система есть всегда больший или меньший комплекс анализаторных приборов, анализаторов. Оптический отдел выделяет для организма световые колебания, акустический - воздушные, и т. д. В свою очередь каждый из этих отделов дробит соответствующую часть внешнего мира на длиннейший ряд отдельных элементов. К каждому данному анализатору должны быть отнесены как периферические приборы всевозможных афферентных нервов (трансформаторы, из которых каждый превращает в нервный процесс только определенную энергию), так и сами нервы и клеточные мозговые концы. Понятно отсюда, в анализаторной работе участвуют как те, так и другие. Более низкие степени анализа свойственны, конечно, и низшим отделам нервной системы (как и малодифференцированному веществу животных без нервной системы), так как и организм, лишенный головного мозга, отвечает очень различно на различные по месту, интенсивности и качеству раздражения его внешней поверхности. Но высший, тончайший анализ, на который способно данное животное, достигается только при помощи больших полушарий. Ясно, что усложнение связи животного организма с окружающим миром, все более разнообразное и более точное приспособление к внешним обстоятельствам, более совершенное уравновешивание организмов внешней среды идет параллельно и неразрывно с этой все прогрессирующей анализаторной деятельностью нервной системы. В современной физиологии изучение анализаторной деятельности составляет очень важный отдел так называемой физиологии органов чувств. Этот отдел представляет огромный интерес и является в высшей степени содержательным, отчасти, может быть, потому, что имел привилегию привлекать на себя внимание величайших физиологов, с гениальным Гельмгольцем во главе. В этом отделе заложены прочные основы учения о деятельности как периферических конструкций анализаторов, так и их мозговых концов, указаны многие пределы анализаторной работы (у человека), разъяснены некоторые сложные случаи этой работы и установлены многие частные правила, относящиеся к ней. Но весь этот огромный материал, субъективный в преобладающей его части, построен на наших ощущениях, этих наипростейших субъективных сигналах объективных отношений организма к внешнему миру. И это составляет значительный недостаток этого отдела. Из него таким образом исключено все то, что должно дата физиологии изучение разнообразия и развития анализаторной деятельности в животном царстве; вследствие этого же натуралист лишен неисчерпаемой возможности экспериментирования, которая представляется ему опытами над животными. Условные рефлексы все изучение важнейшей функции нервной системы, функции анализа, отдают в руки чистой физиологии, строго объективного, естественно-научного исследования. С помощью их удобно и точно определяются объем и предел анализаторной деятельности разных животных и выясняются законы этой деятельности. Пусть сейчас физиология анализаторов у животных еще очень скудна, но ее содержание все растет и будет расти все стремительнее, расширяя и углубляя наше знание о связи животного организма с окружающим миром. Как можно объективно, по видимкциям, изучать у животных работу анализаторов? Как уже мной кратко указывалось раньше, всякое колебание окружающей животное среды влечет за собой если не специальные - врожденные или приобретенные - реакции, то вообще ориентировочную реакцию, исследовательский рефлекс. Он, этот рефлекс, прежде всего и может служить для констатирования того, в какой степени нервная система данного животного может отличить одно от другого. Если, положим, в обычной среде между прочим имеется определенный звук - тон, то хотя бы и небольшое изменение его высоты должно вызвать и вызывает на себя ориентировочную реакцию, т. е. определенную постановку животным в отношении звука ушного аппарата. То же самое относится и ко всяческим другим раздражениям, простым или сложным, при малейших их изменениях. Но это, конечно, только в том случае, если анализаторный аппарат, по тонкости его конструкции, в состоянии отметить происшедшую разницу. В таком случае исследовательский рефлекс может служить для исследования или сам по себе, или, вследствие влияния его на условные рефлексы как чувствительнейшие реакции организма, в виде то тормозящего, то растормаживающего агента. Однако как постоянный прием для изучения анализаторной деятельности нервной системы исследовательский рефлекс, несмотря на его часто большую чувствительность, представляет и много неудобств. Из них главнейшее то, что он при некоторых слабых раздражениях очень летуч, не может быть повторяемым, чем существенно и подрывается возможность точного исследования с его помощью. Наоборот, условный рефлекс сам по себе является объектом, в высшей степени соответствующим цели данного изучения. Из строго определенного внешнего агента мы делаем обыкновенной процедурой специальный условный раздражитель и повторсниями постоянно утверждаем его в этой роли, а ближайший к нему по интенсивности, месту или качеству агент, при известных условиях, может оставаться постоянно без этой специальной реакции и, таким образом, легко и точно отличается от него. Так как в действительности анализаторная и синтезирующая работы нервной системы постоянно встречаются и перемежаются между собой и так как анализу внешних агентов с помощью условных рефлексов предшествует особенное нервное явление - род синтетической деятельности, то им прежде всего и приходится заняться, Всякий агент, из которого мы образуем наш условный раздражитель, непременно носит обобщенный характер. Если нами сделан условный раздражитель, например из тона в 1000 колебаний в секунду, то вместе с тем и многие другие тоны сами по себе получают то же условное действие, но тем меньшее, чем они дальше отстоят по высоте вверх и вниз от тона в 1000 колебаний. Точно так же, если механическое раздражение определенного места кожи сделано условным раздражителем, то такое же действие принадлежит затем и другим местам кожи тем в меньшем размере, чем дальше эти места находятся от места, на котором вырабатывался условный рефлекс. Тот жё факт повторяется при раздражениях других рецепторных поверхностей. Описанное явление с биологической точки зрения можно было бы понять так, что натуральные агенты большей частью не оказываются строго определенными, a скорее постоянно колеблются, меняются, переходя из одной родственной группы в другую. Возьмем для примера звук, издаваемый животнымврагом и являющийся условным раздражителем оборонительного рефлекса животного-жертвы. Он может представлять большие колебания в высоте, силе и составе в зависимости от напряжения голосового аппарата, расстояния, резонанса. Дальше мы встретились при условных рефлексах и с таким обобщением, жизненное значение которого не было ясно прямо. До сих пор мы имели дело с обобщением условного раздражителя в пределах одного анализатора. В случае условных следовых раздражителей, именно образованных на поздних следах (13 минуты спустя после прекращения действия внешнего агента), приходится видеть совсем другое. Здесь имеется обобщение универсальное. Мы образовали условный раздражитель, например, из механического раздражения кожи, а у нас в качестве условных раздражителей начинают действовать агенты, относящиеся к другим анализаторам и не стоящие ни в какое связи с нашим рефлексом. Я позволю себе остановить ваше внимание на этом факте несколько дольше, так как онедставляет интерес в отношении его разработки. Вот примеры этих опытов из работы Гроссмана. Механическое раздражение кожи - условный следовой (через минуту по прекращении наличного раздражения) кислотный раздражитель. Холод и тон применяются впереые.
 Мы видим, что и агенты, никогда ранее не приходившие в связь с действием кислоты, оказались возбудителями кислотной реакции. Мы замечаем вместе с тем, что впервые испробованные агенты действуют, так сказать, по той же программе, как и выработанный следовой раздражитель, т. е. действу: от не во время их применения, а лишь главным или исключительным образом по прекращении их. Это, конечно, с самого начала склоняет к заключению, что здесь дело идет об обобщенных рефлексах, воспроизводящихся по типу выработанного следового рефлекса. Но этого, разумеется, мало, и, ввиду особенности факта, мы подвергли его многократному переисследованию. Принимая во внимание все нам известное относительно условных рефлексов, мы сделали еще два возможных предположения о ближайшем механизме разбираемого факта.
Ta Так как следовой рефлекс на поздних следах образуется нелегко и нескоро, то нельзя при этом не заметить, что до образования его очень быстро образуются условные рефлексы на посторонние раздражения, случайно совпадающие с применением безусловного раздражителя и идущие от самого экспериментатора. Здесь эта опасность особенно велика. Поэтому эти опыты были повторены с удалением экспериментатора из комнаты, где находилась собака, и в новом специальном здании, чем было совершенно исключено всякое несознаваемое влияние со стороны экспериментатора на животное. Но чрезвычайная обобщенность следового рефлекса оставалась и тут.
Второе предположение, казавшееся тоже очень серьезным, состояло в следующем. При образовании у собак в лаборатории в первый раз условных рефлексов соответствующий рефлекс прежде всего образуется на всю лабораторную обстановку, начиная с привода животного в экспериментальную комнату. Его можно было бы назвать условным обстановочным рефлексом. Затем, когда начинает появляться рефлекс на применяемый нами специальный агент, элементы обстановки постепенно теряют свое условное действие, очевидно в силу развития на них постепенно внутреннего торможения. Но это торможение, особенно на первых порах, легко и растормаживается всякими экстренными посторонними агентами. Вот относящийся сюда яркий случаи, очень обычный в лаборатории при старой манере, когда экспериментатор и собака находились вместе в одной комнате. Обстановочный рефлекс, когда слюна текла сама по себе, миновал, и теперь на фоне покоя железы имеется свой специальный условный рефлекс. Но стоило мне впервые зайти в комнату для ознакомления с положением дела, и у собаки наступало сплошное слюноотделение, пока я находился в комнате. Я как экстренный посторонний агент растормаживал уже заторможенный обстановочный рефлекс. Мы предположили: не есть ли то, что мы называем обобщенным следовым рефлексом, растормаживание обстановочного рефлекса? Но при проверке и это предположение должно было отпасть. Во-первых, чрезвычайное обобщение следовых раздражителей получается легко постоянно у собак, у которых обстановочный рефлекс давно и прочно заторможен, так что почти невозможно его растормозить. Во-вторых, нами проконтролировано особенно строго следующее обстоятельство. Прошу припомнить, что при запаздывании растормаживание недеятельной фазы рефлекса происходило сейчас же, как пускался в ход посторонний раздражитель. Здесь же при следовых рефлексах этот раздражитель производит эффект почти всегда только по его прекращении. Если, настаивая на рассматриваемом предположении, принимать, что в данном случае все посторонние раздражители действуют почему-то всегда как очень сильные и потому не растормаживают, а еще глубже при их наличии тормозят обстановочный рефлекс (как это было и на запаздывающих рефлексах) и растормаживают только своими следами как слабеющим раздражением, то против такого изысканного допущения говорит следующее. При повторном применении сильных посторонних раздражителей их тормозящее действие непременно постепенно падает, и затем наступает фаза, когда они прямо только растормаживают, что мы и видели при запаздывающих рефлексах. Здесь при следовых рефлексах этого никогда не наблюдается, а многократновторяющиеся посторонние раздражители производят слюноотделение только после. Наконец, в-третьих, при следовых рефлексах после применения посторонних раздражителей, которечно, не сопровождаются безусловным раздражителем, специальный условный раздражитель временно слабеет, даже доходит до нуля, что было бы совершенно непонятно, если бы здесь дело шло о растормаживании обстановочного рефлекса. При обобщенном следовом рефлексе это ослабление - простой случай угасания после неподкрепления.
Итак, при образовании условных рефлексов сперва определенный безусловный рефлекс связывается с массой внешних агентов, один раз благодаря внешней среде, действующей на организм совокупностью ее элементов (условный синтетический обстановочный рефлекс), в другой раз в силу свойств самой нервной системы, когда внешнему элементарному агенту ею придается в качестве условного раздражителя обобщенный в большей или меньшей мере характер. Этот факт, как это частью ясно прямо, частью указано выше, конечно, оправдывается действительными отношениями животного к окружающему миру. Но вместе с тем очевидно, что тот же факт должен иметь лишь ограниченное и временное значение, когда при известных условиях возможная, приблизительная и общая связь заменяется действительной, точной и специальной связью.
Как же происходит специализация условного раздражителя, дифференцирование внешних агентов? Сначала нам казалось, что здесь имеют место два приема. Один - это только многократное повторение определенного агента в качестве условного раздражителя с постоянным подкреплением безусловным рефлексом. Другой - перемежающееся противопоставление этого определенного, постоянно покрепляемого, условного раздражителя с близким к нему агентом, но не сопровождаемым безусловным раздражителем. В настоящее время мы склонны признавать действительность только последнего приема. С одной стороны, мы имели условные раздражители, повторенные тысячекратно и которые, однако, через это одно не делались узко специализированными. С другой - было замечено, что даже однократная проба каждого из родственных агентов без подкрепления и редкое (через дни и даже недели) такое же (т. е. без подкрепления) применение ряда их (каждый раз все нового агента), при повторениях с подкреплением основного условного раздражителя, ведет, однако, к специализированию его. Поэтому мы постоянно пользовались вторым приемом, во всяком случае несравнимо скорее приводящим к цели дифференцировки агентов.
Мы прежде всего остановимся на подробностях хода процесса дифференцировки внешних агентов при помощи условных рефлексов. Здесь на первом месте должен быть поставлен факт, казавшийся нам в свое время довольно загадочным. Когда по образовании условного раздражителя из определенного внешнего агента впервые пробуют другие агенты, очень близкие к нему, например тоны, соседние с тоном-раздражителем, то часто получают от них условнорефлекторный эффект, резко меньший, чем от выработанного условного раздражителя. Но затем при повторении их, конечно без подкрепления, этот эффект быстро увеличивается, сравнивается с эффектом условного раздражителя и потом только постепенно начинает уменьшаться, доходя до нуля. Значит, сперва разница между агентами обнаруживается сразу, затем она почему-то исчезает и потом только снова медленно проявляется и, наконец, становится абсолютной. Как понимать этот факт? Мы воспользовались для этого сходством теперешнего факта с подобным фактом, ранее нами анализированным. Вы помните, что при выработке условного торможения наблюдалось такое же отношение. Когда впервые к условному раздражителю присоединяется новый агент, из которого потом образуется условный тормоз, то эта комбинация производит или эффект, свойственный условному раздражителю, но небольшой, или даже остается совсем бездейственной. Но затем, несмотря на то, что мы не сопровождаем ее безусловным рефлексом, она снова и быстро обнаруживает полное действие и лишь позже при повторении постепенно доходит до нуля. При условном торможении мы объяснили - и с полным правом - это явление так, что прибавочный агент вызвал ориентировочную реакцию и сейчас же затормозил действие условного раздражителя, с которым он был комбинирован. При повторении ориентировочная реакция быстро ослабела, и действие условного раздражителя временно восстановилось, чтобы при дальнейших повторениях постепенно быть подавленным развивающимся торможением. Мы и здесь можем представить себе, что сходственные агенты, близкие к агенту условного рефлекса, как бы состоят из двух частей (как и тормозная комбинация) - одной общей с условным раздражителем и другой особенной. Первая есть причина того, что близкие агенты действуют подобно выработанному условному раздражителю. Вторая сначала дает основание для временного ориентировочного рефлекса, тормозящего условный эффект первой части сейчас же и тоже на время, а впоследствии - для развития постоянной и окончательной дифференцировки близких агентов.
Be Веским доказательством основательности нашего понимания явлений при начале выработки дифференцировок является то, что сходство в ходе этих явлений при развитии условного торможения и выработке дифференцировок простирается и на имеющиеся здесь подробности. Как в том, так и в другом случае встречаются одни и те же вариации. Первоначальное ослабление эффекта близких агентов иногда переходит на короткое время в усиление сравнительно с тем, что дает специальный условный раздражитель, чтобы затем, вернувшись величине последнего, постепенно уменьшаясь, кончить нулем. Всего чаще за первоначальным ослаблением следует возврат почти к величине условного раздражителя, а потом развитие дифференцировки. Редко без этих колебаний прямо наступает постепенное образование дифференцировки, и, наконец, тоже редок непосредственный переход первоначального ослабления в прогрессирующее дифференцирование.
Так как при описании выработки условного торможения я только сообщил об их вариациях, не приведя протоколов опытов, то здесь могу с избытком пополнить этот дефект. В данном случае (работа М. М. Губергрица) эти вариации были прослежены в особенности систематично.
Ряд опытов на одном и том же животном.
Вертящийся в направлении часовой стрелки предмет -- условный пищевой раздражитель, тот же предмет, вертящийся в обратном направлении, - дифференцируемый агент.
Мы видим, что и агенты, никогда ранее не приходившие в связь с действием кислоты, оказались возбудителями кислотной реакции. Мы замечаем вместе с тем, что впервые испробованные агенты действуют, так сказать, по той же программе, как и выработанный следовой раздражитель, т. е. действу: от не во время их применения, а лишь главным или исключительным образом по прекращении их. Это, конечно, с самого начала склоняет к заключению, что здесь дело идет об обобщенных рефлексах, воспроизводящихся по типу выработанного следового рефлекса. Но этого, разумеется, мало, и, ввиду особенности факта, мы подвергли его многократному переисследованию. Принимая во внимание все нам известное относительно условных рефлексов, мы сделали еще два возможных предположения о ближайшем механизме разбираемого факта.
Ta Так как следовой рефлекс на поздних следах образуется нелегко и нескоро, то нельзя при этом не заметить, что до образования его очень быстро образуются условные рефлексы на посторонние раздражения, случайно совпадающие с применением безусловного раздражителя и идущие от самого экспериментатора. Здесь эта опасность особенно велика. Поэтому эти опыты были повторены с удалением экспериментатора из комнаты, где находилась собака, и в новом специальном здании, чем было совершенно исключено всякое несознаваемое влияние со стороны экспериментатора на животное. Но чрезвычайная обобщенность следового рефлекса оставалась и тут.
Второе предположение, казавшееся тоже очень серьезным, состояло в следующем. При образовании у собак в лаборатории в первый раз условных рефлексов соответствующий рефлекс прежде всего образуется на всю лабораторную обстановку, начиная с привода животного в экспериментальную комнату. Его можно было бы назвать условным обстановочным рефлексом. Затем, когда начинает появляться рефлекс на применяемый нами специальный агент, элементы обстановки постепенно теряют свое условное действие, очевидно в силу развития на них постепенно внутреннего торможения. Но это торможение, особенно на первых порах, легко и растормаживается всякими экстренными посторонними агентами. Вот относящийся сюда яркий случаи, очень обычный в лаборатории при старой манере, когда экспериментатор и собака находились вместе в одной комнате. Обстановочный рефлекс, когда слюна текла сама по себе, миновал, и теперь на фоне покоя железы имеется свой специальный условный рефлекс. Но стоило мне впервые зайти в комнату для ознакомления с положением дела, и у собаки наступало сплошное слюноотделение, пока я находился в комнате. Я как экстренный посторонний агент растормаживал уже заторможенный обстановочный рефлекс. Мы предположили: не есть ли то, что мы называем обобщенным следовым рефлексом, растормаживание обстановочного рефлекса? Но при проверке и это предположение должно было отпасть. Во-первых, чрезвычайное обобщение следовых раздражителей получается легко постоянно у собак, у которых обстановочный рефлекс давно и прочно заторможен, так что почти невозможно его растормозить. Во-вторых, нами проконтролировано особенно строго следующее обстоятельство. Прошу припомнить, что при запаздывании растормаживание недеятельной фазы рефлекса происходило сейчас же, как пускался в ход посторонний раздражитель. Здесь же при следовых рефлексах этот раздражитель производит эффект почти всегда только по его прекращении. Если, настаивая на рассматриваемом предположении, принимать, что в данном случае все посторонние раздражители действуют почему-то всегда как очень сильные и потому не растормаживают, а еще глубже при их наличии тормозят обстановочный рефлекс (как это было и на запаздывающих рефлексах) и растормаживают только своими следами как слабеющим раздражением, то против такого изысканного допущения говорит следующее. При повторном применении сильных посторонних раздражителей их тормозящее действие непременно постепенно падает, и затем наступает фаза, когда они прямо только растормаживают, что мы и видели при запаздывающих рефлексах. Здесь при следовых рефлексах этого никогда не наблюдается, а многократновторяющиеся посторонние раздражители производят слюноотделение только после. Наконец, в-третьих, при следовых рефлексах после применения посторонних раздражителей, которечно, не сопровождаются безусловным раздражителем, специальный условный раздражитель временно слабеет, даже доходит до нуля, что было бы совершенно непонятно, если бы здесь дело шло о растормаживании обстановочного рефлекса. При обобщенном следовом рефлексе это ослабление - простой случай угасания после неподкрепления.
Итак, при образовании условных рефлексов сперва определенный безусловный рефлекс связывается с массой внешних агентов, один раз благодаря внешней среде, действующей на организм совокупностью ее элементов (условный синтетический обстановочный рефлекс), в другой раз в силу свойств самой нервной системы, когда внешнему элементарному агенту ею придается в качестве условного раздражителя обобщенный в большей или меньшей мере характер. Этот факт, как это частью ясно прямо, частью указано выше, конечно, оправдывается действительными отношениями животного к окружающему миру. Но вместе с тем очевидно, что тот же факт должен иметь лишь ограниченное и временное значение, когда при известных условиях возможная, приблизительная и общая связь заменяется действительной, точной и специальной связью.
Как же происходит специализация условного раздражителя, дифференцирование внешних агентов? Сначала нам казалось, что здесь имеют место два приема. Один - это только многократное повторение определенного агента в качестве условного раздражителя с постоянным подкреплением безусловным рефлексом. Другой - перемежающееся противопоставление этого определенного, постоянно покрепляемого, условного раздражителя с близким к нему агентом, но не сопровождаемым безусловным раздражителем. В настоящее время мы склонны признавать действительность только последнего приема. С одной стороны, мы имели условные раздражители, повторенные тысячекратно и которые, однако, через это одно не делались узко специализированными. С другой - было замечено, что даже однократная проба каждого из родственных агентов без подкрепления и редкое (через дни и даже недели) такое же (т. е. без подкрепления) применение ряда их (каждый раз все нового агента), при повторениях с подкреплением основного условного раздражителя, ведет, однако, к специализированию его. Поэтому мы постоянно пользовались вторым приемом, во всяком случае несравнимо скорее приводящим к цели дифференцировки агентов.
Мы прежде всего остановимся на подробностях хода процесса дифференцировки внешних агентов при помощи условных рефлексов. Здесь на первом месте должен быть поставлен факт, казавшийся нам в свое время довольно загадочным. Когда по образовании условного раздражителя из определенного внешнего агента впервые пробуют другие агенты, очень близкие к нему, например тоны, соседние с тоном-раздражителем, то часто получают от них условнорефлекторный эффект, резко меньший, чем от выработанного условного раздражителя. Но затем при повторении их, конечно без подкрепления, этот эффект быстро увеличивается, сравнивается с эффектом условного раздражителя и потом только постепенно начинает уменьшаться, доходя до нуля. Значит, сперва разница между агентами обнаруживается сразу, затем она почему-то исчезает и потом только снова медленно проявляется и, наконец, становится абсолютной. Как понимать этот факт? Мы воспользовались для этого сходством теперешнего факта с подобным фактом, ранее нами анализированным. Вы помните, что при выработке условного торможения наблюдалось такое же отношение. Когда впервые к условному раздражителю присоединяется новый агент, из которого потом образуется условный тормоз, то эта комбинация производит или эффект, свойственный условному раздражителю, но небольшой, или даже остается совсем бездейственной. Но затем, несмотря на то, что мы не сопровождаем ее безусловным рефлексом, она снова и быстро обнаруживает полное действие и лишь позже при повторении постепенно доходит до нуля. При условном торможении мы объяснили - и с полным правом - это явление так, что прибавочный агент вызвал ориентировочную реакцию и сейчас же затормозил действие условного раздражителя, с которым он был комбинирован. При повторении ориентировочная реакция быстро ослабела, и действие условного раздражителя временно восстановилось, чтобы при дальнейших повторениях постепенно быть подавленным развивающимся торможением. Мы и здесь можем представить себе, что сходственные агенты, близкие к агенту условного рефлекса, как бы состоят из двух частей (как и тормозная комбинация) - одной общей с условным раздражителем и другой особенной. Первая есть причина того, что близкие агенты действуют подобно выработанному условному раздражителю. Вторая сначала дает основание для временного ориентировочного рефлекса, тормозящего условный эффект первой части сейчас же и тоже на время, а впоследствии - для развития постоянной и окончательной дифференцировки близких агентов.
Be Веским доказательством основательности нашего понимания явлений при начале выработки дифференцировок является то, что сходство в ходе этих явлений при развитии условного торможения и выработке дифференцировок простирается и на имеющиеся здесь подробности. Как в том, так и в другом случае встречаются одни и те же вариации. Первоначальное ослабление эффекта близких агентов иногда переходит на короткое время в усиление сравнительно с тем, что дает специальный условный раздражитель, чтобы затем, вернувшись величине последнего, постепенно уменьшаясь, кончить нулем. Всего чаще за первоначальным ослаблением следует возврат почти к величине условного раздражителя, а потом развитие дифференцировки. Редко без этих колебаний прямо наступает постепенное образование дифференцировки, и, наконец, тоже редок непосредственный переход первоначального ослабления в прогрессирующее дифференцирование.
Так как при описании выработки условного торможения я только сообщил об их вариациях, не приведя протоколов опытов, то здесь могу с избытком пополнить этот дефект. В данном случае (работа М. М. Губергрица) эти вариации были прослежены в особенности систематично.
Ряд опытов на одном и том же животном.
Вертящийся в направлении часовой стрелки предмет -- условный пищевой раздражитель, тот же предмет, вертящийся в обратном направлении, - дифференцируемый агент.
 Затем с колебаниями дифференцируемый агент доходит до нуля действия.
Другая собака. Тон - условный пищевой раздражитель, полутон к нему - дифференцируемый агент.
Затем с колебаниями дифференцируемый агент доходит до нуля действия.
Другая собака. Тон - условный пищевой раздражитель, полутон к нему - дифференцируемый агент.
 Далее с колебаниями к тринадцатому повторению полутон доходит до нуля.
Та же собака, что в первых опытах. Появление круга перед глазами собаки - условный пищевой раздражитель, появление квадрата той же площади и того же освещения - дифференцируемый агент.
Далее с колебаниями к тринадцатому повторению полутон доходит до нуля.
Та же собака, что в первых опытах. Появление круга перед глазами собаки - условный пищевой раздражитель, появление квадрата той же площади и того же освещения - дифференцируемый агент.

 Затем с колебаниями к одиннадцатому повторению квадрат становится совершенно недействительным.
При дифференцировании внешних агентов с помощью условных рефлексов, помимо только что описанной начальной стадии, связанной с вмешательством ориентировочной реакции, привлекли к себе наше внимание и многие другие стороны этого нервного акта.
Можно, конечно, производить сразу дифференцировку агента условного рефлекса от очень близкого к нему агента, но можно подходить к ней и постепенно, начиная сперва с дифференцирования отдаленных агентов. Оказалась огромная разница между одним и другим приемом. Мы начинаем прямо с дифференцирорования близкого агента, и, несмотря на значительное число противопоставлений одного агента другому, искомой разницы не оказывается. Переходим тогда к более отдаленным и потом постепенно возвращаемся к первому -- теперь через этапы достигнутых дифференцировок на более отдаленных агентах, в целом в гораздо более короткий срок, имеем дифференцировку близкого агента.
Вот подтвержающие это отношение опыта тоже из работы Губергрица.
У У одной собаки условным пищевым раздражителем был сделан круг из белой бумаги, появляющимия перед ее глазами. От него начали дифференцировать круг из бумаги № 10 циммермановской коллекции (50 оттенков от белого цвета до черного); 75 применений серого круга (№ 10), конечно без подкрепления, чередуемых с белым, постоянно подкрепляемым, не обнаружили ни малейших признаков различения. Тогда перешли к дифференцированию сперва более темного круга № 35, а затем последовательно к кругу № 25 и кругу № 15 и, наконец, вернулись к кругу № 10. Теперь полная дифференцировка была достигнута всего через 20 применений этих кругов, считая все вместе.
То же было проделано на другой собаке также с глазным раздражителем, но в другой форме. У нее условным пищевым раздражителем был сделан также круг, но от него дифференцировался эллипсис, из той же бумаги и той же величины площади с отношением полуосей 8 : 9. Несмотря на 70 противопоставлений, никакой дифференцировки не обнаружилось. Когда же после этого стали последовательно дифференцировать эллипсисы, начиная с эллипсиса с отношением осей 4:5 и переходя через эллипсисы 5:6, 6: 7, 7: 8, то уже после 18 применений, считая вместе все потребовавшиеся для выработки каждой из них, имелась дифференцировка и на эллипсис 8 : 9.
Если мы производим дифференцировку постепенно, начиная с отдаленных агентов, то скоро бросается в глаза следующий постоянный факт. При выработке дифференцировки, хотя бы и грубой, в первый раз дело идет довольно медленно, особенно если настаивать на абсолютном дифференцировании, т. е. на нуле эффекта от дифференцируемого агента. Но раз на первом этапе достигнута абсолютная или близкая к ней дифференцировка, то процедура дальнейшего дифференцирования происходит все быстрее, но опять несколько замедляясь, когда мы подходим к предельной дифференцировке.
Привожу один из примеров.
У собаки белый круг определенного диаметра - условный пищевой раздражитель, то же белые эллипсисы, той же величины площади с разными отношениями их полуосей - дифференцируемые агенты. Первый эллипсис с отношением полуосей 4 : 5 потребовал 24 применений (34 деления за 30 секунд действия дал круг, а эллипсис только 4). При следующем эллипсисе 5: 6 нужно было только троекратное его повторение, чтобы его эффект оказался даже нулевым. То же и со следующим эллипсисом 6 : 7.
Ход дифференцирования, выключая начальную фазу с участием ориентировочной реакции, может быть как почти прямолинейным, так и волнообразным, и последнее не всегда может быть отнесено к каким-нибудь внешним нарушающим влияниям, а, вероятно, связано с колебаниями внутреннего нервного процесса, при этом происходящего.
Прочность дифференцировки определяется по длине промежутка между применением условного раздражителя и отдифференцированного от него агента, при котором дифференцировка сохраняется. Сначала эти промежутки короткие, затем они становятся все длиннее. Мы считали дифференцировку окончательно выработанной, когда она через сутки, поставленная в опыте на первом месте, т. е. ранее других раздражений, оказывалась полной.
Наши неоднократно повторенные опыты показали, что дифференцирование различных агентов достигает приблизительно одинаковых степеней, применяются ли они в качестве положительных условных раздражителей или отрицательных, тормозных.
Так же могут и в той же степени дифференцироваться агенты и в роли следовых условных раздражителей, причем тоже как положительных, так и тормозных.
Вот пример дифференцировки следового тормоза из работы Фролова.
У собаки удары метронома (104 удара в минуту) - условный пищевой раздражитель. Определенный тон органной трубы (№ 16), продолжавшийся 15 секунд и прекращаемый за минуту до ударов метронома, - условный тормоз. Такая же комбинация ударов метронома с соседним тоном той же органной трубы (№ 15) сопровождалась безусловным рефлексом и стала положительным раздражителем. Дифференцировка следового тормоза представилась в следующем виде.
Затем с колебаниями к одиннадцатому повторению квадрат становится совершенно недействительным.
При дифференцировании внешних агентов с помощью условных рефлексов, помимо только что описанной начальной стадии, связанной с вмешательством ориентировочной реакции, привлекли к себе наше внимание и многие другие стороны этого нервного акта.
Можно, конечно, производить сразу дифференцировку агента условного рефлекса от очень близкого к нему агента, но можно подходить к ней и постепенно, начиная сперва с дифференцирования отдаленных агентов. Оказалась огромная разница между одним и другим приемом. Мы начинаем прямо с дифференцирорования близкого агента, и, несмотря на значительное число противопоставлений одного агента другому, искомой разницы не оказывается. Переходим тогда к более отдаленным и потом постепенно возвращаемся к первому -- теперь через этапы достигнутых дифференцировок на более отдаленных агентах, в целом в гораздо более короткий срок, имеем дифференцировку близкого агента.
Вот подтвержающие это отношение опыта тоже из работы Губергрица.
У У одной собаки условным пищевым раздражителем был сделан круг из белой бумаги, появляющимия перед ее глазами. От него начали дифференцировать круг из бумаги № 10 циммермановской коллекции (50 оттенков от белого цвета до черного); 75 применений серого круга (№ 10), конечно без подкрепления, чередуемых с белым, постоянно подкрепляемым, не обнаружили ни малейших признаков различения. Тогда перешли к дифференцированию сперва более темного круга № 35, а затем последовательно к кругу № 25 и кругу № 15 и, наконец, вернулись к кругу № 10. Теперь полная дифференцировка была достигнута всего через 20 применений этих кругов, считая все вместе.
То же было проделано на другой собаке также с глазным раздражителем, но в другой форме. У нее условным пищевым раздражителем был сделан также круг, но от него дифференцировался эллипсис, из той же бумаги и той же величины площади с отношением полуосей 8 : 9. Несмотря на 70 противопоставлений, никакой дифференцировки не обнаружилось. Когда же после этого стали последовательно дифференцировать эллипсисы, начиная с эллипсиса с отношением осей 4:5 и переходя через эллипсисы 5:6, 6: 7, 7: 8, то уже после 18 применений, считая вместе все потребовавшиеся для выработки каждой из них, имелась дифференцировка и на эллипсис 8 : 9.
Если мы производим дифференцировку постепенно, начиная с отдаленных агентов, то скоро бросается в глаза следующий постоянный факт. При выработке дифференцировки, хотя бы и грубой, в первый раз дело идет довольно медленно, особенно если настаивать на абсолютном дифференцировании, т. е. на нуле эффекта от дифференцируемого агента. Но раз на первом этапе достигнута абсолютная или близкая к ней дифференцировка, то процедура дальнейшего дифференцирования происходит все быстрее, но опять несколько замедляясь, когда мы подходим к предельной дифференцировке.
Привожу один из примеров.
У собаки белый круг определенного диаметра - условный пищевой раздражитель, то же белые эллипсисы, той же величины площади с разными отношениями их полуосей - дифференцируемые агенты. Первый эллипсис с отношением полуосей 4 : 5 потребовал 24 применений (34 деления за 30 секунд действия дал круг, а эллипсис только 4). При следующем эллипсисе 5: 6 нужно было только троекратное его повторение, чтобы его эффект оказался даже нулевым. То же и со следующим эллипсисом 6 : 7.
Ход дифференцирования, выключая начальную фазу с участием ориентировочной реакции, может быть как почти прямолинейным, так и волнообразным, и последнее не всегда может быть отнесено к каким-нибудь внешним нарушающим влияниям, а, вероятно, связано с колебаниями внутреннего нервного процесса, при этом происходящего.
Прочность дифференцировки определяется по длине промежутка между применением условного раздражителя и отдифференцированного от него агента, при котором дифференцировка сохраняется. Сначала эти промежутки короткие, затем они становятся все длиннее. Мы считали дифференцировку окончательно выработанной, когда она через сутки, поставленная в опыте на первом месте, т. е. ранее других раздражений, оказывалась полной.
Наши неоднократно повторенные опыты показали, что дифференцирование различных агентов достигает приблизительно одинаковых степеней, применяются ли они в качестве положительных условных раздражителей или отрицательных, тормозных.
Так же могут и в той же степени дифференцироваться агенты и в роли следовых условных раздражителей, причем тоже как положительных, так и тормозных.
Вот пример дифференцировки следового тормоза из работы Фролова.
У собаки удары метронома (104 удара в минуту) - условный пищевой раздражитель. Определенный тон органной трубы (№ 16), продолжавшийся 15 секунд и прекращаемый за минуту до ударов метронома, - условный тормоз. Такая же комбинация ударов метронома с соседним тоном той же органной трубы (№ 15) сопровождалась безусловным рефлексом и стала положительным раздражителем. Дифференцировка следового тормоза представилась в следующем виде.

 Надо прибавить, что эта дифференцировка была достигнута только благодаря крайней постепенности при ее выработке. Приходилось начинать секундных следов и с удаленных тонов.
Теперь пора определенно поставить вопрос: каким нервным процессом обусловливается переход общинного характера условного раздражителя к его крайней специализации, или, иначе говоря, дифференцирование близких агентов? После знакомства с внутренним торможением и после того, что уже сообщено выше в этой лекции, нетрудно догадаться, в чем тут дело, - и наши дальнейшие многочисленные опыты не оставляютсомнения в том, что в основании дифференцирования лежит тормозной процесс, т. е. как бы постепенное заглушение сначала широко возбужденного мозгового конца анализатора, исключая его мельчайшую часть, отвечающую данному условному раздражителю. Вот первое фактическое оправдание этого вывода. Указанной выше процедурой мыин агент - наш условный раздражитель - отдифференцировали от близкого к нему агента: он дает постоянный и полный условный эффект, отдифференцированный же является, повидимому, совершенно недействительным. Теперь после условного раздражителя применим этот последний, а за ним сейчас же или через некоторый промежуток повторим опять наш условный раздражитель - он будет иметь временно маленькое или никакого действия.
Вот пример из работы В. В. Белякова.
У собаки определенный тон органной трубы - условный пищевой раздражитель, от него прочно отдифференцирован тон на 1 часть выше.
Надо прибавить, что эта дифференцировка была достигнута только благодаря крайней постепенности при ее выработке. Приходилось начинать секундных следов и с удаленных тонов.
Теперь пора определенно поставить вопрос: каким нервным процессом обусловливается переход общинного характера условного раздражителя к его крайней специализации, или, иначе говоря, дифференцирование близких агентов? После знакомства с внутренним торможением и после того, что уже сообщено выше в этой лекции, нетрудно догадаться, в чем тут дело, - и наши дальнейшие многочисленные опыты не оставляютсомнения в том, что в основании дифференцирования лежит тормозной процесс, т. е. как бы постепенное заглушение сначала широко возбужденного мозгового конца анализатора, исключая его мельчайшую часть, отвечающую данному условному раздражителю. Вот первое фактическое оправдание этого вывода. Указанной выше процедурой мыин агент - наш условный раздражитель - отдифференцировали от близкого к нему агента: он дает постоянный и полный условный эффект, отдифференцированный же является, повидимому, совершенно недействительным. Теперь после условного раздражителя применим этот последний, а за ним сейчас же или через некоторый промежуток повторим опять наш условный раздражитель - он будет иметь временно маленькое или никакого действия.
Вот пример из работы В. В. Белякова.
У собаки определенный тон органной трубы - условный пищевой раздражитель, от него прочно отдифференцирован тон на 1 часть выше.
 Следовательно, после применения отдифференцированного тона в нервной системе остается на некоторое время тормозной процесс, который дает себя знать на испытанном последовательно условном раздражителе, уменьшая или совершенно уничтожая его эффект, иначе говоря, перед нами хорошо нам знакомое по ранним лекциям последовательное торможение.
Таким образом мы имеем четвертый случай внутреннего торможения, которое мы соответственно назвали дифференцировочным торможением. Может быть, было бы допуски моего отождествить и объединить общим прилагательным (именно, дифференцировочное) с условным торможением, так как в обоих случаях идет дело об устранении помощью торможения действия посторонних (простых или сложных) агентов, получивших это их действие без специальной выработки, только вследствие той или другой связи их с выработанным условным раздражителем.
Последовательное торможение, с которого мы начали при процессе дифференцирования, представляется одинаковым с последействием при условном торможении в том отношении, что оно также с повторениями делается все более вс времени ограниченным, простираясь на десятки минут вначале и, наконец, сокращаясь до секунд.
Здесь заслуживает еще особенного упоминания тот факт, что чем выше степень дифференцировки, т. е. чем более близки друг к другу дифференцируемые агентом значительнее последовательное торможение при прочих равных условиях.
Вот иллюстрация этого из работы Белякова.
Определенный тон - условный пищевой раздражитель, и того же тона - дифференцируемые агенты.
Следовательно, после применения отдифференцированного тона в нервной системе остается на некоторое время тормозной процесс, который дает себя знать на испытанном последовательно условном раздражителе, уменьшая или совершенно уничтожая его эффект, иначе говоря, перед нами хорошо нам знакомое по ранним лекциям последовательное торможение.
Таким образом мы имеем четвертый случай внутреннего торможения, которое мы соответственно назвали дифференцировочным торможением. Может быть, было бы допуски моего отождествить и объединить общим прилагательным (именно, дифференцировочное) с условным торможением, так как в обоих случаях идет дело об устранении помощью торможения действия посторонних (простых или сложных) агентов, получивших это их действие без специальной выработки, только вследствие той или другой связи их с выработанным условным раздражителем.
Последовательное торможение, с которого мы начали при процессе дифференцирования, представляется одинаковым с последействием при условном торможении в том отношении, что оно также с повторениями делается все более вс времени ограниченным, простираясь на десятки минут вначале и, наконец, сокращаясь до секунд.
Здесь заслуживает еще особенного упоминания тот факт, что чем выше степень дифференцировки, т. е. чем более близки друг к другу дифференцируемые агентом значительнее последовательное торможение при прочих равных условиях.
Вот иллюстрация этого из работы Белякова.
Определенный тон - условный пищевой раздражитель, и того же тона - дифференцируемые агенты.
 Но и помимо последействия дифференцировочное торможение также вполне сходствует с ранними случаями торможения.
Оно также суммируется при повторениях.
Вот опыты также из работы Белякова, поставленные на эту тему.
Другая собака. Тон - условный пищевой раздражитель, полутон ниже - дифференцируемый агент.
Но и помимо последействия дифференцировочное торможение также вполне сходствует с ранними случаями торможения.
Оно также суммируется при повторениях.
Вот опыты также из работы Белякова, поставленные на эту тему.
Другая собака. Тон - условный пищевой раздражитель, полутон ниже - дифференцируемый агент.
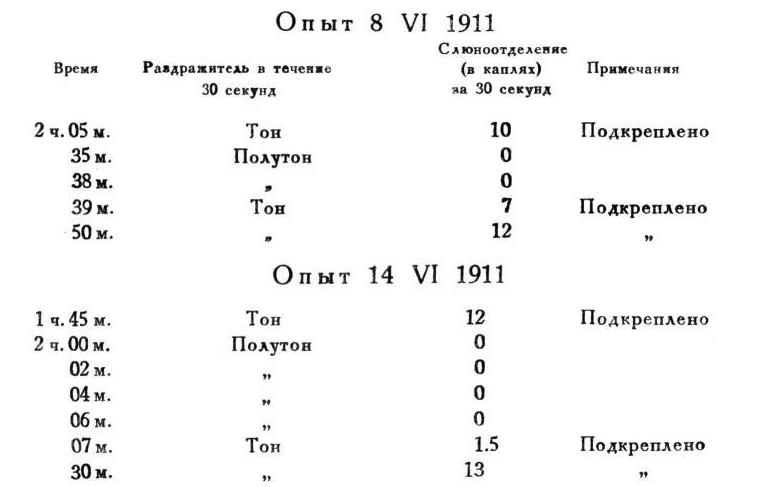 Дифференцировочное торможение так же, так сказать, отмеривается по размеру того раздражительного процесса, с которым имеет дело при выработке, и потому нарушается при увеличении интенсивности дифференцируемого агента и при изменении общей и местной возбудимости центральной нервной системы.
Если мы пользуемся пищевым рефлексом, то стоит только повысить пищевую возбудимость, делая опыт в более поздний час дня, т. е. ближе к сроку обычного корма собаки, или же заставить голодать животное лишний день, как ранняя вполне выработанная дифференцировка оказывается уже недостаточной.
Точно так же, если общую возбудимость центральной нервной системы повысить впрыскиванием кофеина, совершенная ранее дифференцировка нарушается. Вот опыт, сюда относящийся, из работы П. М. Никифоровского.
Механическое раздражение кожи на передней лапе - пищевой условный раздражитель, такое же раздражение кожи на спине - отдифференцированный агент.
Дифференцировочное торможение так же, так сказать, отмеривается по размеру того раздражительного процесса, с которым имеет дело при выработке, и потому нарушается при увеличении интенсивности дифференцируемого агента и при изменении общей и местной возбудимости центральной нервной системы.
Если мы пользуемся пищевым рефлексом, то стоит только повысить пищевую возбудимость, делая опыт в более поздний час дня, т. е. ближе к сроку обычного корма собаки, или же заставить голодать животное лишний день, как ранняя вполне выработанная дифференцировка оказывается уже недостаточной.
Точно так же, если общую возбудимость центральной нервной системы повысить впрыскиванием кофеина, совершенная ранее дифференцировка нарушается. Вот опыт, сюда относящийся, из работы П. М. Никифоровского.
Механическое раздражение кожи на передней лапе - пищевой условный раздражитель, такое же раздражение кожи на спине - отдифференцированный агент.
 Наконец дифференцировочное торможение, как и все остальные случаи внутреннего торможения, подвергается растормаживанию, т. е. и это торможение на короткое время устраняется, уступая место положительному эффекту под влиянием посторонних раздражений.
Вот два опыта из работы Белякова на одном и том же животном с разными посторонними раздражителями.
Тон (800 колебаний в секунду) - условный пищевой раздражитель, тона (812 колебаний в секунду) - основательно отдифференцированный агент. Оба посторонние раздражителя, бульканье и запах уксусного амила, испытанные отдельно, не сопровождались слюноотделением.
Наконец дифференцировочное торможение, как и все остальные случаи внутреннего торможения, подвергается растормаживанию, т. е. и это торможение на короткое время устраняется, уступая место положительному эффекту под влиянием посторонних раздражений.
Вот два опыта из работы Белякова на одном и том же животном с разными посторонними раздражителями.
Тон (800 колебаний в секунду) - условный пищевой раздражитель, тона (812 колебаний в секунду) - основательно отдифференцированный агент. Оба посторонние раздражителя, бульканье и запах уксусного амила, испытанные отдельно, не сопровождались слюноотделением.
 Интересно, что растормаживание можно констатировать и в фазе последействия дифференцировочного торможения.
О п ы т Белякова на том же животном, у которого никогда не наблюдалось случайного слюноотделения.
Интересно, что растормаживание можно констатировать и в фазе последействия дифференцировочного торможения.
О п ы т Белякова на том же животном, у которого никогда не наблюдалось случайного слюноотделения.
 Между примененными нами посторонними раздражителями были и такие, которые вызывали не ориентировочную реакцию, но специальный рефлекс, более сложный и резкий. В таком случае растормаживающее действие давало себя знать еще долго по прекращении самого раздражителя.
О пыт Белякова на том же животном. Посторонним раздражителем была игрушечная труба, производившая резкие разнообразные и в высшей степени неровные звуки. Собака при этом лает, дрожит, рвется из станка.
Между примененными нами посторонними раздражителями были и такие, которые вызывали не ориентировочную реакцию, но специальный рефлекс, более сложный и резкий. В таком случае растормаживающее действие давало себя знать еще долго по прекращении самого раздражителя.
О пыт Белякова на том же животном. Посторонним раздражителем была игрушечная труба, производившая резкие разнообразные и в высшей степени неровные звуки. Собака при этом лает, дрожит, рвется из станка.
 Все сообщенные опыты делают несомненным, что выработка дифференцировок основана на развитии внутреннего торможения в отношении дифференцируемых агентов.
На основании разных фактов, описанных в последних лекциях, необходимо придти к заключению, что между констатированием нервной системой разницы между внешними агентами вообще и дифференцированием тех же агентов при помощи условных рефлексов есть существенная разница. Первое обнаруживается раздражительным процессом в виде ориентировочной реакции, исследовательского рефлекса, только вторично влияющим на условные рефлексы или тормозящим, или растормаживающим образом. Второе выражается в развитии тормозного процесса, являющегося в результате, так сказать, борьбы между раздражением и торможением. Как увидим это позже, эта борьба часто бывает очень трудно. Мыслимо, что иногда она и непосильна, и тогда при этой борьбе не всегда дело может доходить до полной утилизации для общей деятельности организма результата действительного анализа внешних агентов. Если это так, то изучение анализаторной деятельности нервной системы помощью условных рефлексов также будет иметь свой недостаток. Во всяком случае это интересный вопрос.
Все сообщенные опыты делают несомненным, что выработка дифференцировок основана на развитии внутреннего торможения в отношении дифференцируемых агентов.
На основании разных фактов, описанных в последних лекциях, необходимо придти к заключению, что между констатированием нервной системой разницы между внешними агентами вообще и дифференцированием тех же агентов при помощи условных рефлексов есть существенная разница. Первое обнаруживается раздражительным процессом в виде ориентировочной реакции, исследовательского рефлекса, только вторично влияющим на условные рефлексы или тормозящим, или растормаживающим образом. Второе выражается в развитии тормозного процесса, являющегося в результате, так сказать, борьбы между раздражением и торможением. Как увидим это позже, эта борьба часто бывает очень трудно. Мыслимо, что иногда она и непосильна, и тогда при этой борьбе не всегда дело может доходить до полной утилизации для общей деятельности организма результата действительного анализа внешних агентов. Если это так, то изучение анализаторной деятельности нервной системы помощью условных рефлексов также будет иметь свой недостаток. Во всяком случае это интересный вопрос.
Лекция восьмая. Анализаторная и синтезирующая работа больших полушарий: в) примеры анализаторной работы; г) синтез и анализ одновременных комплексных раздражителей; д) синтез и анализ последовательных комплексных раздражителей
Мм. гг.! В прошлой лекции мы видели, как отд. льный агент внешнего мира, примененный в качестве условного раздражителя, сначала является обобщенным в большей или меньшей степени, но потом, благодаря возникновению при соответствующей процедуре тормозного процесса, все более и более специализируется, обособляется. Таким образом в окончательном результате создается хорошая экспериментальная возможность изучить объем и границы деятельности различных анализаторов животного. В этом отношении мы располагаем значительным материалом, касающимся многих анализаторов собаки. Относительно всех этих опытов надо сказать, что большие затруднения при постановке их возникают не с физиологоической стороны, а с физической, инструментальной стороны. Во многих случаях становится трудным достать или сконструировать такие физические приборы, которые вполне отвечали бы поставленной цели физиологического опыта. Задача наша сводится на абсолютное изолирование инструментом данного элементарного внешнего агента или той, или другой его интенсивности, а оказывается, например, почти невозможным получить прибор для механического раздражения кожи, чтобы его работа не сопровождалась какими-нибудь звуками, или достигнуть изменений высоты тона без одновременного изменения его силы. Ввиду этого я в одной из своих речей даже выразился так, что предстоит интересная конкуренция между животными анализаторами как инструментами и физическими приборами. Теперь наш фактический материал. Возьмем оптический анализатор собаки. С одной стороны, этот анализатор оказался чрезвычайным, далеко превосходящим человеческое зрение. Это имена отношении различения интенсивностей света. У собаки был образован условный рефлекс на появление перед ее глазами черного экрана, совершенно равномерно окрашенного, без малейших выделяющихся линий или точек. От него был отдифференцирован совершенно такой же формы и размера белый экран, также вполне равномерной окраски. Затем в распоряжении экспериментатора имелся комплекс постепенных оттенков (50 номеров циммермановской коллекции) от белого цвета через серые до черного. После белого постепенно отдифференцировывалисерые экраны, все более и более темные. Кончилось тем, что отчетливо отличался оптическим прибором собаки № 49 от 50, тогда как человеческий глаз не только при последовательном рассматривании этих оттенков через известные промежутки времени, но и при одновременном сравнении не мог уловить между ними никакой разницы. То же относилось и к нескольким более удаленным друг от друга оттенкам (опыты Фролова). Таким образом оптический анализатор собаки в отношении анализа интенсивности освещения идет чрезвычайно далеко, и мы не могли определить предел его деятельности в этом отношении.
Совершенно другое оказалось в отношении других сторон деятельности этого прибора: именно в отношении анализа цветов и форм.
Л. А. Орбели в первой своей работе совсем не мог констатировать различения цветов его собаками. Во второй работе это удалось, наконец, у одной собаки, да и то в не совсем безупречных опытах. Принимая во внимание результаты других исследователей, как наших, так и иностранных, приходится заключить, что цветное зрение у собак существует вообще в зачаточной форме и у многих собак совсем отсутствует.
Что касается дифференцирования форм, то мы располагаем следующими опытами (опыты Н. Р. Шенгер-Крестовниковой). У собаки был выработан условный пищевой рефлекс на светлый круг, который отбрасывался на стоящий перед собакой экран. Затем были предприняты дифференцировки от этого круга эллипсисов той же величины площади и того же освещения. Первый из примененных эллипсисов имел отношение полуосей 2 : 1. Дифференцировка произошла. Далее употребляемые эллипсисы постепенно приближались к кругу, и таким образом достигалось все более и более тонкое различение формы. При эллипсисе с отношением полуосей 9: 8 оказался предел, обнаружившийся в таком виде. Образовавшаяся было на него дифференцировка с повторением постепенно разрушалась, и это повлекло за собой исчезание и ранних, более грубых дифференцировок. Пришлось снова и осторожно вырабатывать дифференцировки с начала. Применение эллипсиса с отношением 9 : 8 теперь в первый раз дало нуль условнорефлекторного эффекта, а затем при следующих пробах с ним произошла старая история: не только не повторилась эта дифференцировка (если это была истинная дифференцировка), но исчезли и все ранние грубые. Этот факт ясно воспроизводит тот случаи, о котором упомянуто в конце предшествующей лекции. Анализ сам по себе на этой ступени еще возможен, но продолжительная постоянная утилизация его для соответствующей деятельности организма встречает неодолимое препятствие на почве отношений между раздражительным и тормощным процессами.
При наших опытах применялись также дифференцировки направлений движущихся фигур и точек, но при них предел отличения не изучался.
Анализаторная деятельность звукового нервного аппарата у собаки подверглась нами особенно подробному исследованию в очень различных направлениях. На первом месте и здесь надо поставить анализ интенсивностей одного и того же звука. Оказалось, что каждая интенсивность звука может быть легко сделана особым постоянным условным раздражителем, отдифференцированным как от высшей, так и от низшей интенсивности. К сожалению, эти опыты (опыты Н. П. Тихомирова) исполнены в довольно упрощенной форме. Звук (тон в 1740 колебаний в секунду) получался от органной трубы, надуваемой из газометра под постоянным давлением (3.6--3.88 см водяного столба). Органная труба укреплялась в центре деревянной доски, покрытой слоем ваты. Сверху над этой доской висел деревянный ящик, выложенный внутри ватой и открытый только внизу. Точным подниманием и опусканием этого ящика над трубой достигалось определенное заглушение звука. Целью опытов имелось определить предел различения интенсивностей звука собакой по сравнению с человеком. Уже предпоследняя из приближающихся к обычной, т. е. сделанной условным раздражителем интенсивностей, абсолютно различалась собакой на расстоянии 17 часов после применения обычной, в то время как экспериментатор мог уловить разницу этих звуков только при непосредственном следовании их одного за другим.
Таким образом оптический анализатор собаки в отношении анализа интенсивности освещения идет чрезвычайно далеко, и мы не могли определить предел его деятельности в этом отношении.
Совершенно другое оказалось в отношении других сторон деятельности этого прибора: именно в отношении анализа цветов и форм.
Л. А. Орбели в первой своей работе совсем не мог констатировать различения цветов его собаками. Во второй работе это удалось, наконец, у одной собаки, да и то в не совсем безупречных опытах. Принимая во внимание результаты других исследователей, как наших, так и иностранных, приходится заключить, что цветное зрение у собак существует вообще в зачаточной форме и у многих собак совсем отсутствует.
Что касается дифференцирования форм, то мы располагаем следующими опытами (опыты Н. Р. Шенгер-Крестовниковой). У собаки был выработан условный пищевой рефлекс на светлый круг, который отбрасывался на стоящий перед собакой экран. Затем были предприняты дифференцировки от этого круга эллипсисов той же величины площади и того же освещения. Первый из примененных эллипсисов имел отношение полуосей 2 : 1. Дифференцировка произошла. Далее употребляемые эллипсисы постепенно приближались к кругу, и таким образом достигалось все более и более тонкое различение формы. При эллипсисе с отношением полуосей 9: 8 оказался предел, обнаружившийся в таком виде. Образовавшаяся было на него дифференцировка с повторением постепенно разрушалась, и это повлекло за собой исчезание и ранних, более грубых дифференцировок. Пришлось снова и осторожно вырабатывать дифференцировки с начала. Применение эллипсиса с отношением 9 : 8 теперь в первый раз дало нуль условнорефлекторного эффекта, а затем при следующих пробах с ним произошла старая история: не только не повторилась эта дифференцировка (если это была истинная дифференцировка), но исчезли и все ранние грубые. Этот факт ясно воспроизводит тот случаи, о котором упомянуто в конце предшествующей лекции. Анализ сам по себе на этой ступени еще возможен, но продолжительная постоянная утилизация его для соответствующей деятельности организма встречает неодолимое препятствие на почве отношений между раздражительным и тормощным процессами.
При наших опытах применялись также дифференцировки направлений движущихся фигур и точек, но при них предел отличения не изучался.
Анализаторная деятельность звукового нервного аппарата у собаки подверглась нами особенно подробному исследованию в очень различных направлениях. На первом месте и здесь надо поставить анализ интенсивностей одного и того же звука. Оказалось, что каждая интенсивность звука может быть легко сделана особым постоянным условным раздражителем, отдифференцированным как от высшей, так и от низшей интенсивности. К сожалению, эти опыты (опыты Н. П. Тихомирова) исполнены в довольно упрощенной форме. Звук (тон в 1740 колебаний в секунду) получался от органной трубы, надуваемой из газометра под постоянным давлением (3.6--3.88 см водяного столба). Органная труба укреплялась в центре деревянной доски, покрытой слоем ваты. Сверху над этой доской висел деревянный ящик, выложенный внутри ватой и открытый только внизу. Точным подниманием и опусканием этого ящика над трубой достигалось определенное заглушение звука. Целью опытов имелось определить предел различения интенсивностей звука собакой по сравнению с человеком. Уже предпоследняя из приближающихся к обычной, т. е. сделанной условным раздражителем интенсивностей, абсолютно различалась собакой на расстоянии 17 часов после применения обычной, в то время как экспериментатор мог уловить разницу этих звуков только при непосредственном следовании их одного за другим.
 Но для собаки этот звук мог быть еще более приближен к условному раздражителю, причем даже на расстоянии часов от него дифференцировка была налицо. Однако эта дифференцировка в условиях нашей обстановки (комната обыкновенной лаборатории) очень легко нарушалась. Конечно, эти опыты должны быть повторены при помощи совершенных методов для получения абсолютных величин.
Затем много опытов было сделано с дифференцировкой высоты тона. Употреблялись главным образом разные духовые инструменты, причем предельная разница, нами испытанная и совершенно точно уловленная слуховым аппаратом собаки, была /s тона, как это приведено в предшествующей лекции. Дальше этого мы не шли, не уверенные в точности нашей методики. Повторение этих опытов с чистыми тонами не обнаружило ничего особенного сравнительно с обыкновенными сложными тонами (опыты Г. В. Анрепа и Т. М. Мануйлова).
Дальше мы исследовали предел раздражаемости слухового аппарата высокими тонами. Как при помощи гальтоновского свистка (опыты В. А. Бурмакина), так и при помощи прибора с чистыми тонами (опыты Л. A. Андреева) оказалось, что собака продолжает раздражаться такими высокими тонами, которых человек уже не слышит. И эта разница идет довольно далеко. Было интересно видеть, как собака резко и точно реагировала на звуки, которые для нас не существовали.
Затем были испытаны дифференцировки тембра тонов и места звуков, но на тонкости этих дифференцировок мы не настаивали.
В этом же ряду опытов можно поставить испытание звуков, где суть дела была, однако, не в самых звуках, а в промежутках времени между этими звуками, именно - исследование дифференцирования различной частоты ударов метронома. ДифФеренцировки на различную частоту ударов метронома образуются очень легко. Ho представляет интерес предел такой дифференцировки. Этот предел оказался очень тонким, недоступным для человека, - именно отличаются очень точно и при больших промежутках времени 100 ударов в минуту от 96.
Меньшее число испытаний пришлось в наших работах на кожные анализаторы, как на механический, так и термическии. Прежде всего испытывалась дифференцировка по месту, локализация. Она, конечно, оказалась, но не исследовалась на ее тонкость, ее предел. Затем отдифференцировывались различные формы механического раздражения: чесание, надавливание гладкой и шероховатой поверхностью, покалывание притупленными штифтиками при их различном расположении, чесание в различных направлениях. Так же определялись дифференцировки градусов температуры.
Хотя химический носовой прибор собаки является совершеннейшим из ее анализаторов, однако исследование его у нас остается и до сих пор самым скудным, конечно по методическим основаниям. С запахами трудно или невозможно, по крайней мере сейчас, так точно и определенно оперировать, как и со всякими другими раздражителями. Нельзя точно ограничить их действие во времени, и нет ни субъективного, ни объективного критерия их наличности или отсутствия, что касается их следов. Поэтому мы ограничились немногими опытами. Отличались друг от друга разные запахи: камфоры, ванилина и т. д. Из одних делались пищевые условные раздражители, из других кислотные; из одних - положительные условные раздражители, из других - тормозные. Испытывался анализ смеси запахов, когда к ней прибавлялся новый запах.
Наконец мы собрали некоторый материал относительно ротового химического анализатора. Тут дело принимает у нас несколько иной вид, так как оба наших обычных безусловных рефлекса относятся именно к вкусовому анализатору. Чтобы быть в одинаковом положении с изучением других анализаторов, следовало бы иметь безусловный раздражитель, принадлежащий к тому или другому из этих анализаторов, а химические раздражения полости рта применять в качестве условных. Мы таких опытов не делали, а заменили их до известной степени другой формой опытов: взаимодействием, задерживанием одного другим, условных рефлексов, образованных на разные пищевые и отвергаемые вещества (мясной порошок, сухарный порошок, сахар, сыр, кислота, сода и т. д.). Вот пример таких опытов из работы Я. И. Егорова.
Механическое раздражение кожи условный раздражитель, связанный с едой мясо-сухарного порошка; вертящийся предмет - условный раздражитель, связанный с едой голландского сыра.
Но для собаки этот звук мог быть еще более приближен к условному раздражителю, причем даже на расстоянии часов от него дифференцировка была налицо. Однако эта дифференцировка в условиях нашей обстановки (комната обыкновенной лаборатории) очень легко нарушалась. Конечно, эти опыты должны быть повторены при помощи совершенных методов для получения абсолютных величин.
Затем много опытов было сделано с дифференцировкой высоты тона. Употреблялись главным образом разные духовые инструменты, причем предельная разница, нами испытанная и совершенно точно уловленная слуховым аппаратом собаки, была /s тона, как это приведено в предшествующей лекции. Дальше этого мы не шли, не уверенные в точности нашей методики. Повторение этих опытов с чистыми тонами не обнаружило ничего особенного сравнительно с обыкновенными сложными тонами (опыты Г. В. Анрепа и Т. М. Мануйлова).
Дальше мы исследовали предел раздражаемости слухового аппарата высокими тонами. Как при помощи гальтоновского свистка (опыты В. А. Бурмакина), так и при помощи прибора с чистыми тонами (опыты Л. A. Андреева) оказалось, что собака продолжает раздражаться такими высокими тонами, которых человек уже не слышит. И эта разница идет довольно далеко. Было интересно видеть, как собака резко и точно реагировала на звуки, которые для нас не существовали.
Затем были испытаны дифференцировки тембра тонов и места звуков, но на тонкости этих дифференцировок мы не настаивали.
В этом же ряду опытов можно поставить испытание звуков, где суть дела была, однако, не в самых звуках, а в промежутках времени между этими звуками, именно - исследование дифференцирования различной частоты ударов метронома. ДифФеренцировки на различную частоту ударов метронома образуются очень легко. Ho представляет интерес предел такой дифференцировки. Этот предел оказался очень тонким, недоступным для человека, - именно отличаются очень точно и при больших промежутках времени 100 ударов в минуту от 96.
Меньшее число испытаний пришлось в наших работах на кожные анализаторы, как на механический, так и термическии. Прежде всего испытывалась дифференцировка по месту, локализация. Она, конечно, оказалась, но не исследовалась на ее тонкость, ее предел. Затем отдифференцировывались различные формы механического раздражения: чесание, надавливание гладкой и шероховатой поверхностью, покалывание притупленными штифтиками при их различном расположении, чесание в различных направлениях. Так же определялись дифференцировки градусов температуры.
Хотя химический носовой прибор собаки является совершеннейшим из ее анализаторов, однако исследование его у нас остается и до сих пор самым скудным, конечно по методическим основаниям. С запахами трудно или невозможно, по крайней мере сейчас, так точно и определенно оперировать, как и со всякими другими раздражителями. Нельзя точно ограничить их действие во времени, и нет ни субъективного, ни объективного критерия их наличности или отсутствия, что касается их следов. Поэтому мы ограничились немногими опытами. Отличались друг от друга разные запахи: камфоры, ванилина и т. д. Из одних делались пищевые условные раздражители, из других кислотные; из одних - положительные условные раздражители, из других - тормозные. Испытывался анализ смеси запахов, когда к ней прибавлялся новый запах.
Наконец мы собрали некоторый материал относительно ротового химического анализатора. Тут дело принимает у нас несколько иной вид, так как оба наших обычных безусловных рефлекса относятся именно к вкусовому анализатору. Чтобы быть в одинаковом положении с изучением других анализаторов, следовало бы иметь безусловный раздражитель, принадлежащий к тому или другому из этих анализаторов, а химические раздражения полости рта применять в качестве условных. Мы таких опытов не делали, а заменили их до известной степени другой формой опытов: взаимодействием, задерживанием одного другим, условных рефлексов, образованных на разные пищевые и отвергаемые вещества (мясной порошок, сухарный порошок, сахар, сыр, кислота, сода и т. д.). Вот пример таких опытов из работы Я. И. Егорова.
Механическое раздражение кожи условный раздражитель, связанный с едой мясо-сухарного порошка; вертящийся предмет - условный раздражитель, связанный с едой голландского сыра.
 Го же самое получалось, когда применялись одни условные раздражители, без участия безусловных.
О пыт из той же работы. Механическое раздражение кожи давало перед этим опытом обычно 5-6 капель за 30 секунд.
Го же самое получалось, когда применялись одни условные раздражители, без участия безусловных.
О пыт из той же работы. Механическое раздражение кожи давало перед этим опытом обычно 5-6 капель за 30 секунд.
 В этом ряде опытов отчетливо выступила анализаторная деятельность мозговых концов химического анализатора. У одной собаки при обычной еде, состоящей из овсяной каши с мясом и хлебом, были выработаны отдельные условные рефлексы на мясной порошок и сахарный песок. Затем мясо и хлеб были исключены из еды, а к каше было прибавлено большое количество сахара. После некоторого срока такоео пищевого режима условный рефлекс на мясной порошок значительно увеличился. а рефлекс на сахар почти исчез (опыты А. А. Савича).
В других опытах (С. Б. Хазена) то же и более подробно проделано с отвергаемыми веществами. Из прежних наших опытов было известно, что при условных рефлексах на кислоту в течение каждого опытного сеанса обыкновенно величина как условного, так и безусловного рефлексов концу постепенно растет. Точно так же и в ряде опытов изо дня в день эта величина также поднимается до известного максимума. В работе Хазена прежде всего было введено такое изменение, что после первого применения условного раздражителя и подкрепления его несколько раз кислота вливалась без условного раздражителя, а в конце снова испытывался условный раздражитель. Теперь он всегда оказывался увеличенным сравнительно с первой пробой. Затем ежедневные опыты прерывались тремя промежутками, в одном ряду опытов в 5 дней, в другом в 3 дня. В один промежуток у одной собаки в rectum, у другой зондом в желудок вливалось значительное количество раствора кислоты, в другой раз - раствора соды и в третий не делалось ни того, ни другого. После каждого такого промежутка испытывались условный и безусловный рефлексы. После промежутка с вливанием кислоты величина рефлексов или остается такой же, или падает немного сравнительно с опытами перед промежутком, после промежутка без вливаний она значительно падает, а после вливания соды падает еще дальше. Вот цифры из опыта над одной собакой. При ежедневных опытах из подчелюстной слюнной железы на определенное количество вливаемой в рот кислоты выделяется в среднем слюны 5.1 куб. см и на условный раздражитель получается 4 капли. При перерыве без вливания - соответственно 3.8 куб. см и 2 капли. При перерыве с вливанием соды -----апель. При перерыве с вливанием кислоты - 4.5 куб. см и 3 капли. Таким образом тот или другой химический состав крови различался мозговым концом химического анализатора, выражаясь то в повышении, то в понижении возбудимости соответствующей части этого анализатора. Когда в кровь избыточно поступала кислота, повышалась возбудимость кислотной части химического анализатора и организм при встрече с кислотой из внешнего мира энергичными отрицательными двигательным и секреторным рефлексами исключал в большей или меньшей мере дальнейшее ее поступление. То же самое, конечно, происходит и с пищевыми веществами, выражаясь соответственно в усилении и ослаблении то положительной, то отрицательной реакции на те или другие вещества и в тех или других количествах их. Итак, химический ротовой анализатор своими концами соединяет две среды: внутреннюю среду организма и внешнюю, регулируя их соотношение и тем обеспечивая нормальный состав организма.
Последние, только что описанные опыты (не повторенные, к сожалению, потом) относятся более раннему периоду нашей работы, когда нас, при новизне и сложности дела, сторожили многие источники ошибок.
Механизм и локализация синтетической нервной деятельности, в противоположность аналитической, остаются до сих пор мало известными. Проще всего представлять их себе в виде соединений между нервными клетками, будут ли это разделительные мембраны или просто утончающиеся разветвления с принадлежащими им свойствами. Конечно, сейчас первая задача накоплять факты, относящиеся к синтетической деятельности.
Подробности относительно синтетической деятельности полушарий заняли сравнительно мало места в наших исследованиях. Помимо образования условного рефлекса, который прежде всего есть акт синтеза и которым мы постоянно пользовались как исходным фактом нашей работы, мы несколько остановились на комплексных, сложных условных раздражителям.
Мы применяли в качестве условных раздражителей как одновременные, так и последовательные комплексы различных раздражителей.
При одновременных комплексах наблюдались следующие важные отношения.
Если берутся два комплекса раздтносящиеся к разным анализаторам, то одно раздражение почти или совер- ершенно маскирует условное действие другого при отдельной их пробе, как бы долго ни укреплялся рефлекс на комплексный раздражитель. Механическое раздражение кожи заслоняет температурное ее раздражение, звуковое - световое.
О п ы т А. В. Палладина. Образован условный кислотный рефлекс на одновременное охлаждение (температурой тающего льда) и механическое раздражение кожи. Теперь пробуются как комплексный раздражитель, так и отдельные.
В этом ряде опытов отчетливо выступила анализаторная деятельность мозговых концов химического анализатора. У одной собаки при обычной еде, состоящей из овсяной каши с мясом и хлебом, были выработаны отдельные условные рефлексы на мясной порошок и сахарный песок. Затем мясо и хлеб были исключены из еды, а к каше было прибавлено большое количество сахара. После некоторого срока такоео пищевого режима условный рефлекс на мясной порошок значительно увеличился. а рефлекс на сахар почти исчез (опыты А. А. Савича).
В других опытах (С. Б. Хазена) то же и более подробно проделано с отвергаемыми веществами. Из прежних наших опытов было известно, что при условных рефлексах на кислоту в течение каждого опытного сеанса обыкновенно величина как условного, так и безусловного рефлексов концу постепенно растет. Точно так же и в ряде опытов изо дня в день эта величина также поднимается до известного максимума. В работе Хазена прежде всего было введено такое изменение, что после первого применения условного раздражителя и подкрепления его несколько раз кислота вливалась без условного раздражителя, а в конце снова испытывался условный раздражитель. Теперь он всегда оказывался увеличенным сравнительно с первой пробой. Затем ежедневные опыты прерывались тремя промежутками, в одном ряду опытов в 5 дней, в другом в 3 дня. В один промежуток у одной собаки в rectum, у другой зондом в желудок вливалось значительное количество раствора кислоты, в другой раз - раствора соды и в третий не делалось ни того, ни другого. После каждого такого промежутка испытывались условный и безусловный рефлексы. После промежутка с вливанием кислоты величина рефлексов или остается такой же, или падает немного сравнительно с опытами перед промежутком, после промежутка без вливаний она значительно падает, а после вливания соды падает еще дальше. Вот цифры из опыта над одной собакой. При ежедневных опытах из подчелюстной слюнной железы на определенное количество вливаемой в рот кислоты выделяется в среднем слюны 5.1 куб. см и на условный раздражитель получается 4 капли. При перерыве без вливания - соответственно 3.8 куб. см и 2 капли. При перерыве с вливанием соды -----апель. При перерыве с вливанием кислоты - 4.5 куб. см и 3 капли. Таким образом тот или другой химический состав крови различался мозговым концом химического анализатора, выражаясь то в повышении, то в понижении возбудимости соответствующей части этого анализатора. Когда в кровь избыточно поступала кислота, повышалась возбудимость кислотной части химического анализатора и организм при встрече с кислотой из внешнего мира энергичными отрицательными двигательным и секреторным рефлексами исключал в большей или меньшей мере дальнейшее ее поступление. То же самое, конечно, происходит и с пищевыми веществами, выражаясь соответственно в усилении и ослаблении то положительной, то отрицательной реакции на те или другие вещества и в тех или других количествах их. Итак, химический ротовой анализатор своими концами соединяет две среды: внутреннюю среду организма и внешнюю, регулируя их соотношение и тем обеспечивая нормальный состав организма.
Последние, только что описанные опыты (не повторенные, к сожалению, потом) относятся более раннему периоду нашей работы, когда нас, при новизне и сложности дела, сторожили многие источники ошибок.
Механизм и локализация синтетической нервной деятельности, в противоположность аналитической, остаются до сих пор мало известными. Проще всего представлять их себе в виде соединений между нервными клетками, будут ли это разделительные мембраны или просто утончающиеся разветвления с принадлежащими им свойствами. Конечно, сейчас первая задача накоплять факты, относящиеся к синтетической деятельности.
Подробности относительно синтетической деятельности полушарий заняли сравнительно мало места в наших исследованиях. Помимо образования условного рефлекса, который прежде всего есть акт синтеза и которым мы постоянно пользовались как исходным фактом нашей работы, мы несколько остановились на комплексных, сложных условных раздражителям.
Мы применяли в качестве условных раздражителей как одновременные, так и последовательные комплексы различных раздражителей.
При одновременных комплексах наблюдались следующие важные отношения.
Если берутся два комплекса раздтносящиеся к разным анализаторам, то одно раздражение почти или совер- ершенно маскирует условное действие другого при отдельной их пробе, как бы долго ни укреплялся рефлекс на комплексный раздражитель. Механическое раздражение кожи заслоняет температурное ее раздражение, звуковое - световое.
О п ы т А. В. Палладина. Образован условный кислотный рефлекс на одновременное охлаждение (температурой тающего льда) и механическое раздражение кожи. Теперь пробуются как комплексный раздражитель, так и отдельные.
 О п ы т Зеленого. Образован условный пищевой рефлекс на одновременное действие тона духового камертона (очень заглушенного помещением в деревянном ящике, обитом ватой) и 3 электрических лампочек (по 16 свечей каждая), помещенных перед мордой собаки в несколько затемненной комнате.
О п ы т Зеленого. Образован условный пищевой рефлекс на одновременное действие тона духового камертона (очень заглушенного помещением в деревянном ящике, обитом ватой) и 3 электрических лампочек (по 16 свечей каждая), помещенных перед мордой собаки в несколько затемненной комнате.
 Конечно, здесь замаскированные раздражители, примененные отдельно для образования условных рефлексов, могут обладать значительными эффектами.
Как понимать только что описанный факт? Опыты с комплексным раздражителем из разных агентов, принадлежащих к одному и тому же анализатору, дают основание одному из вероятных предположений. Как комплексный раздражитель брались два звука, по слуху приблизительно одинаковой интенсивности. Когда условный рефлекс образовался, испытывалось в отдельности действие каждого из звуков. Они имели равные эффекты. При образовании рефлекса из комплекса звуков резко разной интенсивности отдельное действие слабого звука или было очень мало, или совершенно отсутствовало.
О п ы т ы Зеленого. У собаки выработан комплексный пищевой рефлекс на сильный звук свистка и тон disi духового камертона приблизительно по слуху одинаковой силы. При отдельной пробе оба эти раздражителя дали за минуту по 19 капель. У нее же был, кроме того, образован другой комплексный раздражитель из того же звука свистка, но с более слабым тоном а1 духового камертона. В отдельности свист за 30 секунд дал 7 капель, тон --- 1.
В этих опытах очевидно, что замаскирования одного раздражителя другим определяется различной силой раздражителей. То же естественно предположить и в случае раздражителей, принадлежащих к разным анализаторам. По этому предположению, кожно-механическое раздражение при наших опытах было бы всегда сильнее кожно-температурного, звуковое -сильнее светового. Это важное предположение должно быть проверено применением комплексных раздражителей, составленных из раздражителей, принадлежащих к разным анализаторам, при наибольшем разнообразии интенсивностей раздражителей, например комбинированием слабейшего звукового раздражителя с сильнейшим световым.
Факт замаскирования в одновременном комплексном (из разных анализаторов) раздражителе одного раздражителя другим заключает в себе несколько интересных подробностей.
Часто величина комплексного эффектавняется величине эффекта отдельно примененного сильного компонента, т. е. слабое раздражение как бы вполне покрывается, уничтожается сильным. Между тем, если сильный раздражитель повторять отдельно, хотя бы изредка, без подкрепления безусловным раздражителем, постоянно подкрепляя комплексный, то сильный один совершенно теряет свое действие, а комплексный сохраняет свой обычный размер. Следовательно, слабый раздражитель все же, хотя и латентно, участвует в комплексном раздражителе (опыты Палладина).
Другой факт (из работы Перельцвейга) был приведен в четвертой лекции. Если слабый или даже отдельно совсем недействующий раздражитель из комплекса повторять через короткие промежутки (минуты) без подкрепления безусловным раздражителем, т. е. угашать, то вторично угашаются в известной степени как сильный компонент, так даже и комплексный. Следовательно, и в этом опыте как бы не существующий компонент, вследствие процедуры угасания, переходит в агент, производящий явное тормозное действие.
В единственном доселе сделанном нами, правда безупречном, опыте наблюдалось дальше следующее положение дела. Если из агентов, относящихся к разным анализаторам, сделать сперва отдельные условные раздражители, затем применить их вместе, т. е. сделать из них комплексный раздражитель, то замаскирования одних раздражителей другими почти не происходит, несмотря на большое число повторений комплексного раздражителя. Из этого можно заключить, что в комплексном раздражителе, образуемом впервые из индифферентных агентов, сильный агент сейчас же как-то не позволяет слабому образовать соответственно сильную связь с центром безусловного рефлекса.
Механизм замаскирования, который, конечно, надо понимать как торможение, будет специально анализирован в одной из последующих лекций, посвящаемой главным образом общему обзору нашего материала.
Уже в приведенных случаях замаскирования очевидно взаимодействие разных клеток коры друг на друга, слитие, синтезирование происходящих в них процессов, при их одновременном раздражении. При одновременном комплексе раздражителей, принадлежащих к одному и тому же анализатору, и притом равной интенсивности, это синтезирование как будто не бросается в глаза. Однако почему и в этих случаях нет суммации, а эффект отдельного сильного компонента равен эффекту комплексного раздражителя? Но при следующей вариации опыта при новом условии факт синтезирования и в этом случае, т. е. при раздражителях из одного и того же анализатора, становится в высшей степени резок. Впервые в опытах Зеленого оказалось, а затем в опытах Мануйлова и Крылова повторилось и сделалось заурядным явлением следующее положение вещей. Мы имеем только что упомянутый комплексный раздражитель. Не составляет труда достигнуть того, чтобы его действие продолжалось, а действие его компонентов в отдельности исчезло, и эти ранее положительные условные раздражители превратились в отрицательные, тормозные. Достигается это, конечно, постоянным подкреплением комплексного раздражителя и повторением компонентов без сопровождения безусловным рефлексом. Опыт идет и в обратном виде. Можно таким же способом сделать комплекс тормозным, а компоненты положительными.
Не останавливаясь пока на оценке этого факта, перехожу к последовательным комплексным раздражителям, где то же явление выступает, так сказать, в еще более утонченной форме. Эти комплексные раздражители были у нас очень разнообразными. То комплекс состоял из одного и того же раздражителя (определенного тона), который продолжался секунду и повторялся три раза, причем между первым звуком и вторым была пауза в 2 секунды, а между вторым и третьим секунда, и затем эта группа через промежуток в 5 секунд повторялась и сопровождалась безусловным рефлексом. То в других опытах комплекс состоял из 3-4 раздражителей разных, но принадлежащих к одному и тому же анализатору и следующих в определенном порядке друг за другом при тех же продолжительности и паузах, например 4 тона: до, ре, ми, фа, или шум, два разных тона и звонок. Наконец комплекс составляли 3- 4 раздражителя, принадлежащие к разным анализаторам той же продолжительности и при тех же паузах. Из всех этих комплексов рефлексы образовывались легко, и после известного практикования рефлексов все компоненты их также обнаруживали положительное условное действие соответственно их силе и роду.
Затем мы приступали к изменению этих комплексов. В первом случае переставляли паузу в комплексе: длинную вставляли между вторым и третьим звуками. В остальных изменяли порядок отдельных раздражителей то целиком, превращая его в обратный, то, в случае четырех раздражителей, переставляя местами только два внутренних. И такие измененные комплексы повторно применяли без сопровождения безусловным рефлексом, постоянно подкрепляя первоначальные. нчательном результате происходило дифференцирование первоначальных комплексов от измененных, последние теряли их условное положительное действие, становясь отрицательными, тормозными (опыты Б. П. Бабкина, В. В. Строганова, Л. С. Григорович, А. Г. Иванова-Смоленского и М. Н. Юрман).
О п ы т Юрман. Положительный пищевой условный раздражитель состоял из вспыхивания электрической лампочки (С), механического раздражителя кожи (К) и звука бульканья (3); тормозной - из 3 К С.
Конечно, здесь замаскированные раздражители, примененные отдельно для образования условных рефлексов, могут обладать значительными эффектами.
Как понимать только что описанный факт? Опыты с комплексным раздражителем из разных агентов, принадлежащих к одному и тому же анализатору, дают основание одному из вероятных предположений. Как комплексный раздражитель брались два звука, по слуху приблизительно одинаковой интенсивности. Когда условный рефлекс образовался, испытывалось в отдельности действие каждого из звуков. Они имели равные эффекты. При образовании рефлекса из комплекса звуков резко разной интенсивности отдельное действие слабого звука или было очень мало, или совершенно отсутствовало.
О п ы т ы Зеленого. У собаки выработан комплексный пищевой рефлекс на сильный звук свистка и тон disi духового камертона приблизительно по слуху одинаковой силы. При отдельной пробе оба эти раздражителя дали за минуту по 19 капель. У нее же был, кроме того, образован другой комплексный раздражитель из того же звука свистка, но с более слабым тоном а1 духового камертона. В отдельности свист за 30 секунд дал 7 капель, тон --- 1.
В этих опытах очевидно, что замаскирования одного раздражителя другим определяется различной силой раздражителей. То же естественно предположить и в случае раздражителей, принадлежащих к разным анализаторам. По этому предположению, кожно-механическое раздражение при наших опытах было бы всегда сильнее кожно-температурного, звуковое -сильнее светового. Это важное предположение должно быть проверено применением комплексных раздражителей, составленных из раздражителей, принадлежащих к разным анализаторам, при наибольшем разнообразии интенсивностей раздражителей, например комбинированием слабейшего звукового раздражителя с сильнейшим световым.
Факт замаскирования в одновременном комплексном (из разных анализаторов) раздражителе одного раздражителя другим заключает в себе несколько интересных подробностей.
Часто величина комплексного эффектавняется величине эффекта отдельно примененного сильного компонента, т. е. слабое раздражение как бы вполне покрывается, уничтожается сильным. Между тем, если сильный раздражитель повторять отдельно, хотя бы изредка, без подкрепления безусловным раздражителем, постоянно подкрепляя комплексный, то сильный один совершенно теряет свое действие, а комплексный сохраняет свой обычный размер. Следовательно, слабый раздражитель все же, хотя и латентно, участвует в комплексном раздражителе (опыты Палладина).
Другой факт (из работы Перельцвейга) был приведен в четвертой лекции. Если слабый или даже отдельно совсем недействующий раздражитель из комплекса повторять через короткие промежутки (минуты) без подкрепления безусловным раздражителем, т. е. угашать, то вторично угашаются в известной степени как сильный компонент, так даже и комплексный. Следовательно, и в этом опыте как бы не существующий компонент, вследствие процедуры угасания, переходит в агент, производящий явное тормозное действие.
В единственном доселе сделанном нами, правда безупречном, опыте наблюдалось дальше следующее положение дела. Если из агентов, относящихся к разным анализаторам, сделать сперва отдельные условные раздражители, затем применить их вместе, т. е. сделать из них комплексный раздражитель, то замаскирования одних раздражителей другими почти не происходит, несмотря на большое число повторений комплексного раздражителя. Из этого можно заключить, что в комплексном раздражителе, образуемом впервые из индифферентных агентов, сильный агент сейчас же как-то не позволяет слабому образовать соответственно сильную связь с центром безусловного рефлекса.
Механизм замаскирования, который, конечно, надо понимать как торможение, будет специально анализирован в одной из последующих лекций, посвящаемой главным образом общему обзору нашего материала.
Уже в приведенных случаях замаскирования очевидно взаимодействие разных клеток коры друг на друга, слитие, синтезирование происходящих в них процессов, при их одновременном раздражении. При одновременном комплексе раздражителей, принадлежащих к одному и тому же анализатору, и притом равной интенсивности, это синтезирование как будто не бросается в глаза. Однако почему и в этих случаях нет суммации, а эффект отдельного сильного компонента равен эффекту комплексного раздражителя? Но при следующей вариации опыта при новом условии факт синтезирования и в этом случае, т. е. при раздражителях из одного и того же анализатора, становится в высшей степени резок. Впервые в опытах Зеленого оказалось, а затем в опытах Мануйлова и Крылова повторилось и сделалось заурядным явлением следующее положение вещей. Мы имеем только что упомянутый комплексный раздражитель. Не составляет труда достигнуть того, чтобы его действие продолжалось, а действие его компонентов в отдельности исчезло, и эти ранее положительные условные раздражители превратились в отрицательные, тормозные. Достигается это, конечно, постоянным подкреплением комплексного раздражителя и повторением компонентов без сопровождения безусловным рефлексом. Опыт идет и в обратном виде. Можно таким же способом сделать комплекс тормозным, а компоненты положительными.
Не останавливаясь пока на оценке этого факта, перехожу к последовательным комплексным раздражителям, где то же явление выступает, так сказать, в еще более утонченной форме. Эти комплексные раздражители были у нас очень разнообразными. То комплекс состоял из одного и того же раздражителя (определенного тона), который продолжался секунду и повторялся три раза, причем между первым звуком и вторым была пауза в 2 секунды, а между вторым и третьим секунда, и затем эта группа через промежуток в 5 секунд повторялась и сопровождалась безусловным рефлексом. То в других опытах комплекс состоял из 3-4 раздражителей разных, но принадлежащих к одному и тому же анализатору и следующих в определенном порядке друг за другом при тех же продолжительности и паузах, например 4 тона: до, ре, ми, фа, или шум, два разных тона и звонок. Наконец комплекс составляли 3- 4 раздражителя, принадлежащие к разным анализаторам той же продолжительности и при тех же паузах. Из всех этих комплексов рефлексы образовывались легко, и после известного практикования рефлексов все компоненты их также обнаруживали положительное условное действие соответственно их силе и роду.
Затем мы приступали к изменению этих комплексов. В первом случае переставляли паузу в комплексе: длинную вставляли между вторым и третьим звуками. В остальных изменяли порядок отдельных раздражителей то целиком, превращая его в обратный, то, в случае четырех раздражителей, переставляя местами только два внутренних. И такие измененные комплексы повторно применяли без сопровождения безусловным рефлексом, постоянно подкрепляя первоначальные. нчательном результате происходило дифференцирование первоначальных комплексов от измененных, последние теряли их условное положительное действие, становясь отрицательными, тормозными (опыты Б. П. Бабкина, В. В. Строганова, Л. С. Григорович, А. Г. Иванова-Смоленского и М. Н. Юрман).
О п ы т Юрман. Положительный пищевой условный раздражитель состоял из вспыхивания электрической лампочки (С), механического раздражителя кожи (К) и звука бульканья (3); тормозной - из 3 К С.
 О п ы т Иванова-Смоленского. Положительный пищевой условный раздражитель состоял из шипения (Ш), высокого тона (вТ), низкого тона (нТ) и звон тормозной из Ш н вТ З.
О п ы т Иванова-Смоленского. Положительный пищевой условный раздражитель состоял из шипения (Ш), высокого тона (вТ), низкого тона (нТ) и звон тормозной из Ш н вТ З.
 Образование тормозных рефлексов в этих случаях, в особенности в некоторых из них, чрезвычайно затягивалось. Если иногда и обнаруживалось довольно рано относительное дифференцирование, то полная, абсолютная дифференцировка достигалась в крайнем случае через сотни повторение. Приходилось даже к окончательному решению задачи итти иногда через этапы дифференцирования более простых комплексных раздражителей. Особенно трудно далась дифференцировка комплекса: шум, высокий тон, низкий тон и звонок, в котором местами были переставлены тоны. Все эти дифференцировки, и особенно трудные из них, оказывались очень нестойкими. С одной стороны, они очень страдали от частых повторений, с другой - слабели или исчезали временно от больших перерывов в работе. После достижения полного дифференцированиспытанные отдельно компоненты комплексов теряли свое условное положительное действие.
Как понимать эти факты? Каким образом комплекс из одних и тех же раздражителей, действующих, следовательно, на одни и те же клетки больших полушарий, мог хронически в один и тот же период времени оказываться различным раздражителем, вызывающим в этих клетках то раздражительный процесс, то тормозной? Где могло лежать основание для его различного действия? Как показывает факт, основанием его могло быть только синтезирование деятельности раздражаемых клеток. Клетки при данном условии должны были связаться, образовать сложную единицу, как это мы видим на постоянном факте образования условных рефлексов. При этом необходимо происходит влияние раздражаемых клеток друг на друга, их взаимодействие, как это ярко обнаружилось на одновременном комплексе раздражителей. При последовательном комплексе взаимодействие должно быть сложнее. Каждая клетка в нем иначе влияет на раздражаемую после нее клетку, смотря по тому, как на нее самоё влияла раздражаемая перед нею клетка. Следовательно, порядок одних и тех же раздражителей и паузы между ними должны быть факторами, определяющими окончательный результат раздражения, должны давать количественный (а может быть, и качественный) тот или другой итог всего комплексного раздражителя. мы знаем уже, что различные интенсивности одного и того же раздражения могут быть очень тонко дифференцированы, т. е. быть связанными одни с раздражительным процессом, другие с тормощным.
Из всего сообщенного следует, что надо различать элементарный анализ и также элементарный синтез от высшего анализа и синтеза. Если первые (специально анализ) свое первое основание имеют в свойствах и деятельности периферических концов анализаторов, то высший синтез и анализ осуществляются благодаря главным образом свойствам и деятельности центральных концов анализаторов.
При помощи условных рефлексов можно и должно производить подробнейшее экспериментальное исследование на животных как периферических, так и центральных концов анализаторов. Мы можем в доказательство широты этого экспериментырования на животных привести следующие наши примеры.
Одна задача состояла в том, чтобы посредством условных рефлексов получить экспериментальные данные относительно резонаторной теории Гельмгольца: нельзя ли путем частичного разрушения кортиева органа и membrana basilaris достигнуть выладения определенных тонов. Андреевым пока сделан один опыт с этой целью, и исследование продолжается. Вот этот опыт.
Применялись чистые тоны, доставляемые двумя приборами: одним от 100 до 3000, другим от 3000 до 26 000 колебаний в секунду. У собаки были выработаны разнообразные пищевые условные рефлексы: на кожно-механическое раздражение, световое и различные звуковые (звонок, удары метронома, шум и многочисленные чистые тоны). Сперва была вполне разрушена улитка с одной стороны. На шестой день после операции при перзом испытании все звуковые условные рефлексы были налицо. Затем была произведена вторая операция (10 III 1923) с другой :тороны с расчетом выключить только нижнюю часть тоновой шкалы. Костная часть улитки была разрушена бором на границе ге средней и нижней трети, перепончатая часть и кортиев орган были травматизированы тонкой иглой. Уже на десятый день после операции все звуковые раздражители действовали, за исключением тона в 600 колебаний в секунду и ниже. Но затем в течение 3 месяцев после операции постепенно восстановлялось действие и тонов ниже 600, остановившись на 300 с небольшим. Многочисленные пробы за остальную часть времени до года после операции установили верхнюю границу исчезнувших тонов между 309 и 317 колебаниями в секунду. Нижняя не могла быть определена за неимением у нас чистых тонов ниже 100 колебаний в секунду.
Вот два протокола из последнего периода испытаний.
Образование тормозных рефлексов в этих случаях, в особенности в некоторых из них, чрезвычайно затягивалось. Если иногда и обнаруживалось довольно рано относительное дифференцирование, то полная, абсолютная дифференцировка достигалась в крайнем случае через сотни повторение. Приходилось даже к окончательному решению задачи итти иногда через этапы дифференцирования более простых комплексных раздражителей. Особенно трудно далась дифференцировка комплекса: шум, высокий тон, низкий тон и звонок, в котором местами были переставлены тоны. Все эти дифференцировки, и особенно трудные из них, оказывались очень нестойкими. С одной стороны, они очень страдали от частых повторений, с другой - слабели или исчезали временно от больших перерывов в работе. После достижения полного дифференцированиспытанные отдельно компоненты комплексов теряли свое условное положительное действие.
Как понимать эти факты? Каким образом комплекс из одних и тех же раздражителей, действующих, следовательно, на одни и те же клетки больших полушарий, мог хронически в один и тот же период времени оказываться различным раздражителем, вызывающим в этих клетках то раздражительный процесс, то тормозной? Где могло лежать основание для его различного действия? Как показывает факт, основанием его могло быть только синтезирование деятельности раздражаемых клеток. Клетки при данном условии должны были связаться, образовать сложную единицу, как это мы видим на постоянном факте образования условных рефлексов. При этом необходимо происходит влияние раздражаемых клеток друг на друга, их взаимодействие, как это ярко обнаружилось на одновременном комплексе раздражителей. При последовательном комплексе взаимодействие должно быть сложнее. Каждая клетка в нем иначе влияет на раздражаемую после нее клетку, смотря по тому, как на нее самоё влияла раздражаемая перед нею клетка. Следовательно, порядок одних и тех же раздражителей и паузы между ними должны быть факторами, определяющими окончательный результат раздражения, должны давать количественный (а может быть, и качественный) тот или другой итог всего комплексного раздражителя. мы знаем уже, что различные интенсивности одного и того же раздражения могут быть очень тонко дифференцированы, т. е. быть связанными одни с раздражительным процессом, другие с тормощным.
Из всего сообщенного следует, что надо различать элементарный анализ и также элементарный синтез от высшего анализа и синтеза. Если первые (специально анализ) свое первое основание имеют в свойствах и деятельности периферических концов анализаторов, то высший синтез и анализ осуществляются благодаря главным образом свойствам и деятельности центральных концов анализаторов.
При помощи условных рефлексов можно и должно производить подробнейшее экспериментальное исследование на животных как периферических, так и центральных концов анализаторов. Мы можем в доказательство широты этого экспериментырования на животных привести следующие наши примеры.
Одна задача состояла в том, чтобы посредством условных рефлексов получить экспериментальные данные относительно резонаторной теории Гельмгольца: нельзя ли путем частичного разрушения кортиева органа и membrana basilaris достигнуть выладения определенных тонов. Андреевым пока сделан один опыт с этой целью, и исследование продолжается. Вот этот опыт.
Применялись чистые тоны, доставляемые двумя приборами: одним от 100 до 3000, другим от 3000 до 26 000 колебаний в секунду. У собаки были выработаны разнообразные пищевые условные рефлексы: на кожно-механическое раздражение, световое и различные звуковые (звонок, удары метронома, шум и многочисленные чистые тоны). Сперва была вполне разрушена улитка с одной стороны. На шестой день после операции при перзом испытании все звуковые условные рефлексы были налицо. Затем была произведена вторая операция (10 III 1923) с другой :тороны с расчетом выключить только нижнюю часть тоновой шкалы. Костная часть улитки была разрушена бором на границе ге средней и нижней трети, перепончатая часть и кортиев орган были травматизированы тонкой иглой. Уже на десятый день после операции все звуковые раздражители действовали, за исключением тона в 600 колебаний в секунду и ниже. Но затем в течение 3 месяцев после операции постепенно восстановлялось действие и тонов ниже 600, остановившись на 300 с небольшим. Многочисленные пробы за остальную часть времени до года после операции установили верхнюю границу исчезнувших тонов между 309 и 317 колебаниями в секунду. Нижняя не могла быть определена за неимением у нас чистых тонов ниже 100 колебаний в секунду.
Вот два протокола из последнего периода испытаний.
 Гистологического исследования не сделано, собака еще живет.
Очевидно, наш положительный опыт должен в этом вопросе стать на место отрицательных опытов Калишера (Kalischer).
Другая задача касалась вопроса: нужно ли для дифференцирования места звука содействие обоих полушарий? Решение вопроса дали опыты К. М. Быкова. У собаки перерезался corpus callosum. Когда животное оправлялось от операции, приступали к выработке пищевых условных рефлексов. Образование ик не представляло никаких особенностей и происходило так же скоро, как и у нормальных собак. Между прочим, у собаки имелся рефлекс на звук свистка в 1500 колебаний в секунду. Свисток, помещенный в картонный футляр, укреплялся на стене на уровне и на стороне левого уха в определенном расстоянии от станка с собакой. Рефлекс появился на восьмом разе применений и при 70 применениях сделался максимальным и постоянным. Затем тот же свисток помещался совершенно так же с правой стороны собаки. В этом положении звук свистка не сопровождался безусловным рефлексом. Применяя звук то слева, то справа, старались получить дифференцировку. Но ни малейшего намека на нее не оказалось, несмотря на 115 повторений звука справа, так что не было основания продолжать опыт дальше в таком виде. Нужно было заключить, что для дифференцирования места звука необходима соединенная работа полушарий.
Вот один из последних опытов.
Гистологического исследования не сделано, собака еще живет.
Очевидно, наш положительный опыт должен в этом вопросе стать на место отрицательных опытов Калишера (Kalischer).
Другая задача касалась вопроса: нужно ли для дифференцирования места звука содействие обоих полушарий? Решение вопроса дали опыты К. М. Быкова. У собаки перерезался corpus callosum. Когда животное оправлялось от операции, приступали к выработке пищевых условных рефлексов. Образование ик не представляло никаких особенностей и происходило так же скоро, как и у нормальных собак. Между прочим, у собаки имелся рефлекс на звук свистка в 1500 колебаний в секунду. Свисток, помещенный в картонный футляр, укреплялся на стене на уровне и на стороне левого уха в определенном расстоянии от станка с собакой. Рефлекс появился на восьмом разе применений и при 70 применениях сделался максимальным и постоянным. Затем тот же свисток помещался совершенно так же с правой стороны собаки. В этом положении звук свистка не сопровождался безусловным рефлексом. Применяя звук то слева, то справа, старались получить дифференцировку. Но ни малейшего намека на нее не оказалось, несмотря на 115 повторений звука справа, так что не было основания продолжать опыт дальше в таком виде. Нужно было заключить, что для дифференцирования места звука необходима соединенная работа полушарий.
Вот один из последних опытов.
 Другие дифференцировки давались этой собаке легко и скоро. На других нормальных собаках дифференцировка на место звука не отличалась по трудности от других, наступая уже после немногих повторений.
После всех фактов, приведенных в этой и предшествующей лекциях, едва ли можно оспаривать, что, действительно, все вопросы, составляющие доселе область так называемой физиологии органов чувств, могут разрабатываться на животных при помощи условных рефлексов. Сами знаменитые "бессознательные заключения» Гельмгольца (см. его «Физиологическую оптику» не суть ли истинные условные рефлексы? Возьмем простой случай хорошо переданного рисунком рельефа. Кожно-механические и двигательные раздражения, идущие от рельефа, суть первоначальные и основные раздражения, а световые раздражения от его более или менее освещенных и от более или менее затемненных мест представляют собой сигнальные условные раздражения, получившие свое жизненное значение лишь впоследствии в силу совпадения их во времени с первыми. Факты и другого рода, объективно воспроизводимые на собаках и совершенно отвечающие фактам из физиологии органов чувств, будут встречаться и дальше в наших лекциях.
Другие дифференцировки давались этой собаке легко и скоро. На других нормальных собаках дифференцировка на место звука не отличалась по трудности от других, наступая уже после немногих повторений.
После всех фактов, приведенных в этой и предшествующей лекциях, едва ли можно оспаривать, что, действительно, все вопросы, составляющие доселе область так называемой физиологии органов чувств, могут разрабатываться на животных при помощи условных рефлексов. Сами знаменитые "бессознательные заключения» Гельмгольца (см. его «Физиологическую оптику» не суть ли истинные условные рефлексы? Возьмем простой случай хорошо переданного рисунком рельефа. Кожно-механические и двигательные раздражения, идущие от рельефа, суть первоначальные и основные раздражения, а световые раздражения от его более или менее освещенных и от более или менее затемненных мест представляют собой сигнальные условные раздражения, получившие свое жизненное значение лишь впоследствии в силу совпадения их во времени с первыми. Факты и другого рода, объективно воспроизводимые на собаках и совершенно отвечающие фактам из физиологии органов чувств, будут встречаться и дальше в наших лекциях.
Лекция девятая. Иррадиирование и концентрирование нервных процессов в коре больших полушарии: а) иррадиирование и концентрирование тормозного процесса в отдельном анализаторе (кожном и звуковом)
Мм. гг.! До сих пор мы занимались главным образом, так сказать, внешней деятельностью больших полушарий, устанавливающей сложнейшие и тончайшие отношения между организмом и внешним миром. Прежде всего большие полушария бесчисленными агентами природы сигнализировали организму те относительно немногие агенты той же природы, которые как непосредственно благоприятствовали ему, так и были для него вредны, вызывая в нем соответствующие реакции. Дальше эти условно действующие агенты полушариями постоянно корригировались, когда они оказывались не отвечающиствительным отношениям, навсегда или временно лишаясь при определенных условиях их действия. Наконец гармонии с беспрерывно и многообразно колеблющейся природой агенты в качестве условных раздражителей то выделялись полушариями для организма виде крайне мелких элементов (анализировались), то сливались в многоразличные комплексы (синтезеровались). Теперь обратимся к внутреннему механизму работы больших полушарий. Что делается при описанной деятельности их c основными нервными процессами: раздражением и торможением? Первое, что здесь прежде всего привлекает себе внимание, это движение этих процессов, которым мы теперь и займемся. Как это часто бывает в физиологии с различными членами одной родственной группы явлений, изучение одного члена оказывается гораздо более удобным для исследователя, чем другого. При нашем исследовании в данном случае преимущество оказалось на стороне внутреннего торможения. Исследование его движения вместе с тем является блистательной иллюстрацией чрезвычайных выгод, которые предоставляет физиологу кожный анализатор с его огромной и вполне доступной рецепторной поверхностью. Вот наш основной исходный опыт, которым открылась для нас новая и важная глава физиологии больших полушарий. Этот опыт и несколько следующих за ним принадлежат Н. И. Красногорскому. О п ы т. Вдоль задней ноги собаки располагался ряд приборчиков (числом 5) для механического раздражения кожи, начиная со стопы вверх на расстоянии от самого нижнего приборчика на 3, 9, 15 и 22 см. Самый нижний приборчик служил для тормозного действия, все верхние - для положительного эффекта. Достигнуто это было, как обыкновенно, так, что сначала был выработан пищевой условный положительный рефлекс на одном месте. В силу первоначальной генерализации действовали более или менее и все остальные места. Затем раздражение на верхних 4 местах постоянно сопровождалось едой, пока положительный эффект от всех их не сделался почти или совершенно одинаковым, а раздражение нижнего места, наоборот, повторялось без сопровождения едой до тех пор, пока оно не лишилось всякого положительного действия, замененного теперь тормозным. В нижеприводимых протоколах условное раздражение, т. е. до присоединения безусловного, продолжается по 30 секунд каждый раз и обозначено номерами мест: 0 (самое нижнее), 1, 2, 3, 4 (по направлению кверху), причем при четырех верхних номерах в скобках указано расстояние верхних мест от нижнего. Конечно, положительные раздраженровождаются едой, тормозное - нет. Что мы видим в этом протоколе? Проба трех различных положительных раздражителей дала для всех них один и тот же слюноотделительный эффект -- 5 капель за 30 секунд. Затем при совершенно тождественных условиях времени, т. e. через минуту после троекратного применения тормозного раздражителя, испытываются то один, то другой положительные раздражители. Теперь их эффект совершенно различен. Ближайший к тормозному один раз остался без малейшего действия, в другой едва (часть капли) действовал. Следующий за ним вверх потерял почти половину своего эффекта, а самые верхние то остались без изменения, то даже несколько увеличились в их действии. Смысл опыта ясен. Так пункты кожи, конечно, представляют проекцию соответствующих пунктов коры больших полушарий, то из приведенного опыта и всех ему подобных неизбежно следует, что торможение, производимое раздражением на месте № 0 в соответствующем ему пункте коры, распространяется от него на соседние пункты и в тем более слабом размере,чем эти пункты далее лежат от пункта, где торможение производится, являясь совершенно не ленных пунктах. Значит, мы имеем перед собой иррадиирование тормозного процесса из исходного пункта по массе больших полушарий.
Если мы возьмем другую интенсивность торможения, суммируем не троекратное действие тормозного раздражителя, а или большее, или меньшее, а также возьмем и другой промежуток времени от конца развития тормозного процесса до начала применения положительных раздражителей, то мы, конечно, получим другие цифры для слюноотделения, но с совершенно тем же общим смыслом.
Вот протокол другого опыта, сюда относящегося.
Что мы видим в этом протоколе? Проба трех различных положительных раздражителей дала для всех них один и тот же слюноотделительный эффект -- 5 капель за 30 секунд. Затем при совершенно тождественных условиях времени, т. e. через минуту после троекратного применения тормозного раздражителя, испытываются то один, то другой положительные раздражители. Теперь их эффект совершенно различен. Ближайший к тормозному один раз остался без малейшего действия, в другой едва (часть капли) действовал. Следующий за ним вверх потерял почти половину своего эффекта, а самые верхние то остались без изменения, то даже несколько увеличились в их действии. Смысл опыта ясен. Так пункты кожи, конечно, представляют проекцию соответствующих пунктов коры больших полушарий, то из приведенного опыта и всех ему подобных неизбежно следует, что торможение, производимое раздражением на месте № 0 в соответствующем ему пункте коры, распространяется от него на соседние пункты и в тем более слабом размере,чем эти пункты далее лежат от пункта, где торможение производится, являясь совершенно не ленных пунктах. Значит, мы имеем перед собой иррадиирование тормозного процесса из исходного пункта по массе больших полушарий.
Если мы возьмем другую интенсивность торможения, суммируем не троекратное действие тормозного раздражителя, а или большее, или меньшее, а также возьмем и другой промежуток времени от конца развития тормозного процесса до начала применения положительных раздражителей, то мы, конечно, получим другие цифры для слюноотделения, но с совершенно тем же общим смыслом.
Вот протокол другого опыта, сюда относящегося.
 Таким образом более интенсивное торможение и при меньшем промежутке сказалось значительно и на самом дальнем пункте.
Если в одном и том же опыте пробовать различные положительные раздражители в различные сроки после окончания действия тормозного раздражителя, то можно с полной отчетливостью видеть, как тормозной процесс, сначала далеко распространившийся, затем постепенно оставляет сначала более далекие пункты, а потом и ближайшие к исходному пункту.
Привожу соответствующий опыт.
Таким образом более интенсивное торможение и при меньшем промежутке сказалось значительно и на самом дальнем пункте.
Если в одном и том же опыте пробовать различные положительные раздражители в различные сроки после окончания действия тормозного раздражителя, то можно с полной отчетливостью видеть, как тормозной процесс, сначала далеко распространившийся, затем постепенно оставляет сначала более далекие пункты, а потом и ближайшие к исходному пункту.
Привожу соответствующий опыт.
 Итак, № 4 был свободен от торможения через полминуты, № 2 - через 5 минут и № 1 - через 10 минут.
Чем чаще практикуется дифференцировка в течение дней и недель, тем освобождение дальних пунктов происходит все скорее, и это иногда обнаруживается на протяжении одного опыта при многократных повторениях положительных и отрицательных раздражений.
Достойно упоминания, что опыты эти демонстрировались много раз посторонним лицам и даже с полным успехом были проделаны на многолюдном заседании Медицинского общества.
Как понимать оставление тормозным процессом временно занимаемых им пунктов? Есть ли это уничтожение, затухание процесса на этих пунктах или это - как бы возврат, сосредоточение, концентрирование торможения в исходном пункте под влиянием какого-либо встречного процессор Принимая во внимание точный постоянно повторяющийся факт, что с практикой и укреплением дифференцировки совершенно параллельно идет и укорочение как во времени, так и в пространстве последовательного торможения от нее, естественно склоняться ко второму предположению, т. е. принимать, что мы имеем дело с противоположностью иррадиирования, с концентрированием процесса, не с его рассеянием и, следовательно, ослаблением, а с его сосредоточением и, следовательно, усилением в определенном пункте. Впоследствии мы приведем веские факты в пользу такого заключения.
Как мы только что видели, в опытах Красногорского концентрирование происходило очень медленно, минутами. Что касается иррадиирования, то оно в тех же опытах шло очень стремительно, так что его хода Красногорский уловить не мог, не видал.
Конечно, важность факта двустороннего движения тормозного процесса обязывала всячески варьировать опыты на эту тему. Ближайше наше внимание обратилось к торможению. имеющему место при угасании условных рефлексов (опыты Б. А. Когана), причем мы пользовались тем же кожным анализатором и его механическим раздражением, начиная с того же, что и в опытах Красногорского. Предварительно вырабатывался кислотный положительный кожно-механический условный рефлекс на каком-нибудь одном месте. Затем он как генерализованный сколько возможно выравнивался по всей поверхности одной стороны тела, и тогда приступали к опытам. В данный день подвергался раздражению один какой-нибудь пункт и отмечался его слюноотделительный эффект за одну минуту, но условное раздражение не сопровождалось безусловным рефлексом, условный рефлекс угашался. Через короткие промежутки (2 минуты) то же повторялось до тех пор, пока раздражение не сделалось впервые нулевым. Тогда в разные сроки времени по окончании нулевого раздражения производили раздражение то одного, то другого пункта кожи и также отмечали их эффекты за минуту. Эти другие пункты при определенных условиях времени оказывались также заторможенными в большей или меньшей степени. Первый пункт кожи, на котором производится угасание, мы называем первично угасаемым (как это уже указано в четвертой лекции), а эти последние - вторичнаемыми. Следовательно, и угасательное торможение не остается на том пункте мозга, где оно производится определенным внешним раздражением, а распространяется по мозговой массе, иррадиирует, т. е. получается то же, что мы видели торможении.
Понятно, что мы должны были в каждом отдельном опыте менять места для угашения, иначе мы имели бы спять дифференцировочное торможение, стойкое на выработанном месте, остающееся на большие сроки (многие месяцы), а не угасательное торможение, исчезающее в минуты и самое большее в 1-2 часа на первично угашаемсм месте.
Привожу опыты, иллюстрирующие это.
Итак, № 4 был свободен от торможения через полминуты, № 2 - через 5 минут и № 1 - через 10 минут.
Чем чаще практикуется дифференцировка в течение дней и недель, тем освобождение дальних пунктов происходит все скорее, и это иногда обнаруживается на протяжении одного опыта при многократных повторениях положительных и отрицательных раздражений.
Достойно упоминания, что опыты эти демонстрировались много раз посторонним лицам и даже с полным успехом были проделаны на многолюдном заседании Медицинского общества.
Как понимать оставление тормозным процессом временно занимаемых им пунктов? Есть ли это уничтожение, затухание процесса на этих пунктах или это - как бы возврат, сосредоточение, концентрирование торможения в исходном пункте под влиянием какого-либо встречного процессор Принимая во внимание точный постоянно повторяющийся факт, что с практикой и укреплением дифференцировки совершенно параллельно идет и укорочение как во времени, так и в пространстве последовательного торможения от нее, естественно склоняться ко второму предположению, т. е. принимать, что мы имеем дело с противоположностью иррадиирования, с концентрированием процесса, не с его рассеянием и, следовательно, ослаблением, а с его сосредоточением и, следовательно, усилением в определенном пункте. Впоследствии мы приведем веские факты в пользу такого заключения.
Как мы только что видели, в опытах Красногорского концентрирование происходило очень медленно, минутами. Что касается иррадиирования, то оно в тех же опытах шло очень стремительно, так что его хода Красногорский уловить не мог, не видал.
Конечно, важность факта двустороннего движения тормозного процесса обязывала всячески варьировать опыты на эту тему. Ближайше наше внимание обратилось к торможению. имеющему место при угасании условных рефлексов (опыты Б. А. Когана), причем мы пользовались тем же кожным анализатором и его механическим раздражением, начиная с того же, что и в опытах Красногорского. Предварительно вырабатывался кислотный положительный кожно-механический условный рефлекс на каком-нибудь одном месте. Затем он как генерализованный сколько возможно выравнивался по всей поверхности одной стороны тела, и тогда приступали к опытам. В данный день подвергался раздражению один какой-нибудь пункт и отмечался его слюноотделительный эффект за одну минуту, но условное раздражение не сопровождалось безусловным рефлексом, условный рефлекс угашался. Через короткие промежутки (2 минуты) то же повторялось до тех пор, пока раздражение не сделалось впервые нулевым. Тогда в разные сроки времени по окончании нулевого раздражения производили раздражение то одного, то другого пункта кожи и также отмечали их эффекты за минуту. Эти другие пункты при определенных условиях времени оказывались также заторможенными в большей или меньшей степени. Первый пункт кожи, на котором производится угасание, мы называем первично угасаемым (как это уже указано в четвертой лекции), а эти последние - вторичнаемыми. Следовательно, и угасательное торможение не остается на том пункте мозга, где оно производится определенным внешним раздражением, а распространяется по мозговой массе, иррадиирует, т. е. получается то же, что мы видели торможении.
Понятно, что мы должны были в каждом отдельном опыте менять места для угашения, иначе мы имели бы спять дифференцировочное торможение, стойкое на выработанном месте, остающееся на большие сроки (многие месяцы), а не угасательное торможение, исчезающее в минуты и самое большее в 1-2 часа на первично угашаемсм месте.
Привожу опыты, иллюстрирующие это.

 Ясно, что торможение на вторично угасаемом месте тем меньше, чем дальше отстоит оно от первично угасаемого места.
Теперь, если мы будем раздражать вторично угасаемое место, но через разные промежутки времени после нуля угасания, то увидим, что чем больше будет этот промежуток, тем торможение на этом месте становится все меньше, и наоборот.
Вот опыты, доказывающие это.
Ясно, что торможение на вторично угасаемом месте тем меньше, чем дальше отстоит оно от первично угасаемого места.
Теперь, если мы будем раздражать вторично угасаемое место, но через разные промежутки времени после нуля угасания, то увидим, что чем больше будет этот промежуток, тем торможение на этом месте становится все меньше, и наоборот.
Вот опыты, доказывающие это.

 Очевидно, что мы и тут имеем то, что назвали при диффетренировочном торможении концентрированием торможения, так как торможение с течением времени постепенно оставляет дальние пункты, приближаясь к исходному пункту.
Вместе с тем в приведенных опытах обращает на себя внимание одна подробность. Быстрота, которой торможение оставляет вторично тормозные пункты, очень различна у различных наших животных. То, что происходит у собаки № 1 в минуту, у собаки № 2 занимает 15 минут. Конечно, это очень важная сторона дела, характеризующая цифрами высшую нервную деятельность. Что в этом не играла роли какая-либо случайность, доказывается тем, что указываемая разница у трех животных, на которых производились эти опыты, осталась без изменения за много месяцев работы с ними.
В этом же ряде опытов (Когана) нам удалось видеть иррадиирование торможения в его ходе, т. е. постепенное распространение торможения из исходного пункта. Вот опыты, в которых мы испытывали состояние тормозного процесса на разных местах кожи, находящихся на разных расстояниях от первично угашаемого пункта, сейчас же, через нуль промежутка после полного угашения.
Привожу сюда относящиеся опыты.
Очевидно, что мы и тут имеем то, что назвали при диффетренировочном торможении концентрированием торможения, так как торможение с течением времени постепенно оставляет дальние пункты, приближаясь к исходному пункту.
Вместе с тем в приведенных опытах обращает на себя внимание одна подробность. Быстрота, которой торможение оставляет вторично тормозные пункты, очень различна у различных наших животных. То, что происходит у собаки № 1 в минуту, у собаки № 2 занимает 15 минут. Конечно, это очень важная сторона дела, характеризующая цифрами высшую нервную деятельность. Что в этом не играла роли какая-либо случайность, доказывается тем, что указываемая разница у трех животных, на которых производились эти опыты, осталась без изменения за много месяцев работы с ними.
В этом же ряде опытов (Когана) нам удалось видеть иррадиирование торможения в его ходе, т. е. постепенное распространение торможения из исходного пункта. Вот опыты, в которых мы испытывали состояние тормозного процесса на разных местах кожи, находящихся на разных расстояниях от первично угашаемого пункта, сейчас же, через нуль промежутка после полного угашения.
Привожу сюда относящиеся опыты.
 Сопоставление этих результатов с предшествующими, относящимися к той же собаке, ясно показывает, что непосредственно после полного развития торможения нервично угашаемом месте то же полное торможение имеется только на ближайших местах, тогда как на более удаленных оно почти отсутствует, едва намечается. Интересно, что у трех собак, примененных для этих опытов, состояние дальних пунктов призбираемом условии было очень различно. У собаки № 1 дальний пункт не только не заторможен, а, наоборот, дает сильно увеличенный положительный эффект, а у собаки № 3 он уже находится в значительно заторможенном состоянии.
Вот примеры.
Сопоставление этих результатов с предшествующими, относящимися к той же собаке, ясно показывает, что непосредственно после полного развития торможения нервично угашаемом месте то же полное торможение имеется только на ближайших местах, тогда как на более удаленных оно почти отсутствует, едва намечается. Интересно, что у трех собак, примененных для этих опытов, состояние дальних пунктов призбираемом условии было очень различно. У собаки № 1 дальний пункт не только не заторможен, а, наоборот, дает сильно увеличенный положительный эффект, а у собаки № 3 он уже находится в значительно заторможенном состоянии.
Вот примеры.
 Затем Коганом было собрано значительное число цифр, которое должно было представить более подробно все движение торможения у всех трех собак. Пlo этим данным оказывалось приблизительно следующее. У собаки № 1 период иррадиирования занял около 20 секунд, а период концентрирования продолжался до 75 секунд. У собаки № 2 иррадиирование закончилось только к 3 минутам, а все движение с концентрированием захватило период в 15 минут. У собаки № 3 соответственные цифры были 4-5 минут и 20 минут. Таким образом у всех этих собак, хотя продолжительность всего движения торможения была очень различна, отношение между периодамиррадиирования и концентрирования осталось почти постоянным: в 4-5 раз больше длилось концентрирование. Однако материал, сюда относящийся, нельзя считать вполне безупречным, потому что исходные величины условных рефлексов часто были очень различными, а в некоторых случаях нельзя было точно установить причину отклоняющихся цифр.
Относительно опытов Красногорского и Когана прошу обратить внимание на следующее. В некоторых случаях у Красногорского, но в особенности у Когана и именно на собаке № 1 (см. опыт 6 II 1914), очень часто при пробе удаленных мест, после полного угашения на первично угашенном месте, положительный эффект оказывался не только не угашенным, а в большей или меньшей мере, а иногда и в очень значительной, увеличенным. С этим особенным явлением мы подробно познакомимся через лекцию.
Нами также было изучено движение так называемого нами условного торможения (опыты Анрепа) и тоже на кожном анализаторе. Когда на разных местах кожи было выработано и выравнено механическое раздражение ее в качестве условного, на одном месте это положительное раздражение, комбинированное с раздражением из другого анализатора (электрический звонок) и повторяемое в этой комбинации без сопровождения безусловным рефлексом, было превращено в отрицательное, тормозное. Тормозным пунктом было сделано место на границе шеи и груди левой стороны (№ 0), места же на плече (№ 1), на запястье (№ 2), на границе груди и живота (№ 3), на тазу (№ 4), внизу бедра (№ 5) и, наконец, на предплюсне (№ 6) оставались положительными пунктами. Опыты исполнялись таким образом. Сперва в данном опыте пробовалось определенное место для установления цифры полного положительного условного рефлекса на нем. Затем применялась тормозная комбинация, и дальше через разные промежутки времени после нее повторялся положительный раздражитель. То же самое проделывалось в отдельных опытах со всеми остальными положительными местами. Как положительное, так и тормозное раздражение продолжалось по 30 секунд. Результаты представлены автором в следующей таблице.
Затем Коганом было собрано значительное число цифр, которое должно было представить более подробно все движение торможения у всех трех собак. Пlo этим данным оказывалось приблизительно следующее. У собаки № 1 период иррадиирования занял около 20 секунд, а период концентрирования продолжался до 75 секунд. У собаки № 2 иррадиирование закончилось только к 3 минутам, а все движение с концентрированием захватило период в 15 минут. У собаки № 3 соответственные цифры были 4-5 минут и 20 минут. Таким образом у всех этих собак, хотя продолжительность всего движения торможения была очень различна, отношение между периодамиррадиирования и концентрирования осталось почти постоянным: в 4-5 раз больше длилось концентрирование. Однако материал, сюда относящийся, нельзя считать вполне безупречным, потому что исходные величины условных рефлексов часто были очень различными, а в некоторых случаях нельзя было точно установить причину отклоняющихся цифр.
Относительно опытов Красногорского и Когана прошу обратить внимание на следующее. В некоторых случаях у Красногорского, но в особенности у Когана и именно на собаке № 1 (см. опыт 6 II 1914), очень часто при пробе удаленных мест, после полного угашения на первично угашенном месте, положительный эффект оказывался не только не угашенным, а в большей или меньшей мере, а иногда и в очень значительной, увеличенным. С этим особенным явлением мы подробно познакомимся через лекцию.
Нами также было изучено движение так называемого нами условного торможения (опыты Анрепа) и тоже на кожном анализаторе. Когда на разных местах кожи было выработано и выравнено механическое раздражение ее в качестве условного, на одном месте это положительное раздражение, комбинированное с раздражением из другого анализатора (электрический звонок) и повторяемое в этой комбинации без сопровождения безусловным рефлексом, было превращено в отрицательное, тормозное. Тормозным пунктом было сделано место на границе шеи и груди левой стороны (№ 0), места же на плече (№ 1), на запястье (№ 2), на границе груди и живота (№ 3), на тазу (№ 4), внизу бедра (№ 5) и, наконец, на предплюсне (№ 6) оставались положительными пунктами. Опыты исполнялись таким образом. Сперва в данном опыте пробовалось определенное место для установления цифры полного положительного условного рефлекса на нем. Затем применялась тормозная комбинация, и дальше через разные промежутки времени после нее повторялся положительный раздражитель. То же самое проделывалось в отдельных опытах со всеми остальными положительными местами. Как положительное, так и тормозное раздражение продолжалось по 30 секунд. Результаты представлены автором в следующей таблице.
 Первый вертикальный столбец - номера раздражаемых мест на коже. Следующие за ним - величины торможения на этих местах в процентах по отношению к величине положительного раздражения. Верхний горизонтальный рядромежутки времени в секундах по прекращении тормозного раздражения.
Как видим, и при условном торможении торможение, произведенное на одном определенном пункте, сперва распространяется по всему анализатору и тем в более слабой степени, чем другие пункты лежат дальше от исходного, причем максимальное торможение на всех пунктах достигается только через 30 секунд, а затем постепенно начинает ослабевать также на всех пунктах одновременно. Исключение представляет только пункт № 0, где торможение сразу оказывается максимальным, а не постепенно нарастающим. Конечно, и здесь в опытах действуют нарушающим образом случайные посторонние раздражители, то тормозя положительные рефлексы, то растормаживая тормозные в большей или меньшей степени.
В трех описанных рядах опытов над движением торможения: дифференцировочного, угасательного и так называемого условного, согласно устанавливается, во-первых, факт распространения торможения из пункта, где оно соответственным образом производится, по всему анализатору, а, во-вторых, постепенный его уход. Но в подробностях этого факта имеются значительные и даже иногда существенные разницы. При дифференцировочном торможении у Красногорского иррадиирование торможения происходит моментально, неуловимо для экспериментатора, и только концентрирование совершается в течение определенного срока. У Когана и торможение распространяется по анализатору тоже постепенно, только гораздо скорее, чем происходит концентрирование. Эту разницу можно было бы рассматривать как зависящую от интенсивности развиваемого торможения. Красногорский обыкновенно повторял тормозной раздражитель, несмотря на его полное действие сразу, несколько раз. Коган ограничивался при угасании первым нулем. Но разница с условным торможением гораздо сложнее и существеннее. Условное торможение у Анрепа, как дифференцировочное торможение у Красногорского, распространяется одновременно, хотя в разной степени по всему анализатору, но затем, в отличие от Красногорского, в течение определенного срока, опять же одновременно на всех пунктах, постепенно усиливается до известного максимума. После максимума, также одновременно всех пунктах, начинается постепенное ослабление торможения. Таким образом при условном торможении, в отличие от дифференцировочного и угасательного, не видно самого перехода из пункта в пункт ни в одном, ни в другом направлении, ни вперед, ни назад к исходному пункту.
Так как новая область перед нами все расширялась и углублялась и специально вопрос о движении нервных процессов в коре больших полушарий все усложнялся, как увидим это дальше, то я считаю неизбежным часто передавать наш материал исторически (на протяжении нашей двадцатипятилетней работы), не находя возможным сейчас же на всех пунктах исследования придти к объединяющим и окончательным заключениям.
В только что переданной работе Анрепа приведен подробно факт, уже частью намечавшийся и у других авторов ранее, именно, что выработанные как положительные, так и отрицательные условные рефлексы на разных местах одной половины тела с их колебаниями при движении тов мо же ний (пос лед нее специально у Анрепа) изумительно точно воспроизводятся на симметричных местах другой половины тела сами собой, без малейшей выработки. Этот интересный факт с некоторым его анализом будет описан позже.
Первый вертикальный столбец - номера раздражаемых мест на коже. Следующие за ним - величины торможения на этих местах в процентах по отношению к величине положительного раздражения. Верхний горизонтальный рядромежутки времени в секундах по прекращении тормозного раздражения.
Как видим, и при условном торможении торможение, произведенное на одном определенном пункте, сперва распространяется по всему анализатору и тем в более слабой степени, чем другие пункты лежат дальше от исходного, причем максимальное торможение на всех пунктах достигается только через 30 секунд, а затем постепенно начинает ослабевать также на всех пунктах одновременно. Исключение представляет только пункт № 0, где торможение сразу оказывается максимальным, а не постепенно нарастающим. Конечно, и здесь в опытах действуют нарушающим образом случайные посторонние раздражители, то тормозя положительные рефлексы, то растормаживая тормозные в большей или меньшей степени.
В трех описанных рядах опытов над движением торможения: дифференцировочного, угасательного и так называемого условного, согласно устанавливается, во-первых, факт распространения торможения из пункта, где оно соответственным образом производится, по всему анализатору, а, во-вторых, постепенный его уход. Но в подробностях этого факта имеются значительные и даже иногда существенные разницы. При дифференцировочном торможении у Красногорского иррадиирование торможения происходит моментально, неуловимо для экспериментатора, и только концентрирование совершается в течение определенного срока. У Когана и торможение распространяется по анализатору тоже постепенно, только гораздо скорее, чем происходит концентрирование. Эту разницу можно было бы рассматривать как зависящую от интенсивности развиваемого торможения. Красногорский обыкновенно повторял тормозной раздражитель, несмотря на его полное действие сразу, несколько раз. Коган ограничивался при угасании первым нулем. Но разница с условным торможением гораздо сложнее и существеннее. Условное торможение у Анрепа, как дифференцировочное торможение у Красногорского, распространяется одновременно, хотя в разной степени по всему анализатору, но затем, в отличие от Красногорского, в течение определенного срока, опять же одновременно на всех пунктах, постепенно усиливается до известного максимума. После максимума, также одновременно всех пунктах, начинается постепенное ослабление торможения. Таким образом при условном торможении, в отличие от дифференцировочного и угасательного, не видно самого перехода из пункта в пункт ни в одном, ни в другом направлении, ни вперед, ни назад к исходному пункту.
Так как новая область перед нами все расширялась и углублялась и специально вопрос о движении нервных процессов в коре больших полушарий все усложнялся, как увидим это дальше, то я считаю неизбежным часто передавать наш материал исторически (на протяжении нашей двадцатипятилетней работы), не находя возможным сейчас же на всех пунктах исследования придти к объединяющим и окончательным заключениям.
В только что переданной работе Анрепа приведен подробно факт, уже частью намечавшийся и у других авторов ранее, именно, что выработанные как положительные, так и отрицательные условные рефлексы на разных местах одной половины тела с их колебаниями при движении тов мо же ний (пос лед нее специально у Анрепа) изумительно точно воспроизводятся на симметричных местах другой половины тела сами собой, без малейшей выработки. Этот интересный факт с некоторым его анализом будет описан позже.
 Опыты, аналогичные опытам движения торможения по кожному анализатору, мы пробовали ставить на ушном (звуковом) анализаторе, также предполагая соответствующую периферическому концу его проекцию в коре больших полушарий. С этой целью мы (Мануйлов и Иванов-Смоленский) брали разные звуки (разные тоны, удары метронома и пипение) в качестве условных раздражителей и, угашая один из них, следили за тем, как это сказывается на других.
Привожу относящиеся сюда опыты Иванова-Смоленского. У собаки были выработаны отдельные пищевые условные рефлексы на четыре тона, получаемого от тонвариатора на две пары смежных тонов, разделенных тремя октавами: си большой октавы (Н 123 колебания), до малой октавы (С 130 колебаний) и до и ре (D 1161 колебание, С 1036) третьей октавы, а также на шипение и удары метронома (100 минуту). Опыт велся таким образом, что угашался один какой-либо условный рефлекс до нуля, а затем или непосредственно за этих или через минуты: 1, 3, 5, 7, 10, 12 и 15, испытывались разных опытах все остальные рефлексы. Все рефлексы оказывались заторможенными, но в разной степени. Результаты частично представлены в следующей таблице.
Опыты, аналогичные опытам движения торможения по кожному анализатору, мы пробовали ставить на ушном (звуковом) анализаторе, также предполагая соответствующую периферическому концу его проекцию в коре больших полушарий. С этой целью мы (Мануйлов и Иванов-Смоленский) брали разные звуки (разные тоны, удары метронома и пипение) в качестве условных раздражителей и, угашая один из них, следили за тем, как это сказывается на других.
Привожу относящиеся сюда опыты Иванова-Смоленского. У собаки были выработаны отдельные пищевые условные рефлексы на четыре тона, получаемого от тонвариатора на две пары смежных тонов, разделенных тремя октавами: си большой октавы (Н 123 колебания), до малой октавы (С 130 колебаний) и до и ре (D 1161 колебание, С 1036) третьей октавы, а также на шипение и удары метронома (100 минуту). Опыт велся таким образом, что угашался один какой-либо условный рефлекс до нуля, а затем или непосредственно за этих или через минуты: 1, 3, 5, 7, 10, 12 и 15, испытывались разных опытах все остальные рефлексы. Все рефлексы оказывались заторможенными, но в разной степени. Результаты частично представлены в следующей таблице.

 Таблица показывает, что при угашении одного тона из пары низких рефлекс на другой низкий скорее доходит до максимума торможения, более долгое время остается на максимуме и медленнее освобождается от него, чем рефлексы на каждый из высоких. При угашении одного из высоких тонов рефлекс на другой высокий также скорее доходит до максимума торможения, более долгое время остается на максимуме и медленнее освобождается от него, чем рефлексы на каждый из низких. При угашении всякого из наших тонов вторичное торможение рефлексов на шипение и удары метронома во всех отношениях выражено гораздо слабее, чем на тоновых рефлексах, и, наоборот, при удалении рефлексов на шипение и удары метронома вторичное торможение всех рефлексов на тоны обнаруживается очень энергично во всех отношениях.
Конечно, только относительно опытов с удалением тонов и с последовательной пробой их можно более или менее с основанием говорить о движении торможения по корковому концу звукового анализатора как проекции кортиева органа. При взаимном влиянии угашенных рефлексов на тоны, с одной стороны, и на шипение и удары метронома - с другой, результат, вероятнее всего, определяется различной интенсивностью этих различных раздражителей.
Нужно принимать, что то, что очевидно на кожном анализаторе и обнаруживается также довольно отчетливо на звуковом, т. е. движение тормозного процесса из пункта в пункт на мозговом конце анализатора, имеет место и на других анализаторах. Но тут констатирование его или трудно, или даже пока и невозможно по явным техническим условиям. Установление же подробностей этого факта могло бы дать основу для представления об элементарном составе этих анализаторов.
Таблица показывает, что при угашении одного тона из пары низких рефлекс на другой низкий скорее доходит до максимума торможения, более долгое время остается на максимуме и медленнее освобождается от него, чем рефлексы на каждый из высоких. При угашении одного из высоких тонов рефлекс на другой высокий также скорее доходит до максимума торможения, более долгое время остается на максимуме и медленнее освобождается от него, чем рефлексы на каждый из низких. При угашении всякого из наших тонов вторичное торможение рефлексов на шипение и удары метронома во всех отношениях выражено гораздо слабее, чем на тоновых рефлексах, и, наоборот, при удалении рефлексов на шипение и удары метронома вторичное торможение всех рефлексов на тоны обнаруживается очень энергично во всех отношениях.
Конечно, только относительно опытов с удалением тонов и с последовательной пробой их можно более или менее с основанием говорить о движении торможения по корковому концу звукового анализатора как проекции кортиева органа. При взаимном влиянии угашенных рефлексов на тоны, с одной стороны, и на шипение и удары метронома - с другой, результат, вероятнее всего, определяется различной интенсивностью этих различных раздражителей.
Нужно принимать, что то, что очевидно на кожном анализаторе и обнаруживается также довольно отчетливо на звуковом, т. е. движение тормозного процесса из пункта в пункт на мозговом конце анализатора, имеет место и на других анализаторах. Но тут констатирование его или трудно, или даже пока и невозможно по явным техническим условиям. Установление же подробностей этого факта могло бы дать основу для представления об элементарном составе этих анализаторов.
Лекция десятая. Иррадиирование и концентрирование нервных процессов коре больших полушарии: 6) иррадиирование и концентрирование тормозного процесса по большим полушариям; в) иррадиирование и концентрирование раздражительного процесса
Мм. гг.! В прошлой лекции мы приступили к изучению движения нервных процессов в коре больших полушарий. Начав с внутреннего торможения, мы из опытов убедились в том, что оно, будучи произведено соответствующим раздражением в определенном пункте, сперва быстро иррадиирует по своему анализатору, а затем медленно концентрируется в направлении к исходному пункту, и что даже можно наблюдать это двустороннее движение шаг за шагом. В настоящей лекция мы будем продолжать следить за этим движением по всему полушарию, т. е. за переходом торможения из анализатора в анализатор. Опыты, сюда относящиеся, проделаны со всеми нашими случаями внутреннего торможения. При описании дифференцировочного торможения было уже сообщено, что торможение, произведенное в одном анализаторе, обнаруживается и в других. В опытах над этим торможением (Белякова) оказалось, что при слабой степени его оно не проявляется в другом анализаторе, при сильном же его напряжении оно выступает и тут, но в гораздо меньшей мере, чем в исходном анализаторе. Вот несколько из этого ряда опытов. У собаки между прочими выработаны отдельные пищевые условные рефлексы: на звук гальтоновского свистка в 4000 колебаний в секунду и на бесшумное движение вертящегося перед глазами собаки предмета. Звуковой рефлекс обыкновенно составляют 11-12 капель за 30 секунд, зрительный 7--8 капель. Звук на полтона ниже - дифференцировка. Смысл опытов ясный. После двукратного применения подряд дифференцированного тона и минуту спустя после этого положительный тон теряет в его действии больше половины, зрительный же раздражитель не терпит ни малейшего ущерба. После большего суммирования торможения, после четырехкратного применения (да еще при меньшей паузе) дифференцированного тона, и также минуту спустя, оказывается заторможенным и зрительный рефлекс, но только немного больше чем в два раза, между тем как в тех же условиях звуковой рефлекс задерживается почти полностью.
Совершенно то же самое получается при угасательном торможении, как показывают опыты 9. Л. Горна. Здесь, благодаря некоторой вариации опыта, еще убедительнее выступает факт движения тормозного процесса, что касается специально более раннего ухода его из другого анализатора. Вот некоторые из этих опытов.
У собаки образованы между прочими отдельные пищевые условные рефлексы на тон cis духового камертона и на вспыхивание перед глазами собаки трех шестнадцатисвечных лампочек.
Смысл опытов ясный. После двукратного применения подряд дифференцированного тона и минуту спустя после этого положительный тон теряет в его действии больше половины, зрительный же раздражитель не терпит ни малейшего ущерба. После большего суммирования торможения, после четырехкратного применения (да еще при меньшей паузе) дифференцированного тона, и также минуту спустя, оказывается заторможенным и зрительный рефлекс, но только немного больше чем в два раза, между тем как в тех же условиях звуковой рефлекс задерживается почти полностью.
Совершенно то же самое получается при угасательном торможении, как показывают опыты 9. Л. Горна. Здесь, благодаря некоторой вариации опыта, еще убедительнее выступает факт движения тормозного процесса, что касается специально более раннего ухода его из другого анализатора. Вот некоторые из этих опытов.
У собаки образованы между прочими отдельные пищевые условные рефлексы на тон cis духового камертона и на вспыхивание перед глазами собаки трех шестнадцатисвечных лампочек.

 Угашался рефлекс на свет, и испытывалось вторичное действие на тоновой рефлекс. Световой рефлекс, угашенный до степени двух нулей и предоставленный себе, начал восстанавливаться лишь через 24 минуты. Тоновой рефлекс, после такой же степени угашения светового рефлекса примененный непосредственно без всякого промежутка, потерял в своем эффекте только около половины, а испытанный в другом опыте через 3 минуты оказался уже совершенно свободным от торможения. Следовательно, тормозной процесс из светового анализатора распространился на звуковой только в слабой мере и скорее ушел оттуда совсем. Что он ушел оттуда скорее не потому, что был менее значительным, доказывается дальнейшими подробностями опытов. В световом анализаторе промежуток между повторениями (3 минуты) на всех степенях угасания постоянно влек за собой только падение рефлекса, а в звуковом тот же промежуток повел к полному восстановлению рефлекса. Кроме того, во втором из приведенных опытов при угашении светового рефлекса, когда этот рефлекс представлял уже небольшую величину, даже втрое экстренно увеличенный промежуток только остановил дальнейшее уменьшение рефлекса, но не обусловил сколько-нибудь значительного восстановления его. Что, наконец, разница в восстановлении первично и вторично угашаемых рефлексов не зависела в данном случае от особенностей световых и звуковых клеток, свидетельствуют другие наши опыты, где первично угашался тоновой рефлекс, а вторичное угашение наблюдалось на световом, и где, однако, отношения остались совершенно теми же. Ввиду полного тождества этих и уже приведенных опытов я их не привожу. Таким образом можно с правом принимать, что во вторично угашаемом анализаторе мы имеем дело с периферией иррадиирующего процесса и что с нее начинается обратное движение процесса, концентрирование его, совершенно так, как это мы наблюдали внутри кожного анализатора.
Опыты с движением угасательного торможения по целому полушарию вообще нами очень разнообразились. Мы, имея в первично угашаемом анализаторе два разных раздражителя (например вспыхивание лампочек и верчение предмета, стоящего перед глазами животного) и угашая один из них, следили за торможением как в другом анализаторе, так и в том же анализаторе на обоих раздражителях. Порядок освобождения от иррадиирующего торможения был таков: скорее всего освобождался рефлекс из другого анализатора; близко к нему по времени, но несколько медленнее, вторично угашаемый рефлекс из первично угашаемого анализатора и, наконец, сильно запаздывая, первично угашенный рефлекс. Последнее, т. e. разница между рефлексами, принадлежащими к одному и тому же анализатору, очевидно, указывает на территориальное разграничение в нем различных рефлексов, с чем мы уже познакомились в предшествующей лекции.
Те же результаты дали опыты и над движением условного торможения. Один и тот же внешний агент был сделан условным тормозом нескольких условных раздражителей, принадлежащих к разным анализаторам, т. е. комбинация его с каждым из них отдельно повторялась без сопровождения безусловным раздражителем и таким образом превращалась в тормозную. Опыт исполнялся таким образом. Испытывался один из этих условных раздражителей и отмечался его слюноотделительный эффект. Затем несколько раз повторялась тормозная комбинация или с этим раздражителем, или с условным раздражителем из другого анализатора. Опыт кончался вторичной пробой первого раздражителя через разные промежутки времени после последнего повторения тормозной комбинации. Если тормозная комбинация включала первый раздражитель, то он теперь последовательно оказывался сильно заторможенным и восстановлялся медленно до его исходной величины. Если же в состав тормозной комбинации входил раздражитель из другого анализатора, то испытанный вначале раздражитель был заторможен только в слабой степени и быстро возвращался к его первоначальной величине. Это значило, что последовательное торможение в анализаторе условно-тормозимого раздражителя было очень значительно и долго в нем держалось, в другой же анализатор оно проникло только в малой мере и скоро оттуда ушло.
Вот опыты из работ В. А. Дегтяревой.
У собаки отдельные пищевые условные рефлексы на удары метронома (M) и вспыхивание электрических лампочек (Л). Условный тормоз к тому и другому бесшумное верчение стояшего перед собакой предмета (В). И условные раздражители и тормозная комбинация продолжаются каждый раз минуту, также и слюноотделение отмечается в каплях за минуту.
Угашался рефлекс на свет, и испытывалось вторичное действие на тоновой рефлекс. Световой рефлекс, угашенный до степени двух нулей и предоставленный себе, начал восстанавливаться лишь через 24 минуты. Тоновой рефлекс, после такой же степени угашения светового рефлекса примененный непосредственно без всякого промежутка, потерял в своем эффекте только около половины, а испытанный в другом опыте через 3 минуты оказался уже совершенно свободным от торможения. Следовательно, тормозной процесс из светового анализатора распространился на звуковой только в слабой мере и скорее ушел оттуда совсем. Что он ушел оттуда скорее не потому, что был менее значительным, доказывается дальнейшими подробностями опытов. В световом анализаторе промежуток между повторениями (3 минуты) на всех степенях угасания постоянно влек за собой только падение рефлекса, а в звуковом тот же промежуток повел к полному восстановлению рефлекса. Кроме того, во втором из приведенных опытов при угашении светового рефлекса, когда этот рефлекс представлял уже небольшую величину, даже втрое экстренно увеличенный промежуток только остановил дальнейшее уменьшение рефлекса, но не обусловил сколько-нибудь значительного восстановления его. Что, наконец, разница в восстановлении первично и вторично угашаемых рефлексов не зависела в данном случае от особенностей световых и звуковых клеток, свидетельствуют другие наши опыты, где первично угашался тоновой рефлекс, а вторичное угашение наблюдалось на световом, и где, однако, отношения остались совершенно теми же. Ввиду полного тождества этих и уже приведенных опытов я их не привожу. Таким образом можно с правом принимать, что во вторично угашаемом анализаторе мы имеем дело с периферией иррадиирующего процесса и что с нее начинается обратное движение процесса, концентрирование его, совершенно так, как это мы наблюдали внутри кожного анализатора.
Опыты с движением угасательного торможения по целому полушарию вообще нами очень разнообразились. Мы, имея в первично угашаемом анализаторе два разных раздражителя (например вспыхивание лампочек и верчение предмета, стоящего перед глазами животного) и угашая один из них, следили за торможением как в другом анализаторе, так и в том же анализаторе на обоих раздражителях. Порядок освобождения от иррадиирующего торможения был таков: скорее всего освобождался рефлекс из другого анализатора; близко к нему по времени, но несколько медленнее, вторично угашаемый рефлекс из первично угашаемого анализатора и, наконец, сильно запаздывая, первично угашенный рефлекс. Последнее, т. e. разница между рефлексами, принадлежащими к одному и тому же анализатору, очевидно, указывает на территориальное разграничение в нем различных рефлексов, с чем мы уже познакомились в предшествующей лекции.
Те же результаты дали опыты и над движением условного торможения. Один и тот же внешний агент был сделан условным тормозом нескольких условных раздражителей, принадлежащих к разным анализаторам, т. е. комбинация его с каждым из них отдельно повторялась без сопровождения безусловным раздражителем и таким образом превращалась в тормозную. Опыт исполнялся таким образом. Испытывался один из этих условных раздражителей и отмечался его слюноотделительный эффект. Затем несколько раз повторялась тормозная комбинация или с этим раздражителем, или с условным раздражителем из другого анализатора. Опыт кончался вторичной пробой первого раздражителя через разные промежутки времени после последнего повторения тормозной комбинации. Если тормозная комбинация включала первый раздражитель, то он теперь последовательно оказывался сильно заторможенным и восстановлялся медленно до его исходной величины. Если же в состав тормозной комбинации входил раздражитель из другого анализатора, то испытанный вначале раздражитель был заторможен только в слабой степени и быстро возвращался к его первоначальной величине. Это значило, что последовательное торможение в анализаторе условно-тормозимого раздражителя было очень значительно и долго в нем держалось, в другой же анализатор оно проникло только в малой мере и скоро оттуда ушло.
Вот опыты из работ В. А. Дегтяревой.
У собаки отдельные пищевые условные рефлексы на удары метронома (M) и вспыхивание электрических лампочек (Л). Условный тормоз к тому и другому бесшумное верчение стояшего перед собакой предмета (В). И условные раздражители и тормозная комбинация продолжаются каждый раз минуту, также и слюноотделение отмечается в каплях за минуту.
 Метрономный рефлекс после пятикратного применения тормозной комбинации, в которой он сам участвовал, восстановился вполне только 7 минут спустя; после же комбинации без него он был почти полный уже только через 15 секунд.
Наконец нами было исследовано движение торможения, которое развивается при запаздывании положительного эффекта в сильно отставленных рефлексах, предшествуя деятельной фазе. Соответственно особой постановке торможения, как фазовому явлению, надо было ожидать некоторых особенностей в опытах над его движением, что и оказалось. Между разными собаками наблюдалась большая разница в результате наших испытании. У некоторых животных удавалось легко показать распространение этого торможения в другие анализаторы. Делалось это таким образом. Раздражитель запаздывающего рефлекса прекращался во время недеятельной фазы, и непосредственно или спустя некоторое время пускался в ход условный раздражитель из другого анализатора, предварительно испробованный на его величину. Вот протоколы таких опытов (Горна).
У данной собаки имеются между прочими следующие кислотные условные рефлексы: отставленные на 30 секунд - один на вспыхивание электрических лампочек, другой на кожно-механическое раздражение, и отставленный на 3 минуты на удары метронома.
Метрономный рефлекс после пятикратного применения тормозной комбинации, в которой он сам участвовал, восстановился вполне только 7 минут спустя; после же комбинации без него он был почти полный уже только через 15 секунд.
Наконец нами было исследовано движение торможения, которое развивается при запаздывании положительного эффекта в сильно отставленных рефлексах, предшествуя деятельной фазе. Соответственно особой постановке торможения, как фазовому явлению, надо было ожидать некоторых особенностей в опытах над его движением, что и оказалось. Между разными собаками наблюдалась большая разница в результате наших испытании. У некоторых животных удавалось легко показать распространение этого торможения в другие анализаторы. Делалось это таким образом. Раздражитель запаздывающего рефлекса прекращался во время недеятельной фазы, и непосредственно или спустя некоторое время пускался в ход условный раздражитель из другого анализатора, предварительно испробованный на его величину. Вот протоколы таких опытов (Горна).
У данной собаки имеются между прочими следующие кислотные условные рефлексы: отставленные на 30 секунд - один на вспыхивание электрических лампочек, другой на кожно-механическое раздражение, и отставленный на 3 минуты на удары метронома.
 Результат вполне резкий. Процесс торможения, вызываемый метрономом в начале его действия, проник из звукового анализатора как в зрительный, так и в кожный анализаторы и сказался в уменьшении их рефлексов в 2 раза.
Но это у одной собаки. У другой результат был скорее обратный. Условные раздражители, примененные непосредственно после недеятельной фазы отставленного рефлекса, даже несколько увеличились в своем размере. Причина этой разницы была ясна. Когда у первой собаки между изолированной недеятельной фазой и пробой другого рефлекса ввели промежуток в 2 минуты, то в течение этих 2 минут деятельная фаза не наступила, т. е. тормозная фаза действительно была точно выделена, и поэтому последовательное торможение на другом рефлексе было налицо. У второй же собаки при тех же условиях в промежутке обнаружилась деятельная фаза и, естественно, другой рефлекс был не уменьшенным, а увеличенным. Следовательно, у этой собаки ранее и энергичнее выступил раздражительный процесс, т. e. недеятельная фаза не была полностью изолирована. С этим совпадало и то, что у первой собаки тормозной процесс вообще заметно преобладал сравнительно с раздражительным.
В прошлой и настоящей лекция привел в изобилии опыты, иллюстрирующие движение тормозного процесса по массе больших полушарий. Вам могло показаться, что это было даже и излишне. Но я сделал это с умыслом, чтобы сделать для вас очевидным, как много раз мы наблюдали это явление в более или менее простом, так сказать в чистом, виде. И все же это был только, если можно так выразиться, основной скелет явления, выступавший перед нами благодаря или особенностям экспериментирования, или индивидуальным особенностям животных, скелет, однако, наиболее часто покрываемый добавочными элементами, очень усложняющими, так сказать, полную действительность.
Первое, на что мы уже давно натолкнулись в этих опытах, состояло в следующем.
В описанных в прошлой лекции опытах (Красногорского) об иррадиирование и концентрирормозного процесса в кожном анализаторе встретился такой неожиданный тогда для нас факт. При предварительной пробе в опыте условное механическое раздражение места кожи, удаленного от тормозного пункта на 22 см, дало 6 капель слюны за 30 секунд, а раздражение зрительного анализатора дало 5 капель. Производится троекратное раздражение тормозного пункта. Минуту спустя после последнего раздражения испытывается зрительное раздражение - нуль слюноотделения, т. e. полное последовательное торможение. Через 10 минут применяется теперь четырехкратное раздражение тормозного пункта. Через минуту после него раздражение места, удаленного на 22 см, сопровождается полным эффектом - 8 каплями.
Подобный же факт наблюдался в других наших опытах (Чеботаревой). У собаки имелось несколько условных рефлексов на раздражители из разных анализаторов. Условный метрономный раздражитель, примененный вместе с условным тормозом (из зрительного анализатора), не давал ни капли слюны. 1-2 минуты спустя после того метроном один вполне восстановлял свой обычный слюноотделительный эффект и был при этом подкрепляем, а испытанные вскоре после того условные рефлексы на механическое раздражение кожи и на запах камфоры оказывались еще значительно заторможенными.
Таким образом выходило, что торможение через некоторый срок, после того как оно было произведено в данном анализаторе, уже исчезало из него, между тем в других анализаторах даже через большие сроки оно еще давало себя знать.
Но вместе с тем было обращено внимание на то, что это случалось тогда, когда условные рефлексы из других анализаторов были очень молодые, т. e. только что образованные, или хотя и старые, но давно не практиковавшиеся, или, наконец. вообще слабые, выработанные на агентов малой интенсивности. Ввиду этого можно было себе представить, что, когда известный пункт, ранее заторможенный, оказывался свободным от торможения, это еще не значило, что в его районе не было более торможения. В этом пункте раздражительный процесс, в силу определенных условий, мог брать перевес над тормозным процессом, делаться, так сказать, иммунным по отношению к окружающему его, но уже ослабленному тормозному процессу. И действительно, это постоянный факт, что рефлексы с еще недостаточно концентрированным или вообще со слабым раздеажительным процессом чрезвычайно легко подвергаются действию как внешнего, так и внутреннего торможения, между тем как сильные рефлексы остаются вне влияния тех же торможений. Об этой, так сказать, крепости или даже неприкосновенности сильных и хорошо выработанных как раздражительных, так и тормозных пунктов будет речь еще впереди.
В свое время, когда впервые наблюдались случаи, как бы противоречащие правилу иррадиирования и концентрирования торможения, были исполнены следующие нарочитые опыты (А. М. Павловой).
У собак были выработаны условные рефлексы на раздражители из разных анализаторов, причем обращалось тщательное внимание на то, чтобы все они повторялись одинаковое число раз. Когда все они достигают максимума эффекта, с одним из них комбинируется новый внешний агент. и эта комбинация обычным методом превращается в тормозную. Тормозная комбинация, а после нее ее условный раздражитель повторяются до тех пор, пока последовательное торможение комбинацией положительного раздражителя не ограничится 2 минутами, т. е. после 2 минут уже незаметно на нем. Конечно, и за этот период соблюдается требование об одинаковой практике всех рефлексов. Затем все остальные рефлексы впервые испытываются совместно c условным тормозом. Оказывается, что и эти рефлексы, за исключением одного, тоже освобождаются от торможения к концу 2 минут после применения тормозной комбинации. Исключение приходится на условный рефлекс из слабого раздражения светом, который обычно отстает по слюноотделительному эффекту от остальных рефлексов. Отсюда следует, что одна и та же степень торможения не сказывается на сильных раздражителях и отчетливо дает себя знать на слабых.
Но это только один элемент, осложняющий факт иррадиирования и концентрирования тормозного процесса. Впоследствии и постепенно мы познакомились и c другим, имеющим гораздо большее значение и еще более нарушающим простой ход иррадиирования и концентрирования. Об этом элементе мы сообщим имеющийся у нас материал в следующей лекции.
А теперь я перехожу к нашим опытам относительно движения раздражительного процесса. Этот вопрос обработан у нас в гораздо меньшей степени.
Наш первый опыт (М. К. Петровой) с иррадиированием раздражительного процесса исполнен советшенно по типу тех опытов, которыми мы начали изучение иррадиирование и концентрирования тормозного процессбак вдоль задней конечности, начиная с плюсны до таза, были расположены пять приборчиков для механического раздражения кожи, приблизительно на равных друг от друга расстояниях. Из действия самого нижнего приборчика (№ 1) у одной собаки сделан условный кислотный рефлекс, у другой - пищевой. Верхние приборчики (№ 2, 3, 4 и 5) в силу генерализации, конечно, вызывали также соответствующие рефлексы, но они обычным приемом были отдифференцированы и доведены все одинаково до нуля эффекта. Тогда было приступлено к главной задаче опытоб. Сперва испытывалось действие самого нижнего приборчика, т. е. положительный раздражитель в течение 30 секунд, причем слюноотделение за каждые 15 секунд отмечалось отдельно, a затем, как обыкновенно, присоединялся безусловный раздражитель. Спустя некоторое время нижний приборчик опять пускался в ход, но только на 15 секунд, а затем сейчас же после него начинали действовать, и тоже в течение только 15 секунд, верхние приборчики, то один, то другой в разных опытах, то более близкий, то более дальний. Получилось следующее. При смене № 1 на № 2 (ближайший) слюноотделение в течение действия этого приборчика былобыкновенно много меньше, а иногда и равно слюноотделению, наблюдавшемуся в течение 15 секунд действия приборчика № 1. При смене таким же образом № 1 на № 5, т. е. самого дальнего, слюноотделение при нем резко падало. Приборчики № 3 и 4, примененные в тождественной обстановке, дали средние цифры, приближаясь № 3 к № 2 и № 4 к № 5. В другом ряде опытов отмечалось слюноотделение не только за время действия тормозных раздражителей, но до конца отделительного периода. Тогда разница в действии близких и дальних приборчиков выступала почти без единого исключения и гораздо рельефнее. Отделительный период продолжался дольше и был гораздо значительнее по величине после действия близкого приборчика, чем дальнего.
Привожу данные ряда опытных дней.
У этой собаки кислотный условный рефлекс.
Результат вполне резкий. Процесс торможения, вызываемый метрономом в начале его действия, проник из звукового анализатора как в зрительный, так и в кожный анализаторы и сказался в уменьшении их рефлексов в 2 раза.
Но это у одной собаки. У другой результат был скорее обратный. Условные раздражители, примененные непосредственно после недеятельной фазы отставленного рефлекса, даже несколько увеличились в своем размере. Причина этой разницы была ясна. Когда у первой собаки между изолированной недеятельной фазой и пробой другого рефлекса ввели промежуток в 2 минуты, то в течение этих 2 минут деятельная фаза не наступила, т. е. тормозная фаза действительно была точно выделена, и поэтому последовательное торможение на другом рефлексе было налицо. У второй же собаки при тех же условиях в промежутке обнаружилась деятельная фаза и, естественно, другой рефлекс был не уменьшенным, а увеличенным. Следовательно, у этой собаки ранее и энергичнее выступил раздражительный процесс, т. e. недеятельная фаза не была полностью изолирована. С этим совпадало и то, что у первой собаки тормозной процесс вообще заметно преобладал сравнительно с раздражительным.
В прошлой и настоящей лекция привел в изобилии опыты, иллюстрирующие движение тормозного процесса по массе больших полушарий. Вам могло показаться, что это было даже и излишне. Но я сделал это с умыслом, чтобы сделать для вас очевидным, как много раз мы наблюдали это явление в более или менее простом, так сказать в чистом, виде. И все же это был только, если можно так выразиться, основной скелет явления, выступавший перед нами благодаря или особенностям экспериментирования, или индивидуальным особенностям животных, скелет, однако, наиболее часто покрываемый добавочными элементами, очень усложняющими, так сказать, полную действительность.
Первое, на что мы уже давно натолкнулись в этих опытах, состояло в следующем.
В описанных в прошлой лекции опытах (Красногорского) об иррадиирование и концентрирормозного процесса в кожном анализаторе встретился такой неожиданный тогда для нас факт. При предварительной пробе в опыте условное механическое раздражение места кожи, удаленного от тормозного пункта на 22 см, дало 6 капель слюны за 30 секунд, а раздражение зрительного анализатора дало 5 капель. Производится троекратное раздражение тормозного пункта. Минуту спустя после последнего раздражения испытывается зрительное раздражение - нуль слюноотделения, т. e. полное последовательное торможение. Через 10 минут применяется теперь четырехкратное раздражение тормозного пункта. Через минуту после него раздражение места, удаленного на 22 см, сопровождается полным эффектом - 8 каплями.
Подобный же факт наблюдался в других наших опытах (Чеботаревой). У собаки имелось несколько условных рефлексов на раздражители из разных анализаторов. Условный метрономный раздражитель, примененный вместе с условным тормозом (из зрительного анализатора), не давал ни капли слюны. 1-2 минуты спустя после того метроном один вполне восстановлял свой обычный слюноотделительный эффект и был при этом подкрепляем, а испытанные вскоре после того условные рефлексы на механическое раздражение кожи и на запах камфоры оказывались еще значительно заторможенными.
Таким образом выходило, что торможение через некоторый срок, после того как оно было произведено в данном анализаторе, уже исчезало из него, между тем в других анализаторах даже через большие сроки оно еще давало себя знать.
Но вместе с тем было обращено внимание на то, что это случалось тогда, когда условные рефлексы из других анализаторов были очень молодые, т. e. только что образованные, или хотя и старые, но давно не практиковавшиеся, или, наконец. вообще слабые, выработанные на агентов малой интенсивности. Ввиду этого можно было себе представить, что, когда известный пункт, ранее заторможенный, оказывался свободным от торможения, это еще не значило, что в его районе не было более торможения. В этом пункте раздражительный процесс, в силу определенных условий, мог брать перевес над тормозным процессом, делаться, так сказать, иммунным по отношению к окружающему его, но уже ослабленному тормозному процессу. И действительно, это постоянный факт, что рефлексы с еще недостаточно концентрированным или вообще со слабым раздеажительным процессом чрезвычайно легко подвергаются действию как внешнего, так и внутреннего торможения, между тем как сильные рефлексы остаются вне влияния тех же торможений. Об этой, так сказать, крепости или даже неприкосновенности сильных и хорошо выработанных как раздражительных, так и тормозных пунктов будет речь еще впереди.
В свое время, когда впервые наблюдались случаи, как бы противоречащие правилу иррадиирования и концентрирования торможения, были исполнены следующие нарочитые опыты (А. М. Павловой).
У собак были выработаны условные рефлексы на раздражители из разных анализаторов, причем обращалось тщательное внимание на то, чтобы все они повторялись одинаковое число раз. Когда все они достигают максимума эффекта, с одним из них комбинируется новый внешний агент. и эта комбинация обычным методом превращается в тормозную. Тормозная комбинация, а после нее ее условный раздражитель повторяются до тех пор, пока последовательное торможение комбинацией положительного раздражителя не ограничится 2 минутами, т. е. после 2 минут уже незаметно на нем. Конечно, и за этот период соблюдается требование об одинаковой практике всех рефлексов. Затем все остальные рефлексы впервые испытываются совместно c условным тормозом. Оказывается, что и эти рефлексы, за исключением одного, тоже освобождаются от торможения к концу 2 минут после применения тормозной комбинации. Исключение приходится на условный рефлекс из слабого раздражения светом, который обычно отстает по слюноотделительному эффекту от остальных рефлексов. Отсюда следует, что одна и та же степень торможения не сказывается на сильных раздражителях и отчетливо дает себя знать на слабых.
Но это только один элемент, осложняющий факт иррадиирования и концентрирования тормозного процесса. Впоследствии и постепенно мы познакомились и c другим, имеющим гораздо большее значение и еще более нарушающим простой ход иррадиирования и концентрирования. Об этом элементе мы сообщим имеющийся у нас материал в следующей лекции.
А теперь я перехожу к нашим опытам относительно движения раздражительного процесса. Этот вопрос обработан у нас в гораздо меньшей степени.
Наш первый опыт (М. К. Петровой) с иррадиированием раздражительного процесса исполнен советшенно по типу тех опытов, которыми мы начали изучение иррадиирование и концентрирования тормозного процессбак вдоль задней конечности, начиная с плюсны до таза, были расположены пять приборчиков для механического раздражения кожи, приблизительно на равных друг от друга расстояниях. Из действия самого нижнего приборчика (№ 1) у одной собаки сделан условный кислотный рефлекс, у другой - пищевой. Верхние приборчики (№ 2, 3, 4 и 5) в силу генерализации, конечно, вызывали также соответствующие рефлексы, но они обычным приемом были отдифференцированы и доведены все одинаково до нуля эффекта. Тогда было приступлено к главной задаче опытоб. Сперва испытывалось действие самого нижнего приборчика, т. е. положительный раздражитель в течение 30 секунд, причем слюноотделение за каждые 15 секунд отмечалось отдельно, a затем, как обыкновенно, присоединялся безусловный раздражитель. Спустя некоторое время нижний приборчик опять пускался в ход, но только на 15 секунд, а затем сейчас же после него начинали действовать, и тоже в течение только 15 секунд, верхние приборчики, то один, то другой в разных опытах, то более близкий, то более дальний. Получилось следующее. При смене № 1 на № 2 (ближайший) слюноотделение в течение действия этого приборчика былобыкновенно много меньше, а иногда и равно слюноотделению, наблюдавшемуся в течение 15 секунд действия приборчика № 1. При смене таким же образом № 1 на № 5, т. е. самого дальнего, слюноотделение при нем резко падало. Приборчики № 3 и 4, примененные в тождественной обстановке, дали средние цифры, приближаясь № 3 к № 2 и № 4 к № 5. В другом ряде опытов отмечалось слюноотделение не только за время действия тормозных раздражителей, но до конца отделительного периода. Тогда разница в действии близких и дальних приборчиков выступала почти без единого исключения и гораздо рельефнее. Отделительный период продолжался дольше и был гораздо значительнее по величине после действия близкого приборчика, чем дальнего.
Привожу данные ряда опытных дней.
У этой собаки кислотный условный рефлекс.

 Результат опытов очень резкий. При смене приборчика № 1 на приборчик отделение слюны во время действия (15 секунд) первого приборчика, соответственно приведенному ряду опытов: 7, 7 и 12 капель, а отделение во время действия (15 секунд) второго вместе с последовательным отделением: 15, 11 и 16; при смене же приборчика № 1 на приборчик № 5 отделение во время действия первого: 6, 8 и 8 капель, а при действии второго вместе с последовательным отделением: 3, 6 и 6 капель. Там резко больше, здесь отчетливо меньше. После действия приборчика № 2 последовательное слюноотделение продолжается 45 60 секунд, после приборчика № - только 15 секунд. А постоянное тормозящее действие приборчика № 2 должно быть и есть больше, чем приборчика 5, потому что, как мы уже знаем, чем тоньше дифференцировка, тем при полной дифференцировке на нее должен развиться более интенсивный тормозной процесс. Что действительно тормозящее действие от приборчика № 2 было больше, чем действие от приборчика № 5, в данной работе было констатировано нарочитыми опытами с последовательным торможением и от одного и от другого приборчика, притом как на приборчике № 1, так и на условных раздражителях из других анализаторов. Привожу пример последовательного торможения на приборчике № 1 через 5 минут после действия тормозящих приборчиков.
Результат опытов очень резкий. При смене приборчика № 1 на приборчик отделение слюны во время действия (15 секунд) первого приборчика, соответственно приведенному ряду опытов: 7, 7 и 12 капель, а отделение во время действия (15 секунд) второго вместе с последовательным отделением: 15, 11 и 16; при смене же приборчика № 1 на приборчик № 5 отделение во время действия первого: 6, 8 и 8 капель, а при действии второго вместе с последовательным отделением: 3, 6 и 6 капель. Там резко больше, здесь отчетливо меньше. После действия приборчика № 2 последовательное слюноотделение продолжается 45 60 секунд, после приборчика № - только 15 секунд. А постоянное тормозящее действие приборчика № 2 должно быть и есть больше, чем приборчика 5, потому что, как мы уже знаем, чем тоньше дифференцировка, тем при полной дифференцировке на нее должен развиться более интенсивный тормозной процесс. Что действительно тормозящее действие от приборчика № 2 было больше, чем действие от приборчика № 5, в данной работе было констатировано нарочитыми опытами с последовательным торможением и от одного и от другого приборчика, притом как на приборчике № 1, так и на условных раздражителях из других анализаторов. Привожу пример последовательного торможения на приборчике № 1 через 5 минут после действия тормозящих приборчиков.
 Торможение от № 2 через 5 минут его действия еще очень сильное, от № 5 через тот же срок почти или совсем незаметно.
После этого результат ряда приведенных опытов может быть истолкован только так, что процесс раздражения, вызванный действием приборчика № 1, на близком месте, соответствующем приборчику № 2, дает себя знать больше, чем на месте более дальнем, соответствующем приборчику № 5; другими словами, он распространился, иррадиировал из места его возникновения в ближайшие к нему пункты.
Эти опыты еще раз повторялись в последнее время (Подкопаевым). На одной стороне тела с передней лапы через все туловище до конца задней лапы располагаются приборчики для механического раздражения кожи. Действие прибора на передней лапе при помощи обычной процедуры становится положительным пищевым условным раздражением, действие всех остальных тормозным. Отделение слюны отмечается каждые 5 5 секунд. Один раз положительный раздражитель действует до подкрепления все 30 секунд, другой раз только первые 15 секунд, затем следует пауза в 15 секунд, только после которой и производится подкрепление. Таким образом мы имели перед собой ход отделения слюны при сплошном раздражении и при прерываемом на 15 секунд. При сплошном величина слюноотделения за первые 15 секунд относилась к величинего за вторые 15 секунд как 1 : 2.4; при прерываемом - как 1 : 1.25. В дальнейших опытах положительный раздражитель после пятнадцатисекундного действия сменялся на тормозной раздражитель, тоже пятнадцатисекундного продолжения, то при помощи ближайшего приборчика, то самого дальнего. Теперь отношение величины слюноотделения за первые 15 секунд к величине за вторые в первом случае было как 1 : 1.35, а во втором как 1 : 0. 53.
Очевидно и тут, как в опытах предшествующего сотрудника, через 0 секунд (практически через 0.5--1 секунду) по прекрашении положительного раздражения раздражительный процесс обнаруживается на ближнем тормозном пункте, уничтожая его действие, а на дальнем его нет.
Как в том, так и в этом ряде опытов мы не имеем возможности сказать, какая это фаза движения раздражителя - фаза иррадиирования или концентрирования? Идет процесс вперед или уже возвращается назад? Сейчас предпринимаются опыты, в которых тормозные раздражители применяются не сейчас же по прекращении положительного раздражителя, а спустя то или другое число секунд.
К сожалению, относительно этого вопроса приведенным исчерпывается наш теперешний нарочитый опытный материал, но у нас имеется случайно наблюденный материал, очевидно или по всей вероятности имеющий ближайшее отношение к тому же предмету.
Вот эти отдельные случаи.
В нашем распоряжении была собака чрезвычайно резко выраженным сторожевым рефлексом. Постоянно занимающийся с ней экспериментатор (М. Я. Безбокая), находясь с ней в отдельной комнате, с полным удобством снаряжал ее к опыту, ставил в станок и укреплял на ней разные приборы. С условными рефлексами дело шло вполне удовлетворительно. Но появление в экспериментальной комнате постороннего лица вызывало сильнейшую агрессивную реакцию со стороны животного, тем более, если постороннее лицо (в таком случае был я) заменяло экспериментатора перед собакой. Когда я при таком положении дела применял выработанный пищевой условный раздражитель, то слюноотделительный эффект его был значительно больше обычной величины при работе с собакой ее постоянного экспериментатора и собака ела предлагаемую пищу с чрезвычайным мускульным напряжением. Но агрессивная реакция в отношении меня совершенно прекратилась, когда я намеренно, что называется, не шелохнулся, не делал ни малейшего движения. Из всего рефлекса на меня оставалось только постоянное фиксирование меня глазами. И теперь тот же условный раздражитель давал меньше слюны, чем в норме, или. даже совсем оставался без секреторного эффекта. Но стоило мне держаться свободнее и особенно встать, чтобы сейчас же возвращались и агрессивная реакция и увеличенное слюноотделение. Интересно дальше следующее. Если в присутствии постороннего лица, сидевшего неподвижно, собака успокаивалась, то подкармливание собаки вслед за условным раздражителем вызывало на некоторое время после еды агрессивную реакцию на это лицо, хотя оно продолжало сидеть неподвижно. Едва ли эти факты можно принимать иначе, как только так. Сильное раздражение, производимое видом постороннего делающего движения лица, иррадиировало по мозгу и повысило, между прочим, возбудимость и его района, относящегося к акту еды. Когда же это раздражение при уменьшении внешнего раздражителя ослабло и концентрировалось, то, наоборот, возбудимость других отделов уменьшилась сравнительно с покойным состоянием (наше внешнее торможение). Такое же влияние вою очередь имело и раздражение едой, повысив на короткое время возбудимость центра агрессивной реакции.
Подобное же явление мы наблюдали на другой совершенно исключительной собаке (в работе И. P. Пророкова). У нее механическое раздражение кожи вызывало особенную и в высшей степени резкую двигательную реакцию (может быть, полового характера или что-либо аналогичное эффекту щекотания). Несмотря на это, на механическое раздражение кожи образовался вместе с тем и условный пищевой рефлекс. Этот пищевой рефлекс, в ярком контрасте обычным отношением, был по слюноотделению самым большим из всех остальных условных рефлексов. В данном случае обыкновенно при начале раздражения кожи появлялась особенная двигательная реакция, а 10 15 секунд спустя она сменялась энергичнейшей пищевой двигательной реакцией, какой не было при других условных раздражителях. Когда мы некоторыми приемами, о чем интересная речь будет впереди, устранили странную реакцию, то кожно-механический условный рефлекс занял по своей величине обычное место, уступая первенство звуковым раздражителям. Тоже очевидный случай иррадиации раздражительного процесса.
Таким же образом мы склонны понимать и следующий факт. При взаимодействии условных рефлексов, образованных на различные пищевые вещества, торможение одних другими наступает не сразу, а спустя несколько минут. Можно думать, что сначала имеет место иррадиация из непосредственного данным веществом раздражаемого пункта химического анализатора на соседние, чем временно поднимается их возбудимость, и лишь потом, когда раздражение в первоначальном пункте сконцентрируется, наступает торможение (опыты Савича).
С той же точки зрения можно бы объяснить и неожиданный факт, встретившийся в работе П. H. Васильева при образовании разных температурных условных рефлексов. На раздражение кожи холодом около no был выработан условный кислотный рефлекс, на тепловое раздражение около 47 - пищевой. Потребовалось большое время для образования этих рефлексов. А когда они, наконец, были готовы, они оказались в высшей степени неустойчивыми в следующем отношении. Стоило несколько раз подряд повторить и подкрепить условный кислотный холодовой рефлекс, чтобы примененныйпосле него и хорошо выработанный пищевой тепловой рефлекс превратился как бы в холодовой, и наоборот. Факт превращения был вполне отчетлив и бесспорен. Натура рефлекса определялась, во-первых, двигательной реакцией, конечно совершенно различной при этих обоих рефлексах: при пищевой собака повертывалась в сторону экспериментатора, пристально смотря то на сего, то туда, откуда появлялась еда, и облизывалась; при кислотной отворачивалась от экспериментатора, фыркала, скулила, трясла головой и делала языком выбрасывающие движения. Во-вторых, за это говорил химический характер слюны из подчелюстной слюнной железы: сильно слизистой, густой при пищевом рефлексе и жидкой, водянистой при кислотном. Мы проследили эти разницы особенно тщательно. Факт такого упорного и долгого превращения казался странным. Его было можно бы понять следующим образом. Как известно, периферические концы холодового и теплового анализаторов распределены на поверхности кожи точечно и смешанно. Можно было бы предполагать, что и центральные концы этих температурных анализаторов также тесно переплетены между собой, и из наших опытов (А. A. Iишло) мы действительно знаем, что корковые концы холодовых и тепловых нервов топографически совпадают. Если принять это во внимание, то будет понятно, почему раздражение так легко иррадиирует из холодового анализатора в тепловой, и обратно, почему так трудно достигается в наших опытах разъединение деятельности обоих анализаторов.
Теперь я должен вернуться к факту, который был уже упомянут в первой лекции об анализаторной деятельности коры и который по его механизму должен быть помещен в настоящем отделе нашего изложения. Это факт первоначальной генерализации всякого только что выработанного условного рефлекса, т. е. факт как бы самопроизвольного действия в качестве условных раздражителей таких агентов, которые сами по себе ранее не применялись, но составляют родственную группу с тем агентом, на который при помощи определенной процедуры был выработан данный условный рефлекс.
Для образца я приведу опыты над кожным анализатором (Анрепа).
На одной половине тела были расположены приборчики для механического раздражения кожи. Действие приборчика № 0, находящегося на бедре, было сделано условным пищевым раздражителем. Другие приборчики были распределены таким образом: № 1 - на конце задней конечности, № 2 - на тазу, № 3 на середине туловища, № 4 - на плече, № 5 - на предплечье и № 6 - на конце передней конечности. Действие всех этих приборчиков само по себе, т. е. без всякой предварительной вырабо ботки, также сопровождалось условным слюноотделением, причем слюноотделительный эффект чрезвычайно постепенно падал по мере удаления от № 0. В нижеприводимой табличке содержатся средние цифры, полученные на одной из трех примененных для этих опытов собак. Количества слюны выражены в делениях шкалы, где деление соответствует 0.01 куб. см. Отделение отмечалось за 30 секунд действия раздражителей.
Торможение от № 2 через 5 минут его действия еще очень сильное, от № 5 через тот же срок почти или совсем незаметно.
После этого результат ряда приведенных опытов может быть истолкован только так, что процесс раздражения, вызванный действием приборчика № 1, на близком месте, соответствующем приборчику № 2, дает себя знать больше, чем на месте более дальнем, соответствующем приборчику № 5; другими словами, он распространился, иррадиировал из места его возникновения в ближайшие к нему пункты.
Эти опыты еще раз повторялись в последнее время (Подкопаевым). На одной стороне тела с передней лапы через все туловище до конца задней лапы располагаются приборчики для механического раздражения кожи. Действие прибора на передней лапе при помощи обычной процедуры становится положительным пищевым условным раздражением, действие всех остальных тормозным. Отделение слюны отмечается каждые 5 5 секунд. Один раз положительный раздражитель действует до подкрепления все 30 секунд, другой раз только первые 15 секунд, затем следует пауза в 15 секунд, только после которой и производится подкрепление. Таким образом мы имели перед собой ход отделения слюны при сплошном раздражении и при прерываемом на 15 секунд. При сплошном величина слюноотделения за первые 15 секунд относилась к величинего за вторые 15 секунд как 1 : 2.4; при прерываемом - как 1 : 1.25. В дальнейших опытах положительный раздражитель после пятнадцатисекундного действия сменялся на тормозной раздражитель, тоже пятнадцатисекундного продолжения, то при помощи ближайшего приборчика, то самого дальнего. Теперь отношение величины слюноотделения за первые 15 секунд к величине за вторые в первом случае было как 1 : 1.35, а во втором как 1 : 0. 53.
Очевидно и тут, как в опытах предшествующего сотрудника, через 0 секунд (практически через 0.5--1 секунду) по прекрашении положительного раздражения раздражительный процесс обнаруживается на ближнем тормозном пункте, уничтожая его действие, а на дальнем его нет.
Как в том, так и в этом ряде опытов мы не имеем возможности сказать, какая это фаза движения раздражителя - фаза иррадиирования или концентрирования? Идет процесс вперед или уже возвращается назад? Сейчас предпринимаются опыты, в которых тормозные раздражители применяются не сейчас же по прекращении положительного раздражителя, а спустя то или другое число секунд.
К сожалению, относительно этого вопроса приведенным исчерпывается наш теперешний нарочитый опытный материал, но у нас имеется случайно наблюденный материал, очевидно или по всей вероятности имеющий ближайшее отношение к тому же предмету.
Вот эти отдельные случаи.
В нашем распоряжении была собака чрезвычайно резко выраженным сторожевым рефлексом. Постоянно занимающийся с ней экспериментатор (М. Я. Безбокая), находясь с ней в отдельной комнате, с полным удобством снаряжал ее к опыту, ставил в станок и укреплял на ней разные приборы. С условными рефлексами дело шло вполне удовлетворительно. Но появление в экспериментальной комнате постороннего лица вызывало сильнейшую агрессивную реакцию со стороны животного, тем более, если постороннее лицо (в таком случае был я) заменяло экспериментатора перед собакой. Когда я при таком положении дела применял выработанный пищевой условный раздражитель, то слюноотделительный эффект его был значительно больше обычной величины при работе с собакой ее постоянного экспериментатора и собака ела предлагаемую пищу с чрезвычайным мускульным напряжением. Но агрессивная реакция в отношении меня совершенно прекратилась, когда я намеренно, что называется, не шелохнулся, не делал ни малейшего движения. Из всего рефлекса на меня оставалось только постоянное фиксирование меня глазами. И теперь тот же условный раздражитель давал меньше слюны, чем в норме, или. даже совсем оставался без секреторного эффекта. Но стоило мне держаться свободнее и особенно встать, чтобы сейчас же возвращались и агрессивная реакция и увеличенное слюноотделение. Интересно дальше следующее. Если в присутствии постороннего лица, сидевшего неподвижно, собака успокаивалась, то подкармливание собаки вслед за условным раздражителем вызывало на некоторое время после еды агрессивную реакцию на это лицо, хотя оно продолжало сидеть неподвижно. Едва ли эти факты можно принимать иначе, как только так. Сильное раздражение, производимое видом постороннего делающего движения лица, иррадиировало по мозгу и повысило, между прочим, возбудимость и его района, относящегося к акту еды. Когда же это раздражение при уменьшении внешнего раздражителя ослабло и концентрировалось, то, наоборот, возбудимость других отделов уменьшилась сравнительно с покойным состоянием (наше внешнее торможение). Такое же влияние вою очередь имело и раздражение едой, повысив на короткое время возбудимость центра агрессивной реакции.
Подобное же явление мы наблюдали на другой совершенно исключительной собаке (в работе И. P. Пророкова). У нее механическое раздражение кожи вызывало особенную и в высшей степени резкую двигательную реакцию (может быть, полового характера или что-либо аналогичное эффекту щекотания). Несмотря на это, на механическое раздражение кожи образовался вместе с тем и условный пищевой рефлекс. Этот пищевой рефлекс, в ярком контрасте обычным отношением, был по слюноотделению самым большим из всех остальных условных рефлексов. В данном случае обыкновенно при начале раздражения кожи появлялась особенная двигательная реакция, а 10 15 секунд спустя она сменялась энергичнейшей пищевой двигательной реакцией, какой не было при других условных раздражителях. Когда мы некоторыми приемами, о чем интересная речь будет впереди, устранили странную реакцию, то кожно-механический условный рефлекс занял по своей величине обычное место, уступая первенство звуковым раздражителям. Тоже очевидный случай иррадиации раздражительного процесса.
Таким же образом мы склонны понимать и следующий факт. При взаимодействии условных рефлексов, образованных на различные пищевые вещества, торможение одних другими наступает не сразу, а спустя несколько минут. Можно думать, что сначала имеет место иррадиация из непосредственного данным веществом раздражаемого пункта химического анализатора на соседние, чем временно поднимается их возбудимость, и лишь потом, когда раздражение в первоначальном пункте сконцентрируется, наступает торможение (опыты Савича).
С той же точки зрения можно бы объяснить и неожиданный факт, встретившийся в работе П. H. Васильева при образовании разных температурных условных рефлексов. На раздражение кожи холодом около no был выработан условный кислотный рефлекс, на тепловое раздражение около 47 - пищевой. Потребовалось большое время для образования этих рефлексов. А когда они, наконец, были готовы, они оказались в высшей степени неустойчивыми в следующем отношении. Стоило несколько раз подряд повторить и подкрепить условный кислотный холодовой рефлекс, чтобы примененныйпосле него и хорошо выработанный пищевой тепловой рефлекс превратился как бы в холодовой, и наоборот. Факт превращения был вполне отчетлив и бесспорен. Натура рефлекса определялась, во-первых, двигательной реакцией, конечно совершенно различной при этих обоих рефлексах: при пищевой собака повертывалась в сторону экспериментатора, пристально смотря то на сего, то туда, откуда появлялась еда, и облизывалась; при кислотной отворачивалась от экспериментатора, фыркала, скулила, трясла головой и делала языком выбрасывающие движения. Во-вторых, за это говорил химический характер слюны из подчелюстной слюнной железы: сильно слизистой, густой при пищевом рефлексе и жидкой, водянистой при кислотном. Мы проследили эти разницы особенно тщательно. Факт такого упорного и долгого превращения казался странным. Его было можно бы понять следующим образом. Как известно, периферические концы холодового и теплового анализаторов распределены на поверхности кожи точечно и смешанно. Можно было бы предполагать, что и центральные концы этих температурных анализаторов также тесно переплетены между собой, и из наших опытов (А. A. Iишло) мы действительно знаем, что корковые концы холодовых и тепловых нервов топографически совпадают. Если принять это во внимание, то будет понятно, почему раздражение так легко иррадиирует из холодового анализатора в тепловой, и обратно, почему так трудно достигается в наших опытах разъединение деятельности обоих анализаторов.
Теперь я должен вернуться к факту, который был уже упомянут в первой лекции об анализаторной деятельности коры и который по его механизму должен быть помещен в настоящем отделе нашего изложения. Это факт первоначальной генерализации всякого только что выработанного условного рефлекса, т. е. факт как бы самопроизвольного действия в качестве условных раздражителей таких агентов, которые сами по себе ранее не применялись, но составляют родственную группу с тем агентом, на который при помощи определенной процедуры был выработан данный условный рефлекс.
Для образца я приведу опыты над кожным анализатором (Анрепа).
На одной половине тела были расположены приборчики для механического раздражения кожи. Действие приборчика № 0, находящегося на бедре, было сделано условным пищевым раздражителем. Другие приборчики были распределены таким образом: № 1 - на конце задней конечности, № 2 - на тазу, № 3 на середине туловища, № 4 - на плече, № 5 - на предплечье и № 6 - на конце передней конечности. Действие всех этих приборчиков само по себе, т. е. без всякой предварительной вырабо ботки, также сопровождалось условным слюноотделением, причем слюноотделительный эффект чрезвычайно постепенно падал по мере удаления от № 0. В нижеприводимой табличке содержатся средние цифры, полученные на одной из трех примененных для этих опытов собак. Количества слюны выражены в делениях шкалы, где деление соответствует 0.01 куб. см. Отделение отмечалось за 30 секунд действия раздражителей.
 Стойкость генерализации очень различна у разных собак, и удерживание ее при опытах на определенной степени не всегда легко. Если мы пробуем новые места и всякий раз раздражение подкрепляем, мы выравниваем рефлексы. Если же пробные раздражения оставляем без подкрепления, мы даем повод к развитию торможения, к образованию дифференцировок. Поэтому в этих опытах, чтобы иметь более или менее точный результат, гарантированный повторением, при повторных пробах строго поочередно один раз раздражение подкрепляют, другой раз нет, или же делают эти пробы через очень большие промежутки времени.
Как понимать механизм генерализации? Всего соответственнее нашим теперешним знаниям представлять себе это явление следующим образом. Каждому элементу рецепторного аппарата отвечают свое афферентное нерволокно и своя нервная клетка в коре полушарий, и каждой большей или меньшей группе их - свои группы волокон и клеток. Внешнее раздражение, пришедшее в кору с определенного пункта или района рецептора, не остается в соответствующих им точке или отделе коры, а иррадирует дальше по коре, и в тем более слабой степени, чем дальше отходит от центра раздражения, что мы воочию видели на другом нервном процессе, на торможении. Таким образом рядом с первым пунктом раздражения и в силу иррадиации раздражаемые пункты также должны связаться с центром безусловного рефлекса и повести, следовательно, к образованию прибавочных рефлексов, тем сравнительно более слабых, чем дальше эти пункты находятся от пункта приложения внешнего раздражителя, так как величина условного эффекта определенно связывается с силой раздражения. С такой точкой зрения согласуется и наблюденный (П. С. Купаловым) факт, что генерализация происходит, развивается в течение некоторого времени, потому что более отдаленные рефлексы как более слабые однаруживаются позже.
Объяснение, данное нами для факта первоначальной генерализации условных рефлексов в одном анализаторе, можно было бы применить и к факту универсальной генерализации, наступающей при следовых рефлексах, образованных на поздних следах раздражителя, и о которой было сообщено в седьмой лекции. При позднем присоединении безусловного раздражителя к данному внешнему агенту раздражение от него успевает расплыться по всему полушарию и потому все пункты полушария могут войти в связь с центром безусловного раздражителя. При раннем же присоединении безусловного раздражителя центр его концентрирует раздражение по направлению к себе и тем ограничивает его слишком обширную иррадиацию.
Стойкость генерализации очень различна у разных собак, и удерживание ее при опытах на определенной степени не всегда легко. Если мы пробуем новые места и всякий раз раздражение подкрепляем, мы выравниваем рефлексы. Если же пробные раздражения оставляем без подкрепления, мы даем повод к развитию торможения, к образованию дифференцировок. Поэтому в этих опытах, чтобы иметь более или менее точный результат, гарантированный повторением, при повторных пробах строго поочередно один раз раздражение подкрепляют, другой раз нет, или же делают эти пробы через очень большие промежутки времени.
Как понимать механизм генерализации? Всего соответственнее нашим теперешним знаниям представлять себе это явление следующим образом. Каждому элементу рецепторного аппарата отвечают свое афферентное нерволокно и своя нервная клетка в коре полушарий, и каждой большей или меньшей группе их - свои группы волокон и клеток. Внешнее раздражение, пришедшее в кору с определенного пункта или района рецептора, не остается в соответствующих им точке или отделе коры, а иррадирует дальше по коре, и в тем более слабой степени, чем дальше отходит от центра раздражения, что мы воочию видели на другом нервном процессе, на торможении. Таким образом рядом с первым пунктом раздражения и в силу иррадиации раздражаемые пункты также должны связаться с центром безусловного рефлекса и повести, следовательно, к образованию прибавочных рефлексов, тем сравнительно более слабых, чем дальше эти пункты находятся от пункта приложения внешнего раздражителя, так как величина условного эффекта определенно связывается с силой раздражения. С такой точкой зрения согласуется и наблюденный (П. С. Купаловым) факт, что генерализация происходит, развивается в течение некоторого времени, потому что более отдаленные рефлексы как более слабые однаруживаются позже.
Объяснение, данное нами для факта первоначальной генерализации условных рефлексов в одном анализаторе, можно было бы применить и к факту универсальной генерализации, наступающей при следовых рефлексах, образованных на поздних следах раздражителя, и о которой было сообщено в седьмой лекции. При позднем присоединении безусловного раздражителя к данному внешнему агенту раздражение от него успевает расплыться по всему полушарию и потому все пункты полушария могут войти в связь с центром безусловного раздражителя. При раннем же присоединении безусловного раздражителя центр его концентрирует раздражение по направлению к себе и тем ограничивает его слишком обширную иррадиацию.
Лекция одиннадцатая. Взаимная индукция раздражительного и тормозного процессов: а) положительная фаза индукции; 6) отрицательная фаза индукции
Мм. гг.! Первое, с чем мы встретились, обратившись к внутреннему механизму работы больших полушарий, - это было движение по массе полушарий как возбуждения, так и торможения. Второе, что гораздо позже попало в круг нашего наблюдения, а затем экспериментирования, было возбуждающее влияние одного процесса на другой как в периферии от пункта данного процесса, так и на том же месте по прекращении раздражения, вызывающего непосредственно тот или другой процесс. Это влияние, по примеру Геринга (Hering) и Шеррингтона, мы назвали индукцией. Оно взаимное, речипрокное: процесс раздражения ведет к усиленному торможению, и, обратное, торможение - к усиленному раздражению. Первое обозначается у нас словом «отрицательная фаза индукции», второе - «положительная фаза индукции», или, короче, отрицательной и положительной индукцией. Факт индукции, конечно, был давно перед нашими глазами, но, как это часто случается при исследовании, долго заслонялся для нас другой идеей - идеей о правильном строго поступательном движении нервных процессов и относился нами на счет случайных нарушающих влияний, которых так много встречается при работе над этой сложнейшей частью организма. Впервые он настойчиво напросился на наше внимание в опытах Когана, когда, как упомянуто в позапрошлой лекции, после полного угашения кожно-механического раздражения на одном месте, непосредственно испытанное раздражение отдаленного места у одной из экспериментируемых собак оказалось почти постоянно в состоянии повышенной возбудимости. Однако факт индукции оставался неразработанным вплоть до работы Фурсикова, с которой и началось нарочитое и энергичное исследование этого явления. Вот исходный опыт из работы Фурсикова. У собаки был выработан между прочими условный пищевой рефлекс на механическое раздражение кожи на передней лапе. Такое же раздражение на задней лапе было отдифференцировано, т. е. сделано тормозным, причем дифференцировка была полной - при раздражении не отделялось ни капли слюны. Мы видим, как значительно (почти на 50 %) увеличился секреторный эффект, когда положительный условный раздражитель был применен непосредственно после тормозного, причем укоротился и латентный период слюнной реакции. Кроме того, надо прибавить, что очень резко усилилась и пищевая двигательная реакция животного.
В данном случае представить себе внутренний механизм факта не представляет трудности. Пункт мозга, отвечающий месту кожи на задней лапе, в силу выработки под влиянием соответствующего раздражения, приводится в тормозное состояние. Он таким остается и после окончания и при повторении раздражения, между тем как пункт, отвечающий передней лапе, сейчас же по окончании тормозного раздражения оказывается на некоторое время в состоянии повышенной возбудимости. Здесь явные пространственные отношения дают возможность для простого объяснения. Иначе стоит дело, когда и положительный и отрицательный раздражители должны быть локализированы в одном и том же пункте мозга, как в случае дифференцирования одного и того же раздражителя по его интенсивности, по его постоянному или прерывистому характеру, по той или другой частоте перерывов. Однако факт индукции выступает и здесь с такое же отчетливостью.
Вот опыт также из работы Фурсикова. У собаки между прочими выработан условный пищевой рефлекс на удары метронома, 76 ударов в минуту. От этой частоты ударов отдифференцирована частота 186 ударов в минуту, т. е. она сопровождается нулем эффекта и секреторного и двигательного.
Мы видим, как значительно (почти на 50 %) увеличился секреторный эффект, когда положительный условный раздражитель был применен непосредственно после тормозного, причем укоротился и латентный период слюнной реакции. Кроме того, надо прибавить, что очень резко усилилась и пищевая двигательная реакция животного.
В данном случае представить себе внутренний механизм факта не представляет трудности. Пункт мозга, отвечающий месту кожи на задней лапе, в силу выработки под влиянием соответствующего раздражения, приводится в тормозное состояние. Он таким остается и после окончания и при повторении раздражения, между тем как пункт, отвечающий передней лапе, сейчас же по окончании тормозного раздражения оказывается на некоторое время в состоянии повышенной возбудимости. Здесь явные пространственные отношения дают возможность для простого объяснения. Иначе стоит дело, когда и положительный и отрицательный раздражители должны быть локализированы в одном и том же пункте мозга, как в случае дифференцирования одного и того же раздражителя по его интенсивности, по его постоянному или прерывистому характеру, по той или другой частоте перерывов. Однако факт индукции выступает и здесь с такое же отчетливостью.
Вот опыт также из работы Фурсикова. У собаки между прочими выработан условный пищевой рефлекс на удары метронома, 76 ударов в минуту. От этой частоты ударов отдифференцирована частота 186 ударов в минуту, т. е. она сопровождается нулем эффекта и секреторного и двигательного.
 Сейчас же после дифференцировки положительный раздражитель увеличился на 30%, латентный период резко укоротился, а двигательная реакция очень усилилась.
Приведу еще пример положительной индукции (из работы М. П. Калмыкова). У собаки между другими выработан условный пищевой рефлекс на сильный свет и от него отдифференцирован рефлекс на слабый свет.
Сейчас же после дифференцировки положительный раздражитель увеличился на 30%, латентный период резко укоротился, а двигательная реакция очень усилилась.
Приведу еще пример положительной индукции (из работы М. П. Калмыкова). У собаки между другими выработан условный пищевой рефлекс на сильный свет и от него отдифференцирован рефлекс на слабый свет.
 И здесь непосредственно после тормозного раздражителя действие положительного увеличилось почти на 50%, чрезвычайно резко укоротился латентный период его и отчетливо усилилась двигательная пищевая реакция.
Имея перед собой эти опыты, можно было поставить добавочный вопрос: не есть ли факт положительной индукции явление растормаживания, с которым мы так часто встречались в лекциях о разных случаях внутреннего торможения? Это предположение надо было сделать, принимая во внимание, что при положительной индукции вводится новое отношение, новая (по быстроте) смена условно-отрицательного раздражителя условноположительным, а новизна во внешнем мире может вызвать ориентировочную реакцию, а стало быть и растормаживание. Но Но, помимо многих разниц в фактических подробностях положительной индукции и растормаживания, это предположение решительно отстраняется характером двигательной реакции животного, которая с первого момента отнюдь не общая ориентировочная, а отчетливо специальнйственная данному положительному раздражителю.
Как долго держится индуцирующее действие? Срок довольно колеблющийся, от нескольких секунд до 1-2 минут. В чем лежит основание этих колебаний - не было достаточно выяснено в наших опытах.
После констатирования факта положительной индукции действительность сейчас же поставила перед нами вопрос о подробностях этого факта, так как явление индукции далеко не было постоянным. Хотя мы и доселе не овладели предметом вполне, однако некоторые условия, от которых зависит наличность индукции, более или менее наметились.
Первое условие обратило на себя наше внимание случайно. Калмыков в его опытах с условным пищевым раздражителем ударами метронома (100 ударов в минуту) --получал постоянно положительную индукцию непосредственно после применения отдифференцированной частоты ударов (160 ударов в минуту). Когда он должен был демонстировать это мне с несколькими посторонними лицами, оказалось совсем другое: не усиление, а резкое ослабление после дифференцировки. Неожиданное отклонение всего естественнее было понимать следующим образом. Я с посторонними лицами нашим разговором (собака в звуковом отношении не была достаточно изолирована от экспериментатора и нас) как агент внешнего торможения ослабил дифференцировку, вначале даже явно растормозил ее, - и явление индукции в течение всего опыта ни разу не однаружилось.
Вот этот опыт.
И здесь непосредственно после тормозного раздражителя действие положительного увеличилось почти на 50%, чрезвычайно резко укоротился латентный период его и отчетливо усилилась двигательная пищевая реакция.
Имея перед собой эти опыты, можно было поставить добавочный вопрос: не есть ли факт положительной индукции явление растормаживания, с которым мы так часто встречались в лекциях о разных случаях внутреннего торможения? Это предположение надо было сделать, принимая во внимание, что при положительной индукции вводится новое отношение, новая (по быстроте) смена условно-отрицательного раздражителя условноположительным, а новизна во внешнем мире может вызвать ориентировочную реакцию, а стало быть и растормаживание. Но Но, помимо многих разниц в фактических подробностях положительной индукции и растормаживания, это предположение решительно отстраняется характером двигательной реакции животного, которая с первого момента отнюдь не общая ориентировочная, а отчетливо специальнйственная данному положительному раздражителю.
Как долго держится индуцирующее действие? Срок довольно колеблющийся, от нескольких секунд до 1-2 минут. В чем лежит основание этих колебаний - не было достаточно выяснено в наших опытах.
После констатирования факта положительной индукции действительность сейчас же поставила перед нами вопрос о подробностях этого факта, так как явление индукции далеко не было постоянным. Хотя мы и доселе не овладели предметом вполне, однако некоторые условия, от которых зависит наличность индукции, более или менее наметились.
Первое условие обратило на себя наше внимание случайно. Калмыков в его опытах с условным пищевым раздражителем ударами метронома (100 ударов в минуту) --получал постоянно положительную индукцию непосредственно после применения отдифференцированной частоты ударов (160 ударов в минуту). Когда он должен был демонстировать это мне с несколькими посторонними лицами, оказалось совсем другое: не усиление, а резкое ослабление после дифференцировки. Неожиданное отклонение всего естественнее было понимать следующим образом. Я с посторонними лицами нашим разговором (собака в звуковом отношении не была достаточно изолирована от экспериментатора и нас) как агент внешнего торможения ослабил дифференцировку, вначале даже явно растормозил ее, - и явление индукции в течение всего опыта ни разу не однаружилось.
Вот этот опыт.
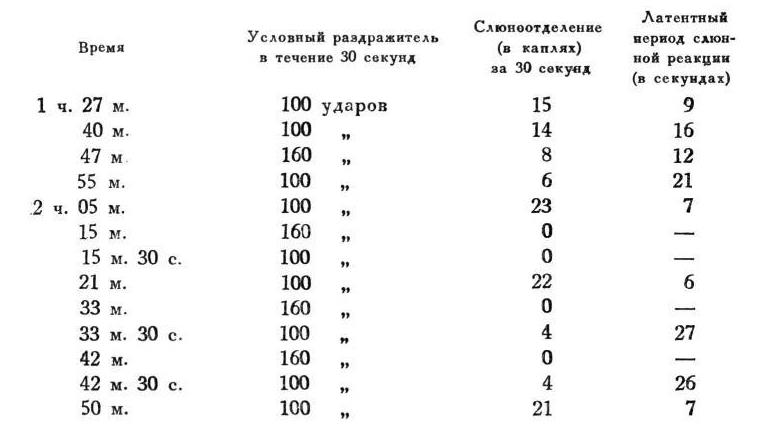 Как видим, дифференцировка, бывшая до этого опыта полной, при первом применении в нашем присутствии нарушилась, дала 4 капли. Дальше она стала снова нулевой, но все разы в опыте после нее положительный рефлекс непосредственно не увеличивался, а уменьшался, т. е. оказывался последовательно заторможенным.
Из этого можно было заключить, что ослабление тормозного процесса повлекло за собой исчезание индукции. Чтобы проверить этот вывод, тормозной процесс, лежащий в основании дифференцировки, был ослаблен другим способом, именно предварительным введением в рот собаки отвергаемых веществ. После этого также в течение всего опыта ни разу не наблюдалась индукция (тоже опыты Калмыкова). Следовательно, появление положительной индукции связано с определенной интенсивностью тормозного процесса.
Второе условие, от которого зависит наличие индукции, выступило перед нами в той же работе Калмыкова. Ради особенной цели, имея дифференцировку на 160 ударов метронома в минуту, мы применяли ее в течение нескольких месяцев большое число раз, не испытывая ее на индуцирующее действие, а затем перешли к выработке более тонкой дифференцировки, именно на 112 ударов в минуту, т. е. ближе к частоте положительного раздражителя (100 ударов в минуту). Когда последняя сделалась полной, проба на индукцию дала очень резкий результат.
Вот этот опыт.
Как видим, дифференцировка, бывшая до этого опыта полной, при первом применении в нашем присутствии нарушилась, дала 4 капли. Дальше она стала снова нулевой, но все разы в опыте после нее положительный рефлекс непосредственно не увеличивался, а уменьшался, т. е. оказывался последовательно заторможенным.
Из этого можно было заключить, что ослабление тормозного процесса повлекло за собой исчезание индукции. Чтобы проверить этот вывод, тормозной процесс, лежащий в основании дифференцировки, был ослаблен другим способом, именно предварительным введением в рот собаки отвергаемых веществ. После этого также в течение всего опыта ни разу не наблюдалась индукция (тоже опыты Калмыкова). Следовательно, появление положительной индукции связано с определенной интенсивностью тормозного процесса.
Второе условие, от которого зависит наличие индукции, выступило перед нами в той же работе Калмыкова. Ради особенной цели, имея дифференцировку на 160 ударов метронома в минуту, мы применяли ее в течение нескольких месяцев большое число раз, не испытывая ее на индуцирующее действие, а затем перешли к выработке более тонкой дифференцировки, именно на 112 ударов в минуту, т. е. ближе к частоте положительного раздражителя (100 ударов в минуту). Когда последняя сделалась полной, проба на индукцию дала очень резкий результат.
Вот этот опыт.
 Индуцирующее действие новой, более тонкой дифференцировки достигло 77% с очень значительным укорочением латентного периода.
Но когда мы теперь испытали старую, грубую дифференцировку, мы, к немалому нашему удивлению, не могли ни разу видеть от нее и следа индукции. Предполагая, что, может быть, в этом имел значение перерыв в применении грубой дифференцировки, когда вырабатывалась новая, более тонкая, мы затем чередовали периоды в 10-15 дней, в один период практикуя только одну дифференцировку, в другой - только другую. Однако это нисколько не изменило факта, как показывают следующие опыты.
Индуцирующее действие новой, более тонкой дифференцировки достигло 77% с очень значительным укорочением латентного периода.
Но когда мы теперь испытали старую, грубую дифференцировку, мы, к немалому нашему удивлению, не могли ни разу видеть от нее и следа индукции. Предполагая, что, может быть, в этом имел значение перерыв в применении грубой дифференцировки, когда вырабатывалась новая, более тонкая, мы затем чередовали периоды в 10-15 дней, в один период практикуя только одну дифференцировку, в другой - только другую. Однако это нисколько не изменило факта, как показывают следующие опыты.
 Новая дифференцировка явно положительно индуцирует, старая влечет за собой прямо последовательное торможение. Но с повторением новой дифференцировки и ее положительно индуцирующее действие стало заметно уменьшаться.
Такие же опыты, только с другим раздражителем, были повторены еще раз (Фроловым). При этом имелось в виду, помимо применения другого раздражителя, ввести разницу еще и в том отношении, чтобы дифференцируемый тормозной агент был большей интенсивности, чем положительный раздражитель. Был взят тон D тонвариатора Макса Коля и заглушен в разной степени. При опытах пользовались тремя интенсивностями: самый слабый тон, средний и наиболее сильный. Из самого слабого был выработан условный пищевой раздражитель. Затем от него сперва был отдифференцирован наиболее сильный. Когда он становился вполне отрицательным, его пробовали на индуцируюшее действие.
Вот первый опыт с индукцией.
Новая дифференцировка явно положительно индуцирует, старая влечет за собой прямо последовательное торможение. Но с повторением новой дифференцировки и ее положительно индуцирующее действие стало заметно уменьшаться.
Такие же опыты, только с другим раздражителем, были повторены еще раз (Фроловым). При этом имелось в виду, помимо применения другого раздражителя, ввести разницу еще и в том отношении, чтобы дифференцируемый тормозной агент был большей интенсивности, чем положительный раздражитель. Был взят тон D тонвариатора Макса Коля и заглушен в разной степени. При опытах пользовались тремя интенсивностями: самый слабый тон, средний и наиболее сильный. Из самого слабого был выработан условный пищевой раздражитель. Затем от него сперва был отдифференцирован наиболее сильный. Когда он становился вполне отрицательным, его пробовали на индуцируюшее действие.
Вот первый опыт с индукцией.

 При непосредственном переходе от дифференцированного тона к положительному последний оказался увеличенным на 50 %. В течение месяца с лишком дифференцировка постоянно практиковалась. Испробованная затем, она уже более не действовала индуцирующим образом.
Вот этот опыт.
При непосредственном переходе от дифференцированного тона к положительному последний оказался увеличенным на 50 %. В течение месяца с лишком дифференцировка постоянно практиковалась. Испробованная затем, она уже более не действовала индуцирующим образом.
Вот этот опыт.
 После этого было приступлено к выработке более тонкой дифференцировки на средний тон, которая была готова к девятнадцатому разу ее применения и сейчас же испробована на индукцию.
Вот этот опыт.
После этого было приступлено к выработке более тонкой дифференцировки на средний тон, которая была готова к девятнадцатому разу ее применения и сейчас же испробована на индукцию.
Вот этот опыт.
 Совершенно так же, как в предшествующих опытах (Калмыкова), старая и более грубая дифференцировка, повторяемая много раз, потеряла свое индуцирующее действие, а новая и более тонкая, будучи вполне образована, также начала с хорошо выраженной индукции.
На основании произведенных опытов надо заключить, что положительная индукция представляет собой временное, фазовое явление при установке новых отношений в нервной деятельности, являясь после полного развития тормозного процесса и исчезая после его основательного упрочнения. Так ли это всегда? В некоторых случаях индукция держалась очень долго и мы не видали ее исчезания. Сколько можно судить по теперешнему нашему материалу, эта разница могла приурочиваться к следующему обстоятельству: происходит ли эта быстрая смена тормозного процесса раздражительным в одном и том же пункте мозга, как в только что приведенных опытах, или же процессы разделены пространственно, как в опытах с кожным анализатором. Вопрос должен быть разрешен новыми опытами, которые сейчас и производятся.
Надо прибавить еще, что нами делались опыты - и неоднократно - с дифференцированием частоты ритмических механических раздражений кожи на одном том же месте. В этих случаях нам до сих пор ни разу не пришлось видеть положительной индукции, когда мы быстро, без промежутка, действующую как условный положительный раздражитель частоту применяли после частоты, связанной с тормозным процессом. Всегда прямо наступал период последовательного торможения.
Факт отрицательной фазы индукции, т. е. усиление тормозного процесса под влиянием предшествующего раздражительного, встретился нам также уже давно, но оставался для нас как что-то совершенно непонятное и даже долго не был снова испробован. Лишь в относительно недавнее время мы, наконец, заинтересовались им и повторили его много раз на разных собаках, анализыровав и оценив его надлежащим образом (опыт Строганова).
При изучении внутреннего торможения мы, между прочим, исследовали вопрос: как происходит устранение, разрушение выработанного тормозного процесса? Опыт (Кржышковского) был поставлен специально над случаем условного торможения. Тон был сделан положительным кислотным раздражителем, а комбинация его с кожно-механическим раздражением была превращена в тормозную обычным способом, т. е. тон один постоянно сопровождался вливанием кислоты, а комбинация, чередуясь с положительным раздражителем, повторялась без вливания кислоты. Когда решено было разрушить тормозное действие комбинации, то процедуру разрушения сначала воспроизвели точно по типу процедуры образования, т. е. чередуя подкрепляемый положительный раздражитель с комбинацией, тоже теперь сопровождаемой вливанием кислоты. Получился довольно неожиданный результат. Несмотря на десятикратное, в течение трех дней, применение комбинации с вливанием кислоты, комбинация не получила положительного действия, оставаясь нулевой.
Вот эти опыты.
Совершенно так же, как в предшествующих опытах (Калмыкова), старая и более грубая дифференцировка, повторяемая много раз, потеряла свое индуцирующее действие, а новая и более тонкая, будучи вполне образована, также начала с хорошо выраженной индукции.
На основании произведенных опытов надо заключить, что положительная индукция представляет собой временное, фазовое явление при установке новых отношений в нервной деятельности, являясь после полного развития тормозного процесса и исчезая после его основательного упрочнения. Так ли это всегда? В некоторых случаях индукция держалась очень долго и мы не видали ее исчезания. Сколько можно судить по теперешнему нашему материалу, эта разница могла приурочиваться к следующему обстоятельству: происходит ли эта быстрая смена тормозного процесса раздражительным в одном и том же пункте мозга, как в только что приведенных опытах, или же процессы разделены пространственно, как в опытах с кожным анализатором. Вопрос должен быть разрешен новыми опытами, которые сейчас и производятся.
Надо прибавить еще, что нами делались опыты - и неоднократно - с дифференцированием частоты ритмических механических раздражений кожи на одном том же месте. В этих случаях нам до сих пор ни разу не пришлось видеть положительной индукции, когда мы быстро, без промежутка, действующую как условный положительный раздражитель частоту применяли после частоты, связанной с тормозным процессом. Всегда прямо наступал период последовательного торможения.
Факт отрицательной фазы индукции, т. е. усиление тормозного процесса под влиянием предшествующего раздражительного, встретился нам также уже давно, но оставался для нас как что-то совершенно непонятное и даже долго не был снова испробован. Лишь в относительно недавнее время мы, наконец, заинтересовались им и повторили его много раз на разных собаках, анализыровав и оценив его надлежащим образом (опыт Строганова).
При изучении внутреннего торможения мы, между прочим, исследовали вопрос: как происходит устранение, разрушение выработанного тормозного процесса? Опыт (Кржышковского) был поставлен специально над случаем условного торможения. Тон был сделан положительным кислотным раздражителем, а комбинация его с кожно-механическим раздражением была превращена в тормозную обычным способом, т. е. тон один постоянно сопровождался вливанием кислоты, а комбинация, чередуясь с положительным раздражителем, повторялась без вливания кислоты. Когда решено было разрушить тормозное действие комбинации, то процедуру разрушения сначала воспроизвели точно по типу процедуры образования, т. е. чередуя подкрепляемый положительный раздражитель с комбинацией, тоже теперь сопровождаемой вливанием кислоты. Получился довольно неожиданный результат. Несмотря на десятикратное, в течение трех дней, применение комбинации с вливанием кислоты, комбинация не получила положительного действия, оставаясь нулевой.
Вот эти опыты.

 Не получив разрушения таким способом, мы на следующий день поступили иначе: стали комбинацию, сопровождаемую вливанием кислоты, повторять сплошь, не чередуя с применением условного положительного раздражителя. Теперь разрушение началось и происходило стремительно.
Не получив разрушения таким способом, мы на следующий день поступили иначе: стали комбинацию, сопровождаемую вливанием кислоты, повторять сплошь, не чередуя с применением условного положительного раздражителя. Теперь разрушение началось и происходило стремительно.
 Казалось, что разница в этих двух способах разрушения очевидна, однако сейчас же возникло предположение: не было ли разрушение, так скоро начавшееся и быстро протекшее при втором способе, собственно подготовлено применением первого способа. Чтобы проверить это предположение, тормозная комбинация была восстановлена, практиковалась в течение одного года и была испытана вновь на разрушение прямо по второму способу. Получилось следующее.
Казалось, что разница в этих двух способах разрушения очевидна, однако сейчас же возникло предположение: не было ли разрушение, так скоро начавшееся и быстро протекшее при втором способе, собственно подготовлено применением первого способа. Чтобы проверить это предположение, тормозная комбинация была восстановлена, практиковалась в течение одного года и была испытана вновь на разрушение прямо по второму способу. Получилось следующее.
 Теперь стало ясно, что оба способа разрушения существенно различаются, но, как сказано выше, факт только спустя много лет сделался опять предметом нашего исследования. В позднейшее время исследование было произведено на четырех собаках (опыты (от Строганова). Условные рефлексы на этот раз были пищевые. Разрушалось дифференцировочное торможение, именно дифференцировки на частоту ударов метронома и на высоту тонов. Факт резкой разницы в быстроте разрушения тормозного процесса посредством двух способов повторился без исключения на всех собаках. При сплошном, одно за другим, применении дифференцировочного раздражителя, сопровождаемого безусловным раздражителем, разрушение тормозного процесса начиналось через один или вообще немного раз; при правильном чередовании этой комбинации с условным положительным раздражителем разрушение сильно затягивалось во времени, наступая через много раз, даже через десятки раз. На разных животных первым употреблялся то один, то другой способ разрушения. Иногда многократно чередовались эти способы на одном и том же животном. Конечно, в таких случаях дифференцировки после разрушения основательно восстановлялись. Наконец было введено несколько вариантов опыта. Вот они.
Когда дифференцировочное торможение при сплошном разрушении почти исчезло, стоило ввести один раз постоянный положительный раздражитель, чтобы прежний тормозной раздражитель восстановился в большей или меньшей и даже в полной мере.
У собаки 120 ударов метронома в минуту положительный условный пищевой раздражитель, 60 ударов - дифференцировка, при испытании 41 раз в течение сорокадневного периода не дававшая ни капли слюны.
Теперь стало ясно, что оба способа разрушения существенно различаются, но, как сказано выше, факт только спустя много лет сделался опять предметом нашего исследования. В позднейшее время исследование было произведено на четырех собаках (опыты (от Строганова). Условные рефлексы на этот раз были пищевые. Разрушалось дифференцировочное торможение, именно дифференцировки на частоту ударов метронома и на высоту тонов. Факт резкой разницы в быстроте разрушения тормозного процесса посредством двух способов повторился без исключения на всех собаках. При сплошном, одно за другим, применении дифференцировочного раздражителя, сопровождаемого безусловным раздражителем, разрушение тормозного процесса начиналось через один или вообще немного раз; при правильном чередовании этой комбинации с условным положительным раздражителем разрушение сильно затягивалось во времени, наступая через много раз, даже через десятки раз. На разных животных первым употреблялся то один, то другой способ разрушения. Иногда многократно чередовались эти способы на одном и том же животном. Конечно, в таких случаях дифференцировки после разрушения основательно восстановлялись. Наконец было введено несколько вариантов опыта. Вот они.
Когда дифференцировочное торможение при сплошном разрушении почти исчезло, стоило ввести один раз постоянный положительный раздражитель, чтобы прежний тормозной раздражитель восстановился в большей или меньшей и даже в полной мере.
У собаки 120 ударов метронома в минуту положительный условный пищевой раздражитель, 60 ударов - дифференцировка, при испытании 41 раз в течение сорокадневного периода не дававшая ни капли слюны.
 Другой вариант состоял в том, что в опыте прямо перед началом сплошного разрушения дифференцировки положительный раздражитель повторялся три раза. Это обусловливало необычно позднее наступление разрушения (на пятом-шестом разе).
У другой собаки те же положительный и отрицательный условные раздражители, отрицате(дифференцировка) даже менее стойкий, легко подвергавшийся растормаживанию.
Другой вариант состоял в том, что в опыте прямо перед началом сплошного разрушения дифференцировки положительный раздражитель повторялся три раза. Это обусловливало необычно позднее наступление разрушения (на пятом-шестом разе).
У другой собаки те же положительный и отрицательный условные раздражители, отрицате(дифференцировка) даже менее стойкий, легко подвергавшийся растормаживанию.
 Третий вариант такой. Разрушение производилось по способу чередования. Когда разрушение началось, и положительное действие дифференцированного раздражителя делалось постоянным, хотя оставалось еще незначительным, то четырехкратное применение подряд постоянного положительного раздражителя непосредственно уничтожало это действие.
Кроме описанной формы опыта для обнаружения отрицательной фазы индукции, мы применяли другую (опыты Пророкова). У многих собак с пищевыми условными рефлексами второй в опыте рефлекс часто бывает наибольший по слюноотделительному эффекту, очевидно вследствие повышенной пищевой возбудимости после первого рефлекторного раздражения из полости рта. Поэтому уже выработанная, но еще не сделавшаяся прочной дифференцировка, будучи поставлена в опыте на втором месте, т. е. после первого подкармливания, нередко нарушается, делается неполной, в известной мере растормаживается. Если имеется несколько положительных условных пищевых рефлексов и при одном из них - дифференцировка, то наблюдается следующее отношение. Если примененный первом в данном опыте условный раздражитель без дифференцировки, то дифференцировка другого раздражителя, поставленная на втором месте, часто и значительно растормаживается. В случае же, когда опыт начинается с положительного раздражителя, имеющего дифференцировку, то она, испытанная на втором месте, нарушается очень мало, редко и в незначительной степени Вот подлинный опыт: 144 удара метронома в минуту - положительный условный раздражитель, 72 удара - дифференцировка. Другой пищевой раздражитель - звонок. Когда опыт начинался со звонка, метрономная дифференцировка, поставленная после него, растормозилась в 8 случаях из 12 и растормаживание доходило в максимуме до 72%. 0/ Дифференцировка, примененная после положительного метронома, была нарушена только в двух случаях из двенадцати, и нарушение не шло дальше 20 %. Конечно, отдельные опыты этих двух серий чередовались, так что результат не мог зависеть от различного состояния дифференцируемого раздражителя вследствие все большего его упрочнения с дальнейшим его применением. Этот результат вместе с тем показывает, что индуцирование, по крайней мере иногда, специально существует между данным условным раздражителем и его дифференцировкой, другой же условный раздражитель, даже из того же анализатора, индуцирующего действия на ту же диф- фФеренцировку не оказывает.
Обе приведенные формы опыта делают очевидным, что раздражительный процесс при известных условиях благоприятствует, усиливает тормозной процесс, тe. что мы имеем перед собой отрицательную фазу индукции.
После ознакомления с этой фазой естественно было предположить, что и то, что мы ранее назвали внешним торможением условных рефлексов, есть тоже явление отрицательной индукции. Раздражительный процесс в одном пункте мозга индуцирует тормозное состояние в остальных в большей или меньшей степени. Но в данном случае вопрос осложнялся. Надо было специально выяснить: есть ли явление внешнего торможения корковое явление, в коре происходящее, или же это торможение имеет место только в низших инстанциях мозга, так как в случае внешнего торможения участвуют центры двух безусловных рефлексов. Что во внешнем торможении условных рефлексов должно давать себя знать взаимодействие в виде торможения центров безусловных рефлексов, надо принять на основании многочисленных опытов над низшими отделами центральной нервной системы. И это торможение, конечно, можно бы представлять себе в виде отрицательной индукции. Но здесь нам требуется доказать, что то же относится и к взаимодействию различных пунктов именно коры в случае внешнего торможения. Это доказательство представить не легко. У нас делались следующие опыты (Фурсиковым). Образовывались условные рефлексы на два безусловных раздражителя: пищевой и разрушительный (электрический ток, приложенный к коже). Как только на условный разрушительный раздражитель начиналась оборонительная реакция, сейчас же присоединялся условный пищевой раздражитель. большинстве случаев пищевой условный рефлекс теперь оказывался заторможенным в большей или меньшей степени, иногда и вполне. Но раз оборонительная реакция была налицо, значит раздражался безусловный центр этой реакции. Следовательно, и в таких опытах должно было участвовать взаимодействие безусловных центров. Однако в этих опытах была подробность, которая давала основание заключить, что и между пунктами условных раздражителей, т. е. в самой коре, существовало взаимодействие всле отрицательной индукции. Если условные раздражители пищевого и оборонительного рефлексов принадлежали к одному и тому же анализатору, например звуковому, то образование условного оборонительного рефлекса часто вело за собой падение величины условного пищевого рефлекса. В противоположном случае, когда условные раздражители обоих рефлексов были взяты из разных анализаторов, то условный пищевой рефлекс нисколько не терял в величине своего эффекта после образования условного оборонительного рефлекса. Больше того. Если в одном и том же, именно кожном, анализаторе к разным пунктам приурочены и пищевой и разрушительный раздражители, то наблюдается следующее. Так как в опытах от условного пункта оборонительной реакции на разных расстояниях располагались два или несколько пунктов пищевой реакции, то оказывалось, что после образования условного оборонительного рефлекса постоянно терял в своей величине только пищевой рефлекс с ближайшего пункта к пункту оборонительного рефлекса, а рефлекс с дальнего пункта оставался без изменения. При этом надо заметить, что ближайший пищевой пункт нисколько не обнаруживал свойств пункта оборонительной реакции, никаких оборонительных движений не примешивалось к пищевому рефлексу. Если бы внешнее торможение имело место только между центрами безусловных рефлексов, то таких отношений в анализаторах быть не могло.
Следовательно, наше так называемое внешнее торможение существует и между разными пунктами коры больших полушарий. А раз так, то это торможение вполне подходило бы под формулу отрицательной индукции и должно рассматриваться как тождественное с внутренним торможением. В настоящее время мы воспроизводим такие формы опыта, которые, вероятно, дадут более прямое доказательство того положения, что по сущности процесса, т. е. в отношении физико-химического субстрата, внешнее и внутреннее торможения представляют одно и то же физиологические явление. Может быть, даже к концу наших лекций эти опыты приведут к совершенно определенному результату.
Ясно, что описанные явления взаимной индукции вполне совпадают с большой группой контрастных явлений, изученных в теперешней физиологии органов чувств, и еще раз показывают, как объективное изучение на животных успешно захватывает то, что ранее считалось доступным только субъективному изучению.
Третий вариант такой. Разрушение производилось по способу чередования. Когда разрушение началось, и положительное действие дифференцированного раздражителя делалось постоянным, хотя оставалось еще незначительным, то четырехкратное применение подряд постоянного положительного раздражителя непосредственно уничтожало это действие.
Кроме описанной формы опыта для обнаружения отрицательной фазы индукции, мы применяли другую (опыты Пророкова). У многих собак с пищевыми условными рефлексами второй в опыте рефлекс часто бывает наибольший по слюноотделительному эффекту, очевидно вследствие повышенной пищевой возбудимости после первого рефлекторного раздражения из полости рта. Поэтому уже выработанная, но еще не сделавшаяся прочной дифференцировка, будучи поставлена в опыте на втором месте, т. е. после первого подкармливания, нередко нарушается, делается неполной, в известной мере растормаживается. Если имеется несколько положительных условных пищевых рефлексов и при одном из них - дифференцировка, то наблюдается следующее отношение. Если примененный первом в данном опыте условный раздражитель без дифференцировки, то дифференцировка другого раздражителя, поставленная на втором месте, часто и значительно растормаживается. В случае же, когда опыт начинается с положительного раздражителя, имеющего дифференцировку, то она, испытанная на втором месте, нарушается очень мало, редко и в незначительной степени Вот подлинный опыт: 144 удара метронома в минуту - положительный условный раздражитель, 72 удара - дифференцировка. Другой пищевой раздражитель - звонок. Когда опыт начинался со звонка, метрономная дифференцировка, поставленная после него, растормозилась в 8 случаях из 12 и растормаживание доходило в максимуме до 72%. 0/ Дифференцировка, примененная после положительного метронома, была нарушена только в двух случаях из двенадцати, и нарушение не шло дальше 20 %. Конечно, отдельные опыты этих двух серий чередовались, так что результат не мог зависеть от различного состояния дифференцируемого раздражителя вследствие все большего его упрочнения с дальнейшим его применением. Этот результат вместе с тем показывает, что индуцирование, по крайней мере иногда, специально существует между данным условным раздражителем и его дифференцировкой, другой же условный раздражитель, даже из того же анализатора, индуцирующего действия на ту же диф- фФеренцировку не оказывает.
Обе приведенные формы опыта делают очевидным, что раздражительный процесс при известных условиях благоприятствует, усиливает тормозной процесс, тe. что мы имеем перед собой отрицательную фазу индукции.
После ознакомления с этой фазой естественно было предположить, что и то, что мы ранее назвали внешним торможением условных рефлексов, есть тоже явление отрицательной индукции. Раздражительный процесс в одном пункте мозга индуцирует тормозное состояние в остальных в большей или меньшей степени. Но в данном случае вопрос осложнялся. Надо было специально выяснить: есть ли явление внешнего торможения корковое явление, в коре происходящее, или же это торможение имеет место только в низших инстанциях мозга, так как в случае внешнего торможения участвуют центры двух безусловных рефлексов. Что во внешнем торможении условных рефлексов должно давать себя знать взаимодействие в виде торможения центров безусловных рефлексов, надо принять на основании многочисленных опытов над низшими отделами центральной нервной системы. И это торможение, конечно, можно бы представлять себе в виде отрицательной индукции. Но здесь нам требуется доказать, что то же относится и к взаимодействию различных пунктов именно коры в случае внешнего торможения. Это доказательство представить не легко. У нас делались следующие опыты (Фурсиковым). Образовывались условные рефлексы на два безусловных раздражителя: пищевой и разрушительный (электрический ток, приложенный к коже). Как только на условный разрушительный раздражитель начиналась оборонительная реакция, сейчас же присоединялся условный пищевой раздражитель. большинстве случаев пищевой условный рефлекс теперь оказывался заторможенным в большей или меньшей степени, иногда и вполне. Но раз оборонительная реакция была налицо, значит раздражался безусловный центр этой реакции. Следовательно, и в таких опытах должно было участвовать взаимодействие безусловных центров. Однако в этих опытах была подробность, которая давала основание заключить, что и между пунктами условных раздражителей, т. е. в самой коре, существовало взаимодействие всле отрицательной индукции. Если условные раздражители пищевого и оборонительного рефлексов принадлежали к одному и тому же анализатору, например звуковому, то образование условного оборонительного рефлекса часто вело за собой падение величины условного пищевого рефлекса. В противоположном случае, когда условные раздражители обоих рефлексов были взяты из разных анализаторов, то условный пищевой рефлекс нисколько не терял в величине своего эффекта после образования условного оборонительного рефлекса. Больше того. Если в одном и том же, именно кожном, анализаторе к разным пунктам приурочены и пищевой и разрушительный раздражители, то наблюдается следующее. Так как в опытах от условного пункта оборонительной реакции на разных расстояниях располагались два или несколько пунктов пищевой реакции, то оказывалось, что после образования условного оборонительного рефлекса постоянно терял в своей величине только пищевой рефлекс с ближайшего пункта к пункту оборонительного рефлекса, а рефлекс с дальнего пункта оставался без изменения. При этом надо заметить, что ближайший пищевой пункт нисколько не обнаруживал свойств пункта оборонительной реакции, никаких оборонительных движений не примешивалось к пищевому рефлексу. Если бы внешнее торможение имело место только между центрами безусловных рефлексов, то таких отношений в анализаторах быть не могло.
Следовательно, наше так называемое внешнее торможение существует и между разными пунктами коры больших полушарий. А раз так, то это торможение вполне подходило бы под формулу отрицательной индукции и должно рассматриваться как тождественное с внутренним торможением. В настоящее время мы воспроизводим такие формы опыта, которые, вероятно, дадут более прямое доказательство того положения, что по сущности процесса, т. е. в отношении физико-химического субстрата, внешнее и внутреннее торможения представляют одно и то же физиологические явление. Может быть, даже к концу наших лекций эти опыты приведут к совершенно определенному результату.
Ясно, что описанные явления взаимной индукции вполне совпадают с большой группой контрастных явлений, изученных в теперешней физиологии органов чувств, и еще раз показывают, как объективное изучение на животных успешно захватывает то, что ранее считалось доступным только субъективному изучению.
Лекция двенадцатая. Комбинирование явлений движения нервных процессов с явлениями их взаимной индукции
Мм. гг.! В последних трех лекциях мы познакомились сперва с явлениями иррадиирования и концентрирования нервных процессов в коре больших полушарий, а потом с явлениями взаимной индукции этих процессов. Эти явления были описаны нами отдельно, как бы вне зависимости друг от друга. В действительности же, как это ясно и по смыслу дела, они должны встречаться вместе, комбинироваться, взаимодействовать. Воспроизведение этих явлений в нашем прежнем изложении разрозненными вернее отнести к более редким случаям в работе больших полушарий, приуроченным или к определенным фазам развития и состояния нервных процессов, или к особенным типам нервной системы у наших экспериментальных животных. Иногда же, вероятно, мы сами искусственно упрощали явления иррадиирования и концентрирования процессов, не имея сначала идеи об их взаимной индукции, как это уже упомянуто в начале предыдущей лекции. В начале нашей работы, когда перед нами стояла подавляющая хаотическая сложность явлений, которые мы, однако, решились изучить, приходилось умышленно закрывать глаза на многое, даже прямо устраняться от особенно трудных случаев, заменяя, например, одних собак другими. Теперь уже не то. Многолетний опыт принес свои дары, обнаружил свою силу. Теперь всякая особенность, неожиданность, представляемая тем или другим животным, привлекает наше специальное внимание, делается для нас интересным вопросом, новой задачей. Комбинирование явлений иррадиирования и концентрирования нервных процессов с явлениями их взаимной индукции представляет собой очень сложный процесс, полного знания которого придется еще долго ждать. Сейчас на эту тему мы только накопляем большей частью отрывочный, почти совершенно не систематизированный материал, к передаче которого я и подхожу теперь. Начну с наиболее простого случая (опыты Е. М. Крепса). У собаки условным раздражителем служит механическое раздражение кожи. Раздражения мест кожи - двух на бедре (первый и второй пункты), по одному на животе (третий пункт), на груди (четвертый) и на плече (пятый) - были положительными условными (безусловный рефлекс - пищевой); раздражение на голени - тормозное, дифференцировка. Дифференцировка полная, положительные раздражения выравнены в отношении величины эффекта. В начале каждогота устанавливается размер слюноотделительной реакции для положительного раздражения. Затем применяется тормозное раздражение. По прекращении его или непосредственно, или через разные промежутки времени испытываются разные положительные раздражения. Такие опыты продолжались в течение 5 месяцев. Результаты их представлены в виде двух таблиц, на которых цифры обозначают величину эффекта положительных раздражений в процентах по отношению к величине эффекта этих раздражений до применения в опыте дифференцировки.
 Первый вертикальный столбец указывает номере мест положительных раздражений, верхний горизонтальный - промежутки времени между концом тормозного и началом положительных раздражений. Первая таблица представляет средние цифры из всех опытов за все время работы, вторая - средние цифры только за последний месяц работы.
Отсутствие в таблице 1 цифр за пяти- и пятнадцатисекундный промежутки имеет свое основание в том, что эти промежутки применялись в опытах только за последний месяц работы.
Основной результат опытов заключается, как вы видите, в том, что непосредственно по прекращении тормозного раздражения эффект положительных раздражений оказывается увеличенным, к 15 секундам он уже спускается ниже нормы, а к 30 секундам достигает максимума падения и, наконец, только к пятой минуте снова представляет нормальную величину. Таким образом картина последействия тормозного раздражения начинается с положительной индукции, а затем только ваступает иррадиирование торможения, кончающееся восстановлением нормы эффекта положительного раздражения. Ty же смену положительной индукции последовательным торможением вы могли заметить и в протоколах опытов, приведеных в предшествующей лекции и констатировавших факт положительной индукции. Далее в изучаемых сейчас опытах обращают на себя внимание еще следующие подробности. Размер положительной индукции несколько возрос к позднему сроку работы. Наоборот, последовательное торможение уменьшилось с течением опытов и во времени, заканчиваясь к 2 минутам, и пространстве, ограничиваясь двумя положительными, ближайшими к тормозному. пунктами. Последнее есть повторение факта, так часто встречалшегося нам в ранних лекциях при изучении разных случаев внутреннего торможения.
Другой, также довольно простой случай представился нам на угасательном торможении (опыты Подкопаева).
В лекции об иррадиировании торможения было упомянуто наблюдение Когана, что у одной из его собак непосредственно после полного угашения кожно-механического условного раздражения на одном пункте, на самом дальнем от этого пункта постоянно замечалось повышение возбудимости - положительная индукция. Теперь этот факт в более подробном виде повторился на собаке, у которой был прослежен результат однократного угашения (неподкрепления) условного кожно-механического пищевого рефлекса. У нее были выработаны условные пищевые рефлексы на механическое раздражение нескольких мест (8) (8) кожи одной половины тела, расположенных по линии от нижней части передней конечности через все туловище до нижней части задней конечности. Все места в отношении величины эффекта раздражения были выровнены. Четыре места из них находились между собой на расстояниях 1, 43 и 89 см. На каждом из них и производилось угашение условного рефлекса. Опыт начинался с пробы того или другого из этих мест ради установки нормы рефлекса. Затем на каком-либо из них угашали рефлекс, и теперь через промежутки времени в 45 секунд, 1, 3, 4 и 8 минут снова испытывали место, с раздражения которого начинался опыт. Опыт кончался раздражением безразлично какого-либо из всех 8 мест для проверки еще раз нормы положительного эффекта данного дня. Опыты с угощением ставились через 4-5 дней в расчете иметь более или менее стационарное положение торможения. Но этот расчет не оправдался. Торможение тем не менее все более и более концентрировалось в течение опытов, и, таким образом, общий результат надо было проследить в трех последовательных сериях, на трех таблицаx. Собака, на которой делались эти опыты, отличалась таким постоянством величины положительного условного эффекта, что можно было пользоваться каждым отдельным опытом, а не средним из нескольких опытов. В таблицах приведены величины эффекта в процентах по отнонению к норме каждого дня. В таблице 5 прибавлены промежутки в 45 секунд, 1 и 3 минуты, потому что только с повторением опытов слюноотделение настолько быстро стало прекращаться, что получилась возможность все раньше испытывать последействие угашения.
Первый вертикальный столбец указывает номере мест положительных раздражений, верхний горизонтальный - промежутки времени между концом тормозного и началом положительных раздражений. Первая таблица представляет средние цифры из всех опытов за все время работы, вторая - средние цифры только за последний месяц работы.
Отсутствие в таблице 1 цифр за пяти- и пятнадцатисекундный промежутки имеет свое основание в том, что эти промежутки применялись в опытах только за последний месяц работы.
Основной результат опытов заключается, как вы видите, в том, что непосредственно по прекращении тормозного раздражения эффект положительных раздражений оказывается увеличенным, к 15 секундам он уже спускается ниже нормы, а к 30 секундам достигает максимума падения и, наконец, только к пятой минуте снова представляет нормальную величину. Таким образом картина последействия тормозного раздражения начинается с положительной индукции, а затем только ваступает иррадиирование торможения, кончающееся восстановлением нормы эффекта положительного раздражения. Ty же смену положительной индукции последовательным торможением вы могли заметить и в протоколах опытов, приведеных в предшествующей лекции и констатировавших факт положительной индукции. Далее в изучаемых сейчас опытах обращают на себя внимание еще следующие подробности. Размер положительной индукции несколько возрос к позднему сроку работы. Наоборот, последовательное торможение уменьшилось с течением опытов и во времени, заканчиваясь к 2 минутам, и пространстве, ограничиваясь двумя положительными, ближайшими к тормозному. пунктами. Последнее есть повторение факта, так часто встречалшегося нам в ранних лекциях при изучении разных случаев внутреннего торможения.
Другой, также довольно простой случай представился нам на угасательном торможении (опыты Подкопаева).
В лекции об иррадиировании торможения было упомянуто наблюдение Когана, что у одной из его собак непосредственно после полного угашения кожно-механического условного раздражения на одном пункте, на самом дальнем от этого пункта постоянно замечалось повышение возбудимости - положительная индукция. Теперь этот факт в более подробном виде повторился на собаке, у которой был прослежен результат однократного угашения (неподкрепления) условного кожно-механического пищевого рефлекса. У нее были выработаны условные пищевые рефлексы на механическое раздражение нескольких мест (8) (8) кожи одной половины тела, расположенных по линии от нижней части передней конечности через все туловище до нижней части задней конечности. Все места в отношении величины эффекта раздражения были выровнены. Четыре места из них находились между собой на расстояниях 1, 43 и 89 см. На каждом из них и производилось угашение условного рефлекса. Опыт начинался с пробы того или другого из этих мест ради установки нормы рефлекса. Затем на каком-либо из них угашали рефлекс, и теперь через промежутки времени в 45 секунд, 1, 3, 4 и 8 минут снова испытывали место, с раздражения которого начинался опыт. Опыт кончался раздражением безразлично какого-либо из всех 8 мест для проверки еще раз нормы положительного эффекта данного дня. Опыты с угощением ставились через 4-5 дней в расчете иметь более или менее стационарное положение торможения. Но этот расчет не оправдался. Торможение тем не менее все более и более концентрировалось в течение опытов, и, таким образом, общий результат надо было проследить в трех последовательных сериях, на трех таблицаx. Собака, на которой делались эти опыты, отличалась таким постоянством величины положительного условного эффекта, что можно было пользоваться каждым отдельным опытом, а не средним из нескольких опытов. В таблицах приведены величины эффекта в процентах по отнонению к норме каждого дня. В таблице 5 прибавлены промежутки в 45 секунд, 1 и 3 минуты, потому что только с повторением опытов слюноотделение настолько быстро стало прекращаться, что получилась возможность все раньше испытывать последействие угашения.
 В этих таблицах мы видим, что в начале опытов (таблица 3) торможение простиралось до самого дальнего пункта и еще было отчетливо спустя 12 минут после угашения. Затем (таблица 4) торможение ограничивалось уже расстоянием в 43 см и было через 4 минуты меньше того, что было в предшествующей фазе опытов через 12 минут. Теперь на дальнем пункте и в более позднее время начинает встречаться, хотя и редко, вместо уменьшения, увеличение эффекта сравнительно с нормальной величиной. Наконец в последней фазе (таблица торможение с колебаниями во времени держится только в первично угашаемом месте. На остальных пунктах, за исключением дальнего, в течение минуты даже на 1 см расстояния и к концу минуты после угашения или преобладает увеличение эффекта против норм, или устанавливается норма.
Перед нами - положительная индукция, сперва возникающая поодаль от исходного пункта торможения и обнаруживающаяся более поздно, а затем постепенно подвигающаяся по направлению к исходному пункту и все быстрее появляющаяся после начала торможения, т. е. как бы постепенно преодолевающая и вытесняющая и во времени и в пространстве тормозной процесс.
Только что приведенные опыты интересны и своими подробностями. Во-первых, обращает на себя внимание высочайшая чувствительность элементов коры, когда однократное неподкрепление сказывается на большой территории коры и держится значительное время (более 12 минут). Во-вторых, перед нами лишнее свидетельство текучести, изменяемости явлений в работе больших полушарии, когда незначительное влияние, повторяемое через длинные сроки, в 4 -5 дней, резко меняет общее положение дела (быстрая смена фаз). Наконец нельзя не заметить, что состояние разных пунктов полушарий представляет ясную волнообразность как во времени, так и в пространстве. Например, в таблице 5 первично угашаемый пункт в течение 12 минут оказывался то заторможенным, то свободным от торможения; то же можно видеть и в пространстве на разных пунктах мозга в одно и то же время. Это капитальный факт, с которым мы будем в дальнейшем встречаться все чаще чаще. Он является совершенно естественным результатом встречи, уравновешивания двух противоположных нервных процессов, раздражения и торможения, результатом, совершенно аналогичным, например, волнам третьего рода кривой кровяного давления как результату взаимодействия прессорных и депрессорных иннерваций.
Та же волнообразность выступает на той же собаке (также в опытах Подкопаева) и при гашении до нуля и именно на пункте, ближайшем к угашаемому. На дальнем пункте она не была заметна. Может быть, за ее слабое выражение надо считать в соответствующем периоде времени остающийся в течение 7 минут без изменения уровень торможения.
Вот этот опыт.
В этих таблицах мы видим, что в начале опытов (таблица 3) торможение простиралось до самого дальнего пункта и еще было отчетливо спустя 12 минут после угашения. Затем (таблица 4) торможение ограничивалось уже расстоянием в 43 см и было через 4 минуты меньше того, что было в предшествующей фазе опытов через 12 минут. Теперь на дальнем пункте и в более позднее время начинает встречаться, хотя и редко, вместо уменьшения, увеличение эффекта сравнительно с нормальной величиной. Наконец в последней фазе (таблица торможение с колебаниями во времени держится только в первично угашаемом месте. На остальных пунктах, за исключением дальнего, в течение минуты даже на 1 см расстояния и к концу минуты после угашения или преобладает увеличение эффекта против норм, или устанавливается норма.
Перед нами - положительная индукция, сперва возникающая поодаль от исходного пункта торможения и обнаруживающаяся более поздно, а затем постепенно подвигающаяся по направлению к исходному пункту и все быстрее появляющаяся после начала торможения, т. е. как бы постепенно преодолевающая и вытесняющая и во времени и в пространстве тормозной процесс.
Только что приведенные опыты интересны и своими подробностями. Во-первых, обращает на себя внимание высочайшая чувствительность элементов коры, когда однократное неподкрепление сказывается на большой территории коры и держится значительное время (более 12 минут). Во-вторых, перед нами лишнее свидетельство текучести, изменяемости явлений в работе больших полушарии, когда незначительное влияние, повторяемое через длинные сроки, в 4 -5 дней, резко меняет общее положение дела (быстрая смена фаз). Наконец нельзя не заметить, что состояние разных пунктов полушарий представляет ясную волнообразность как во времени, так и в пространстве. Например, в таблице 5 первично угашаемый пункт в течение 12 минут оказывался то заторможенным, то свободным от торможения; то же можно видеть и в пространстве на разных пунктах мозга в одно и то же время. Это капитальный факт, с которым мы будем в дальнейшем встречаться все чаще чаще. Он является совершенно естественным результатом встречи, уравновешивания двух противоположных нервных процессов, раздражения и торможения, результатом, совершенно аналогичным, например, волнам третьего рода кривой кровяного давления как результату взаимодействия прессорных и депрессорных иннерваций.
Та же волнообразность выступает на той же собаке (также в опытах Подкопаева) и при гашении до нуля и именно на пункте, ближайшем к угашаемому. На дальнем пункте она не была заметна. Может быть, за ее слабое выражение надо считать в соответствующем периоде времени остающийся в течение 7 минут без изменения уровень торможения.
Вот этот опыт.
 Торможение вычислено в процентах остающегося нормального положительного эффекта.
Волнообразность и вообще еще более сложное отношения между раздражением и торможением на разных пунктах кожного анализатора и при разных паузах после применения тормозного раздражения обнаружились на следующем нашем животном (опыты Андреева). По длине задней конечности с верхней трети бедра до нижней части голени прикреплялись приборчики для механического раздражения кожи, числом четыре, № 0, 1, 2 и 3, считая сверху вниз. Расстояние между ними приблизительно одинаковое, около 15 см. Раздражения тремя нижними приборчиками в отдельности были положительными условными пищевыми раздражениями, а раздражение верхним - тормозным, дифференцировкой. Дифференцировка не была особенно прочной. Общий ход опыта был таков. Паузы между раздражениями были постоянно семиминутные. Опыт начинался с условного раздражения звонком или ударами метронома. Затем применялось два положительных раздражения приборчиками, безразлично какими, за ними следовало тоже положительное кожномеханическое раздражение, специально испытуемое в данном экспериментальном сеансе. Теперь приводился в действие верхний приборчик, производивший торможение, а после него, на этот раз через разные паузы, от 0 до 12--16 минут, снова применялось кожно-механическое раздражение, последнее перед дифференцировкой, и на нем определялось влияние дифференцировки. Это влияние вычислялось в процентах по отношению к нормальной величине эффекта положительного кожно-механического раздражения, принимаемой за 100. За нормальную величину принималась средняя величина эффекта трех кожномеханических раздражений, предшествующих дифференцировке.
Вот числовой результат опытов.
Торможение вычислено в процентах остающегося нормального положительного эффекта.
Волнообразность и вообще еще более сложное отношения между раздражением и торможением на разных пунктах кожного анализатора и при разных паузах после применения тормозного раздражения обнаружились на следующем нашем животном (опыты Андреева). По длине задней конечности с верхней трети бедра до нижней части голени прикреплялись приборчики для механического раздражения кожи, числом четыре, № 0, 1, 2 и 3, считая сверху вниз. Расстояние между ними приблизительно одинаковое, около 15 см. Раздражения тремя нижними приборчиками в отдельности были положительными условными пищевыми раздражениями, а раздражение верхним - тормозным, дифференцировкой. Дифференцировка не была особенно прочной. Общий ход опыта был таков. Паузы между раздражениями были постоянно семиминутные. Опыт начинался с условного раздражения звонком или ударами метронома. Затем применялось два положительных раздражения приборчиками, безразлично какими, за ними следовало тоже положительное кожномеханическое раздражение, специально испытуемое в данном экспериментальном сеансе. Теперь приводился в действие верхний приборчик, производивший торможение, а после него, на этот раз через разные паузы, от 0 до 12--16 минут, снова применялось кожно-механическое раздражение, последнее перед дифференцировкой, и на нем определялось влияние дифференцировки. Это влияние вычислялось в процентах по отношению к нормальной величине эффекта положительного кожно-механического раздражения, принимаемой за 100. За нормальную величину принималась средняя величина эффекта трех кожномеханических раздражений, предшествующих дифференцировке.
Вот числовой результат опытов.
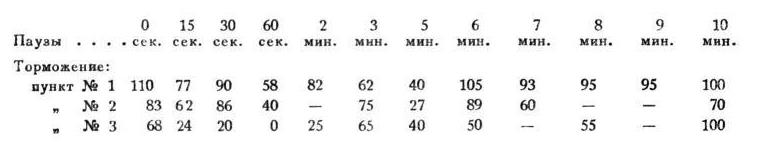 Для более легкого и быстрого представления об оказавшейся в этих опытах сложности отношений тот же результат воспроизвожу в виде кривых. Как видите, влияние тормозного процесса, производимого раздражением на пункте 0, на всех положительных пунктах оказывалось гораздо более различным, чем сходным. Самое общее сходство состояло в том, что торможение рано или поздно достигало максимума, а затем исчезало. Затем оно сказалось и в том, что на всех этих пунктах в одно и то же время обнаружились два максимума торможения: первый к концу первой минуты по прекращении тормозного раздражения, второй к концу пятой минуты. На этом сходство кончилось, а потом выступилн разницы, и значительные. На первом, ближайшем к тормозному пункте сейчас же по прекращении тормозного раздражения было небольшое повышение возбудимости положительная индукция. На двух остальных пунктах ее не было, началось прямо с торможения. На первом и втором пунктах второй максимум торможения был больше первого, на третьем пункте первый максимум представлялся полным торможением, совершенным исчезанием положительного эффекта, второй обозначился слабо. Затем на первом и втором пунктах происходило много колебаний, и очень резких. На третьем пункте было только довольно значительное ослабление торможения на третьей минуте. Без него ход последовательного торможения был бы такой же регулярный, как мы познакомились с ним раньше - в девятой и десятой лекциях. На этом третьем пункте опять так же, как это мы видели раньше, период достижения максимума был в несколько раз короче периода полного исчезания торможения; на первом же и втором пунктах период достижения наибольшего максимума был или равен или даже меньше периода полного возвращения нормальной возбудимости. Наконец если взять валовое последовательное торможение, т. е. если сложить все проценты торможения, обозначенные в таблице, то оказется,что торможение всего более дало себя знать на третьем пункте, менее на втором пункте и всего меньше на первом пункте.
Для более легкого и быстрого представления об оказавшейся в этих опытах сложности отношений тот же результат воспроизвожу в виде кривых. Как видите, влияние тормозного процесса, производимого раздражением на пункте 0, на всех положительных пунктах оказывалось гораздо более различным, чем сходным. Самое общее сходство состояло в том, что торможение рано или поздно достигало максимума, а затем исчезало. Затем оно сказалось и в том, что на всех этих пунктах в одно и то же время обнаружились два максимума торможения: первый к концу первой минуты по прекращении тормозного раздражения, второй к концу пятой минуты. На этом сходство кончилось, а потом выступилн разницы, и значительные. На первом, ближайшем к тормозному пункте сейчас же по прекращении тормозного раздражения было небольшое повышение возбудимости положительная индукция. На двух остальных пунктах ее не было, началось прямо с торможения. На первом и втором пунктах второй максимум торможения был больше первого, на третьем пункте первый максимум представлялся полным торможением, совершенным исчезанием положительного эффекта, второй обозначился слабо. Затем на первом и втором пунктах происходило много колебаний, и очень резких. На третьем пункте было только довольно значительное ослабление торможения на третьей минуте. Без него ход последовательного торможения был бы такой же регулярный, как мы познакомились с ним раньше - в девятой и десятой лекциях. На этом третьем пункте опять так же, как это мы видели раньше, период достижения максимума был в несколько раз короче периода полного исчезания торможения; на первом же и втором пунктах период достижения наибольшего максимума был или равен или даже меньше периода полного возвращения нормальной возбудимости. Наконец если взять валовое последовательное торможение, т. е. если сложить все проценты торможения, обозначенные в таблице, то оказется,что торможение всего более дало себя знать на третьем пункте, менее на втором пункте и всего меньше на первом пункте.
 Эту сложность в распределении возбудимости на разных пунктах кожного анализатора под влиянием тормозного процесса всего естественнее было бы понять так, что индуцированное повышение возбудимости дало себя знать всего больше около пункта, где внешним раздражением было вызвано торможение, и потому здесь сильнее выступили волны борьбы, уравновешивания двух противоположных процессов. А что здесь действительно индуцированный раздражительный процесс был всего значительнее, доказывается фактом явной положительной индукции, которая только на этом пункте и наблюдалась сейчас же по прекращении тормозного раздражения. На дальнем пункте торможение обнаружилось наиболее беспрепятственно, проделав свое обыкновенное и постепенное усиление до максимума, а затем более растянутое во времени ослабление, до возвращения норма. ьного эффекта.
С методической стороны в этом случае надо, однако, указать, что отдельные цифры, из которых были получены приведенные средние цифры, относящиеся к ближнему пункту, расходились между собой гораздо больше, чем цифры, относящиеся к среднему и в особенности к дальнему пунктам. Может быть, отчасти на это следует отнести менее значительные разницы в кривых разных пунктов. Но, может быть, это самое непостоянство отдельных цифр обусловливалось именно большими колебаниями возбудимости на ближнем пункте, чем на дальнем.
Наконец я приведу еще опыты (Подкопаева), опять же на новом животном, которые так же резко иодчеркнут большую сложность разбираемых отношений.
У собаки на одной половине тела по одной линии расположены приборчики для механического раздражения кожи на запястье (№ 0), на плече (№ 1). на груди (№ 2), на животе (№ 3) и на бедре (№ 4). На запястье выработан положительчый условный пищевой рефлекс, на остальных - тормозные, между собой выровненные. После действия каждого из тормозящих приборчиков непосредственно, т. е. без всякого промежутка времени, испытывается положительно С начала опыта определяется его нормальный эффект. Опыты действием разных тормозящих приборчиков на эффект положительно действующего ставились беспорядочно то с одним, то С другим, так что полученный результат отнести на счет фазовых изменений с течением времени нет оснований.
В прилагаемой таблице приведены как отдельные опыты, так и среднее из них. Эффект положительно действующего приборчика вычислен в процентах по отношению к его нормальной величине.
Эту сложность в распределении возбудимости на разных пунктах кожного анализатора под влиянием тормозного процесса всего естественнее было бы понять так, что индуцированное повышение возбудимости дало себя знать всего больше около пункта, где внешним раздражением было вызвано торможение, и потому здесь сильнее выступили волны борьбы, уравновешивания двух противоположных процессов. А что здесь действительно индуцированный раздражительный процесс был всего значительнее, доказывается фактом явной положительной индукции, которая только на этом пункте и наблюдалась сейчас же по прекращении тормозного раздражения. На дальнем пункте торможение обнаружилось наиболее беспрепятственно, проделав свое обыкновенное и постепенное усиление до максимума, а затем более растянутое во времени ослабление, до возвращения норма. ьного эффекта.
С методической стороны в этом случае надо, однако, указать, что отдельные цифры, из которых были получены приведенные средние цифры, относящиеся к ближнему пункту, расходились между собой гораздо больше, чем цифры, относящиеся к среднему и в особенности к дальнему пунктам. Может быть, отчасти на это следует отнести менее значительные разницы в кривых разных пунктов. Но, может быть, это самое непостоянство отдельных цифр обусловливалось именно большими колебаниями возбудимости на ближнем пункте, чем на дальнем.
Наконец я приведу еще опыты (Подкопаева), опять же на новом животном, которые так же резко иодчеркнут большую сложность разбираемых отношений.
У собаки на одной половине тела по одной линии расположены приборчики для механического раздражения кожи на запястье (№ 0), на плече (№ 1). на груди (№ 2), на животе (№ 3) и на бедре (№ 4). На запястье выработан положительчый условный пищевой рефлекс, на остальных - тормозные, между собой выровненные. После действия каждого из тормозящих приборчиков непосредственно, т. е. без всякого промежутка времени, испытывается положительно С начала опыта определяется его нормальный эффект. Опыты действием разных тормозящих приборчиков на эффект положительно действующего ставились беспорядочно то с одним, то С другим, так что полученный результат отнести на счет фазовых изменений с течением времени нет оснований.
В прилагаемой таблице приведены как отдельные опыты, так и среднее из них. Эффект положительно действующего приборчика вычислен в процентах по отношению к его нормальной величине.
 Из таблицы ясно, что состояние положительно действующего пункта очень различно в зависимости от того, действие какого из тормозящих приборчиков ему предшествовало. При раздражении ближайшего и отдаленнейшего пункта наступала резкая положительная индукция: значительно увеличивалось слюноотделение сравнительно с нормой, укорачивался латентный период и часто усиливалась пищевая двигательная реакция, причем индуцирующее действие дальнего пункта было несколько больше, чем ближнего. От третьего пункта не наблюдалось никакого действия, а от второго - чрезвычайно смешанное: тормозящее, положительно индуцирующее и никакого. Сходство отдельных цифр при повторении тождественных опытов в трех рядах так велико, что нет основания заподозрить их точность, и это же располагает отнестись с доверием и к цифрам, относящимся к второму пункту, считая, что он представляет какую-то переходную зону.
Как понимать этот результат? Если считать, что линейное расположение наших пунктов на коже соответствует такому же расположению проекционных пунктов в коре, то представленный результат надо считать ярким доказательством волнообразного состояния возбудимости в пространстве. Мы должны будем признать, что на пути движения того или другого нервного процесса перемежаются районы то с повышенной, то с пониженной возбудимостью, с преобладанием то раздражительного процесса, то тормозного. Ввиду важности этого вывода необходимо иметь специальные доказательства, что принимаемая нами соответственность расположения пунктов на коже и в коре есть точный факт.
Весь приведенный в текущей лекции материал более или менее согласно свидетельствует, что изменения возбудимости различных пунктов коры под влиянием внешних раздражений, специально под влиянием тормозного раздражения, происходят волнообразно как во времени на одном том же пункте, так и в пространстве на разных пунктах в одно и то же время. И это не должно казаться неожиданностью, раз мы имеем несомненные факты, с одной стороны, движения нервных процессов, с другой - их взаимной индукции. Остается только большим вопросом общее точное правило, по которому это происходит: почему один раз выступает волнообразность, в другой раз ее нет, почему в одних случаях распространяющееся торможение предшествуется положительной индукцией, в других ее не видно, и т. д.? Сейчас мы стоим перед беспорядочным разнообразием фактов. Но имеющийся в наших руках материал позволяет заключить, что это разнообразие обусловливает соучастие трех факторов. Это особые типы нервной системы наших экспериментальных животных, фазы установки новых отношений в коре под влиянием внешних раздражений и виды установки, так как, например, наши случаи внутреннего торможения очень различаются между собой по их напряженности и стойкости. Серьезная задача будущего полного анализа исследуемых отношений и заключается в точном определении участия в данный момент деятельности коры каждого из указанных факторов, сведя все на силовые отношения нервных процессов. Постепенное приближение к этой задаче, конечно, возможно в настоящее время. Уже в описанных опытах можно указать на некоторые намечающиеся частичные подходы к решению нашей задачи. Но еще отчетливее выступает это в опытах, которыми мы занимались и занимаемся в последнее время. Мы видели постоянно в последних четырех лекциях, что торможение, производимое соответственным раздражением в определенном пункте, начинает прямо или после предварительной кратковременной положительное индукции распространяться по анализатору лишь по прекращении тормозного раздражения. Почему так? Как создается такое положение дела? Ради ответа на этот вопрос мы поставили, очевидно до сих пор недостававшие в нашем плане исследования предмета, опыты.
Мы испытывали другие пункты анализатора или другие анализаторы не по прекращении тормоздражения, как всегда делали это раньше, а во время продолжающегося тормозного раздражения, на фоне его, как выражался Подкопаев, первый разрабатывавший этот вопрос. Испытание было произведено на четырех различных собаках. Прежде всего опишу опыты (Подкопаева) с угасательным торможением.
Это та же собака, которая служила для приведенных в текущей лекции опытов с иррадиацией торможения после однократного неподкрепления условного рефлекса и после угашения до нуля. В описываемых теперь опытах угашение условного механического раздражения на определенном месте кожи было полное. Раздражение других пунктов производилось или на ближайшем к угашенному пункте (1 см расстояния), или на дальнем (89 см расстояния). Это присоединение происходило на тридцатой секунде продолжающегося тормозного раздражения, причем совместное раздражение также продолжалось 30 секунд. На ближайшем пункте эффект такого раздражения был 84.5 нормального, на дальнем 88%, т. е. одинаковый, в то время как по прекращении тормозного раздражения через 0 секунд промежутка, как это мы видели раньше, раздражение ближайшего и дальнего пунктов соответственно давало 44.5 и 66.5%.
Для полной оценки этого результата надо еще прибавить, что латентный период рефлексов при одновременном раздражении угашенного пункта с другими пунктами был значительно короче, чем в норме, а также и двигательная пищевая реакция резче, чем обыкновенно. И то и другое - бесспорные указания на повышение возбудимости, положительное индуцирование остальных пунктов анализатора, кроме специально угашенного. Несколько меньший слюноотделительный эффект от остальных пунктов, очевидно, надо понимать как результат алгебраического суммирования их эффекта с эффектом из угашенного пункта.
Особенный интерес для нас в этих опытах представляет еще следующее. Угашение до нуля на определенном месте кожи производилось повторением неподкрепляемого условного раздражения через 2 минуты. В новой вариации опытов по получении первого нуля процедура угашения продолжалась, и к третьему нулевому раздражению присоединялось раздражение других пунктов кожи. Теперь эффект рефлекса от соединенного раздражения был большей частью совершенно равен нормальному эффекту изолированного положительного раздражения. Значит, углубление торможения, усиление тормозного процесса поведо к более значительной положительной индукции. Рядом с этим в тех же опытах обращает на себя внимание и следующее сопоставление фактов. У этой собаки, как мы видели, к 2 минутам после угашения до нуля иррадиированное торможение на кожном анализаторе еще держится на уровне не ниже 50% 0/ полного торможения, между тем как при совместном раздражении угашаемого пункта в третий раз до нуля (через 2 минуты после предшествующего угашения) и остальных пунктов не видно ни малейшего знака торможения. Следовательно, оно во время повторного тормозного раздражения опять сконцентрировалось в пункте этого раздражения.
В той же работе (Подкопаева) те же результаты были получены на другой собаке, у которой были, между прочим, выработаны положительные пищевые условные рефлексы на определенный тон и удары метронома (120 ударов в минуту) и тормозной (дифференцировка на 60 ударов метронома в минуту). На «фоне» дифференцировочного торможения от метронома тон из 9 проб только один раз дал мало заторможенный эффект, в 4 случаях - нормальную величину его и еще в 4 - величину, даже значительно превосходившую норму.
Для следующих опытов, относящихся к той же теме, были употреблены две собаки. У одной (опыты В. П. Головиной) среди многих пищевых условных рефлексов были положительные: на метроном, определенный тон, свисток, лампочку, вспыхивающую в определенном месте, и механическое раздражение кожи, и три тормозных дифференцировки: на удары метронома другой частоты, на другое место кожи и на другое место вспыхивающей лампочки. Когда все эти рефлексы были хорошо выработаны, применялось то или другое положительное раздражение совместно с каким-нибудь из указанных тормозных раздражений. Все эти пробы дали совершенно однообразный результат. Положительные раздражители в этих комбинациях всегда имели или нормальный для них по величине эффект, или несколько увеличенный, и более часто с укороченным латентным периодом.
У другой собаки (опыты Павловой) также среди многих условных рефлексов были положительные: на свисток, звук бульканья и механическое раздражение кожи, и тормозной дифференцировка на удары метронома. В опытах над этим животным каждое из указанных положительных раздражений по нескольку раз применялось с одним и тем же тормозным раздражением. Здесь результат получился несколько иной. Хотя латентный период также большей частью был укорочен, но величина эффекта комбинированного раздражения почти постоянно была меньше, и часто значительно, сравнительно с величиной эффекта одиночного положительного раздражения. Значит, у этой собаки концентрирование нервных процессов было менее совершенным.
Нужно прибавить, что эти две последние собаки явно принадлежали к двум разным типам нервной системы. В то время как у первой дифференцировки были вообще очень прочны и не страдали от комбинированных раздражений, у второй дифференцировка часто сама по себе была неполной и всегда еще более нарушалась после комбинации положительными раздражениями.
Опыты последнего ряда ясно показывают, с одной стороны, как положительная индукция, возникающая под влиянием тормозного раздражения определеннкта, ограничивает распространение тормозного процесса, с другой - свидетельствуют, что взаимодействие между раздражительным и тормозным процессами определяется то фазой установки новых отношений в коре (опыт Подкопаева), то типом нервной системы разных животных (опыты Головиной и Павловой).
Из таблицы ясно, что состояние положительно действующего пункта очень различно в зависимости от того, действие какого из тормозящих приборчиков ему предшествовало. При раздражении ближайшего и отдаленнейшего пункта наступала резкая положительная индукция: значительно увеличивалось слюноотделение сравнительно с нормой, укорачивался латентный период и часто усиливалась пищевая двигательная реакция, причем индуцирующее действие дальнего пункта было несколько больше, чем ближнего. От третьего пункта не наблюдалось никакого действия, а от второго - чрезвычайно смешанное: тормозящее, положительно индуцирующее и никакого. Сходство отдельных цифр при повторении тождественных опытов в трех рядах так велико, что нет основания заподозрить их точность, и это же располагает отнестись с доверием и к цифрам, относящимся к второму пункту, считая, что он представляет какую-то переходную зону.
Как понимать этот результат? Если считать, что линейное расположение наших пунктов на коже соответствует такому же расположению проекционных пунктов в коре, то представленный результат надо считать ярким доказательством волнообразного состояния возбудимости в пространстве. Мы должны будем признать, что на пути движения того или другого нервного процесса перемежаются районы то с повышенной, то с пониженной возбудимостью, с преобладанием то раздражительного процесса, то тормозного. Ввиду важности этого вывода необходимо иметь специальные доказательства, что принимаемая нами соответственность расположения пунктов на коже и в коре есть точный факт.
Весь приведенный в текущей лекции материал более или менее согласно свидетельствует, что изменения возбудимости различных пунктов коры под влиянием внешних раздражений, специально под влиянием тормозного раздражения, происходят волнообразно как во времени на одном том же пункте, так и в пространстве на разных пунктах в одно и то же время. И это не должно казаться неожиданностью, раз мы имеем несомненные факты, с одной стороны, движения нервных процессов, с другой - их взаимной индукции. Остается только большим вопросом общее точное правило, по которому это происходит: почему один раз выступает волнообразность, в другой раз ее нет, почему в одних случаях распространяющееся торможение предшествуется положительной индукцией, в других ее не видно, и т. д.? Сейчас мы стоим перед беспорядочным разнообразием фактов. Но имеющийся в наших руках материал позволяет заключить, что это разнообразие обусловливает соучастие трех факторов. Это особые типы нервной системы наших экспериментальных животных, фазы установки новых отношений в коре под влиянием внешних раздражений и виды установки, так как, например, наши случаи внутреннего торможения очень различаются между собой по их напряженности и стойкости. Серьезная задача будущего полного анализа исследуемых отношений и заключается в точном определении участия в данный момент деятельности коры каждого из указанных факторов, сведя все на силовые отношения нервных процессов. Постепенное приближение к этой задаче, конечно, возможно в настоящее время. Уже в описанных опытах можно указать на некоторые намечающиеся частичные подходы к решению нашей задачи. Но еще отчетливее выступает это в опытах, которыми мы занимались и занимаемся в последнее время. Мы видели постоянно в последних четырех лекциях, что торможение, производимое соответственным раздражением в определенном пункте, начинает прямо или после предварительной кратковременной положительное индукции распространяться по анализатору лишь по прекращении тормозного раздражения. Почему так? Как создается такое положение дела? Ради ответа на этот вопрос мы поставили, очевидно до сих пор недостававшие в нашем плане исследования предмета, опыты.
Мы испытывали другие пункты анализатора или другие анализаторы не по прекращении тормоздражения, как всегда делали это раньше, а во время продолжающегося тормозного раздражения, на фоне его, как выражался Подкопаев, первый разрабатывавший этот вопрос. Испытание было произведено на четырех различных собаках. Прежде всего опишу опыты (Подкопаева) с угасательным торможением.
Это та же собака, которая служила для приведенных в текущей лекции опытов с иррадиацией торможения после однократного неподкрепления условного рефлекса и после угашения до нуля. В описываемых теперь опытах угашение условного механического раздражения на определенном месте кожи было полное. Раздражение других пунктов производилось или на ближайшем к угашенному пункте (1 см расстояния), или на дальнем (89 см расстояния). Это присоединение происходило на тридцатой секунде продолжающегося тормозного раздражения, причем совместное раздражение также продолжалось 30 секунд. На ближайшем пункте эффект такого раздражения был 84.5 нормального, на дальнем 88%, т. е. одинаковый, в то время как по прекращении тормозного раздражения через 0 секунд промежутка, как это мы видели раньше, раздражение ближайшего и дальнего пунктов соответственно давало 44.5 и 66.5%.
Для полной оценки этого результата надо еще прибавить, что латентный период рефлексов при одновременном раздражении угашенного пункта с другими пунктами был значительно короче, чем в норме, а также и двигательная пищевая реакция резче, чем обыкновенно. И то и другое - бесспорные указания на повышение возбудимости, положительное индуцирование остальных пунктов анализатора, кроме специально угашенного. Несколько меньший слюноотделительный эффект от остальных пунктов, очевидно, надо понимать как результат алгебраического суммирования их эффекта с эффектом из угашенного пункта.
Особенный интерес для нас в этих опытах представляет еще следующее. Угашение до нуля на определенном месте кожи производилось повторением неподкрепляемого условного раздражения через 2 минуты. В новой вариации опытов по получении первого нуля процедура угашения продолжалась, и к третьему нулевому раздражению присоединялось раздражение других пунктов кожи. Теперь эффект рефлекса от соединенного раздражения был большей частью совершенно равен нормальному эффекту изолированного положительного раздражения. Значит, углубление торможения, усиление тормозного процесса поведо к более значительной положительной индукции. Рядом с этим в тех же опытах обращает на себя внимание и следующее сопоставление фактов. У этой собаки, как мы видели, к 2 минутам после угашения до нуля иррадиированное торможение на кожном анализаторе еще держится на уровне не ниже 50% 0/ полного торможения, между тем как при совместном раздражении угашаемого пункта в третий раз до нуля (через 2 минуты после предшествующего угашения) и остальных пунктов не видно ни малейшего знака торможения. Следовательно, оно во время повторного тормозного раздражения опять сконцентрировалось в пункте этого раздражения.
В той же работе (Подкопаева) те же результаты были получены на другой собаке, у которой были, между прочим, выработаны положительные пищевые условные рефлексы на определенный тон и удары метронома (120 ударов в минуту) и тормозной (дифференцировка на 60 ударов метронома в минуту). На «фоне» дифференцировочного торможения от метронома тон из 9 проб только один раз дал мало заторможенный эффект, в 4 случаях - нормальную величину его и еще в 4 - величину, даже значительно превосходившую норму.
Для следующих опытов, относящихся к той же теме, были употреблены две собаки. У одной (опыты В. П. Головиной) среди многих пищевых условных рефлексов были положительные: на метроном, определенный тон, свисток, лампочку, вспыхивающую в определенном месте, и механическое раздражение кожи, и три тормозных дифференцировки: на удары метронома другой частоты, на другое место кожи и на другое место вспыхивающей лампочки. Когда все эти рефлексы были хорошо выработаны, применялось то или другое положительное раздражение совместно с каким-нибудь из указанных тормозных раздражений. Все эти пробы дали совершенно однообразный результат. Положительные раздражители в этих комбинациях всегда имели или нормальный для них по величине эффект, или несколько увеличенный, и более часто с укороченным латентным периодом.
У другой собаки (опыты Павловой) также среди многих условных рефлексов были положительные: на свисток, звук бульканья и механическое раздражение кожи, и тормозной дифференцировка на удары метронома. В опытах над этим животным каждое из указанных положительных раздражений по нескольку раз применялось с одним и тем же тормозным раздражением. Здесь результат получился несколько иной. Хотя латентный период также большей частью был укорочен, но величина эффекта комбинированного раздражения почти постоянно была меньше, и часто значительно, сравнительно с величиной эффекта одиночного положительного раздражения. Значит, у этой собаки концентрирование нервных процессов было менее совершенным.
Нужно прибавить, что эти две последние собаки явно принадлежали к двум разным типам нервной системы. В то время как у первой дифференцировки были вообще очень прочны и не страдали от комбинированных раздражений, у второй дифференцировка часто сама по себе была неполной и всегда еще более нарушалась после комбинации положительными раздражениями.
Опыты последнего ряда ясно показывают, с одной стороны, как положительная индукция, возникающая под влиянием тормозного раздражения определеннкта, ограничивает распространение тормозного процесса, с другой - свидетельствуют, что взаимодействие между раздражительным и тормозным процессами определяется то фазой установки новых отношений в коре (опыт Подкопаева), то типом нервной системы разных животных (опыты Головиной и Павловой).
Лекция тринадцатая. Кора как мозаика: a) примеры мозаичностии коры и наиболее простые способы ее происхождения; 6) изменяемость физиологической роли отдельных пунктов; в) стойкость некоторых пунктов. Кора как объединенная сложная динамическая система
Мм. гг.! Как очевидно из всего сообщенного доселе, бесконечная масса явлений природы постоянно обусловливает посредством аппарата больших полушарий образование то положительных, то отрицательных условных рефлексов и тем подробно определяет всю деятельность животного, его ежедневное поведение. Для каждого из этих рефлексов коре полушарий должна иметься своя точка приложения, т. e. своя клетка или группа клеток. Одна такая единица коры связана с одной деятельностью организма, другая - с другой, одна вызывает деятельность, другая ее не допускает, подавляет. Таким образом кора полушарий должна представлять собой грандиозную мозаику, грандиозную сигнализационную доску. При этом наой доске постоянно остаются огромные запасы пунктов для образования новых сигнальных условных раздражителей, а, кроме того, занятые ранее пункты более или менее часто подвергаются изменениям в их связях с разными деятельностями организма, в их физиологическом значении. Механизм такой функциональной мозаичности коры полушарий как с анатомической стороны, так и с физиологической отчасти намечается и сейчас. Как состав рецепторов из множества элементов, так и обилие клеток коры с их огромным разнообразием в величине, форме и расположении в значительной степени отвечают требованию этой мозаичности. С этим достаточно согласуется, хотя и более грубая, локализация, устанавливаемая современной физиологией коры. Но в целом вопрос о функциональных разграничениях коры, конечно, вопрос чрезвычайной, исключительной сложности. В настоящее время могут быть делаемы только самые первые шаги экспериментального подхода к изучению его наипростейших сторон. Однако и тут предмет все еще сложен и представляет большие трудности для исследования. Пусть функциональное размежевание коры, по крайней мере в некоторых случаях, допускалось бы на чисто анатомических основаниях, т. е. благодаря существованию отдельных клеток, отвечающих отдельным рецепторным элементам. Но мы знаем из факта первоначальной генерализации условиях раздражителей, что раздражение сперва непременно иррадиирует из исходного пункта, захватывает клетки и других рецепторных элементов. Каким же образом оно опять сосредоточивается в исходном пункте? Мы должны различать тут два случая: как размежевываются друг от друга два положительных пункта, когда они связываются с двумя разными безусловными рефлексами, и как разграничиваются положительный и отрицательный (т. е. торможно действующий) пункты, когда они относятся к одному и тому же безусловному рефлексу. Наш наличный материал почти исключительно до последнего времени имел дело только со вторым случаем как с более простым. За решение первого вопроса мы взялись только теперь, и потому мое дальнейшее изложение будет касаться только второго вопроса. Здесь я соединю наши данные, относящиеся к теме о мозаике и не вошедшие в предшествующие лекции. Начнем с наиболее простого и наиболее очевидного случая. Вот собака, у которой, кроме друсловных пищевых рефлексов, имеются кожно-механические: положительный на правом плече и отрицательный на правом бедре. Когда эти рефлексы были хорошо выработаны, впервые испытывались на их эффект новые места: на передней лапе на 17 см вниз от места на плече, на боку на 12 см кзади от места на плече, то же на боку на 15 см кпереди от места на бедре и, наконец, на задней лапе на 18 см вниз от места на бедре. Слюноотделение отмечалось за 30 секунд изолированного действия условного раздражения. То же получилось и еще на двух собаках, служивших для этих опытов (опыты Фурсикова).
Мы видим, что под влиянием внешних раздражений, действовавших при разных условиях (одно сопровождалось безусловным рефлексом, другое - нет), в каждом анализаторе возникли два разных пространственно разделенных пункта: пункт раздражения и пункт торможения, причем около каждого из них образовался соответственный район. Эти районы, распространяясь навстречу друг другу, пространственно разграничивались. Резкое уменьшение положительного эффекта на боку ближе к бедру и нуль эффекта на задней ноге надо рассматривать как преобладание тормозного процесса, потому что генерализация условного положительного кожно-механического раздражения, как это мы видели в десятой лекции, обыкновенно захватывает весь анализатор и уменьшение эффекта по мере удаления от места выработанного рефлекса происходит очень постепенно, а не обрывисто.
Таким образом первое основание для функциональной мозаики коры дают внешние раздражения, производящие противоположные процессы, как это мы уже видели в лекции об анализаторной деятельности коры.
Затем на следующих трех собаках мы поставили более сложные опыты. У двух собак на разных тонах (опыты В. В. Сирятского), а у одной на разных местах кожи (опыта Купалова) мы правильно чередовали несколько положительных и отрицательных пунктов. Это предпринималос несколькими целями, чтобы видеть: как вырабатывается мозаика, как далеко она может простираться, какой достигает точности, как взаимно действуют ее разные пункты, как относятся промежуточные пункты и как эта мозаика отзывается на общем состоянии животного.
Некоторые из результатов этих опытов веду сейчас, другие будут использованы в других отделах моего изложения. Вообще же исследование этих животных еще не закончено.
У одной собаки из пяти смежных октав фисгармонии от большой октавы до третьей включительнот 64 колебаний в секунду до 1440) все тоны до были сделаны положительными условными возбудителями пищевой реакции, все тоны фа отрицательными. В протоколах тоны до и фа отдельно обозначены номерами по порядку, начиная с низких.
Вот окончательное положение дела у этой собаки.
То же получилось и еще на двух собаках, служивших для этих опытов (опыты Фурсикова).
Мы видим, что под влиянием внешних раздражений, действовавших при разных условиях (одно сопровождалось безусловным рефлексом, другое - нет), в каждом анализаторе возникли два разных пространственно разделенных пункта: пункт раздражения и пункт торможения, причем около каждого из них образовался соответственный район. Эти районы, распространяясь навстречу друг другу, пространственно разграничивались. Резкое уменьшение положительного эффекта на боку ближе к бедру и нуль эффекта на задней ноге надо рассматривать как преобладание тормозного процесса, потому что генерализация условного положительного кожно-механического раздражения, как это мы видели в десятой лекции, обыкновенно захватывает весь анализатор и уменьшение эффекта по мере удаления от места выработанного рефлекса происходит очень постепенно, а не обрывисто.
Таким образом первое основание для функциональной мозаики коры дают внешние раздражения, производящие противоположные процессы, как это мы уже видели в лекции об анализаторной деятельности коры.
Затем на следующих трех собаках мы поставили более сложные опыты. У двух собак на разных тонах (опыты В. В. Сирятского), а у одной на разных местах кожи (опыта Купалова) мы правильно чередовали несколько положительных и отрицательных пунктов. Это предпринималос несколькими целями, чтобы видеть: как вырабатывается мозаика, как далеко она может простираться, какой достигает точности, как взаимно действуют ее разные пункты, как относятся промежуточные пункты и как эта мозаика отзывается на общем состоянии животного.
Некоторые из результатов этих опытов веду сейчас, другие будут использованы в других отделах моего изложения. Вообще же исследование этих животных еще не закончено.
У одной собаки из пяти смежных октав фисгармонии от большой октавы до третьей включительнот 64 колебаний в секунду до 1440) все тоны до были сделаны положительными условными возбудителями пищевой реакции, все тоны фа отрицательными. В протоколах тоны до и фа отдельно обозначены номерами по порядку, начиная с низких.
Вот окончательное положение дела у этой собаки.
 У другой собаки в пределах одной октавы от фа малой октавы до фа диез первой октавы тоны тоже сделаны чередующимися положительными и отрицательными условными раздражителями, но без правильного ритма, что касается промежутков между действующими тонами и на более коротких расстояниях между ними, именно: фа диез - положительный, ми - отрицательный, до - положительный, ля отрицательный, соль положительный и фа - отрицательный.
Вот окончательное положение дела у этой собаки.
У другой собаки в пределах одной октавы от фа малой октавы до фа диез первой октавы тоны тоже сделаны чередующимися положительными и отрицательными условными раздражителями, но без правильного ритма, что касается промежутков между действующими тонами и на более коротких расстояниях между ними, именно: фа диез - положительный, ми - отрицательный, до - положительный, ля отрицательный, соль положительный и фа - отрицательный.
Вот окончательное положение дела у этой собаки.
 У третьей собаки были выработаны условные кожно-механические пищевые рефлексы. Приборы для раздражения были расположены на строго определеннных местах по линии от левой голени через все туловище до левого предплечья, на одинаковых расстояниях один от другого, именно на расстоянии 12 см, считая от центра приборов. Счет приборов велся от голени к предплечью. Всех приборов было 9. Раздражения нечетными приборами были сделаны положительными, раздражения четными отрицательными.
Вот один из более резких опытов.
У третьей собаки были выработаны условные кожно-механические пищевые рефлексы. Приборы для раздражения были расположены на строго определеннных местах по линии от левой голени через все туловище до левого предплечья, на одинаковых расстояниях один от другого, именно на расстоянии 12 см, считая от центра приборов. Счет приборов велся от голени к предплечью. Всех приборов было 9. Раздражения нечетными приборами были сделаны положительными, раздражения четными отрицательными.
Вот один из более резких опытов.
 Представленная функциональная мозаичность коры давалась ей трудно, но только на первых порах выработки. Но чем дело шло дальше, тем выработка становилась все более легкой. Ocoбенный интерес заключается в том, что в конце выработки у первой и третьей собак некоторые новые рефлексы были готовы сами собой сразу. Нужно иметь в виду, что новые условные раздражения в этих случаях возникали в районах противоположного нервного процесса.
У первой собаки был, и уже давно, выработан положительный условный рефлекс на до4. В данном опыте это до4 имело эффект в 8 капель за 30 секунд. Когда впервые было применено раздражение фа4, то оно осталось без положительного эффекта, т. е., надо было думать, было прямо тормозным. Это доказал опыт следующего дня. Этот день был начат с пробы фа4, который и теперь оказался без положительного эффекта, а последовавшее за ним положительное раздражение до4 являлось в значительной степени заторможенным. Следовательно, фа4 действительно сделалось тормозным само собой без выработки, находясь в районе положительного до4. То же самое повторилось с до5, которое, испробованное в первый раз, дало положительный эффект, находясь в районе тормозного фа4.
У третьей собаки с кожно-механическими условными раздражениями оба крайние положительные раздражения № 1 и № 9 также были налицо прямо без выработки, притом с полным эффектом, находясь оба в районах тормозных пунктов № 2 и № 8.
Естественное объяснение последних фактов надо искать в явлениях взаимной индукции, исходящей из пунктов фиксированного раздражения и торможения, пунктов, расположенных притом ранее в правильном чередовании. Отсюда становится также понятным, почему ритмическая деятельность вообще особенно легка и выгодна. На всех только что описанных собаках постоянно повторялся факт, что при правильном чередовании положительных и отрицательных раздражении рефлексы отличались особенной точностью. Вот пример, как это чередование постепенно даже в течение одного опыта все резче отграничивает положительные и отрицательные пункты, если их эффекты вначале в значительной степени сливались.
Третья собака с кожно-механическими раздражениями.
Представленная функциональная мозаичность коры давалась ей трудно, но только на первых порах выработки. Но чем дело шло дальше, тем выработка становилась все более легкой. Ocoбенный интерес заключается в том, что в конце выработки у первой и третьей собак некоторые новые рефлексы были готовы сами собой сразу. Нужно иметь в виду, что новые условные раздражения в этих случаях возникали в районах противоположного нервного процесса.
У первой собаки был, и уже давно, выработан положительный условный рефлекс на до4. В данном опыте это до4 имело эффект в 8 капель за 30 секунд. Когда впервые было применено раздражение фа4, то оно осталось без положительного эффекта, т. е., надо было думать, было прямо тормозным. Это доказал опыт следующего дня. Этот день был начат с пробы фа4, который и теперь оказался без положительного эффекта, а последовавшее за ним положительное раздражение до4 являлось в значительной степени заторможенным. Следовательно, фа4 действительно сделалось тормозным само собой без выработки, находясь в районе положительного до4. То же самое повторилось с до5, которое, испробованное в первый раз, дало положительный эффект, находясь в районе тормозного фа4.
У третьей собаки с кожно-механическими условными раздражениями оба крайние положительные раздражения № 1 и № 9 также были налицо прямо без выработки, притом с полным эффектом, находясь оба в районах тормозных пунктов № 2 и № 8.
Естественное объяснение последних фактов надо искать в явлениях взаимной индукции, исходящей из пунктов фиксированного раздражения и торможения, пунктов, расположенных притом ранее в правильном чередовании. Отсюда становится также понятным, почему ритмическая деятельность вообще особенно легка и выгодна. На всех только что описанных собаках постоянно повторялся факт, что при правильном чередовании положительных и отрицательных раздражении рефлексы отличались особенной точностью. Вот пример, как это чередование постепенно даже в течение одного опыта все резче отграничивает положительные и отрицательные пункты, если их эффекты вначале в значительной степени сливались.
Третья собака с кожно-механическими раздражениями.
 Вот еще примеры, взятые из работы над первой собакой, согда рефлексы вообще еще не были регулярны.
Вот еще примеры, взятые из работы над первой собакой, согда рефлексы вообще еще не были регулярны.
 Та же собака спустя 3 дня.
Та же собака спустя 3 дня.
 Таким образом дальнейшее основание для функциональной мозаичности коры надо видеть во взаимной индукции нервных процессов.
Затем в этих же опытах мы занимались определением районов (полей) как раздражительного, так и тормозного процессов, а также возможных индифферентомежуточных между деятельными районами.
На собаках с тоновой мозаикой мы пробовали другие тоны, находящиеся между нашими положительными и отрицательными раздражителями, и наблюдали их эффекты. Положительные эффекты в виде наступающего слюноотделения, конечно, были очевидны прямо. Что касается до нулевых, то надо было разобраться особыми вариациями опыта в их индифферентном или тормозном значении. Мы испыталих ради этого или на их последовательное торможение, или на их положительную индукцию.
Таким образом дальнейшее основание для функциональной мозаичности коры надо видеть во взаимной индукции нервных процессов.
Затем в этих же опытах мы занимались определением районов (полей) как раздражительного, так и тормозного процессов, а также возможных индифферентомежуточных между деятельными районами.
На собаках с тоновой мозаикой мы пробовали другие тоны, находящиеся между нашими положительными и отрицательными раздражителями, и наблюдали их эффекты. Положительные эффекты в виде наступающего слюноотделения, конечно, были очевидны прямо. Что касается до нулевых, то надо было разобраться особыми вариациями опыта в их индифферентном или тормозном значении. Мы испыталих ради этого или на их последовательное торможение, или на их положительную индукцию.
 Мы видим, что из расположенных между положительным до и отрицательным ми промежуточные тон ре и полутоны до диез и ми бемоль оказались разного значения. Полутон до до диез имел хотя и уменьшенный, но положительный эффект, т. е. принадлежал к району положительного до. Тон ре и полутон ми бемоль, повидимому, были одинаковы по действию, давши нуль. Но испытание их на последовательное торможение обнаружило их разницу. 2 минуты спустя после применения полутона ми бемоль условное механическое раздражение потеряло в своем обыкновенном эффекте 50%, т. е. было заторможена, после же тона ре в тех же условиях времени осталось без изменения. Следовательно, полутон ми безоль бесспорно находился в районе отрицательного ми, тон же ре или был совершенно индифферентным, или заторможен в гораздо меньшей степени. Последнее надо было решить более тонкими опытами, и потому вопрос об истинно индифферентных пунктах подвергается дальнейшему исследованию.
В этих же опытах исследовалось также взаимодействие пунктов при непосредственном переходе c раздражением от одних положительных пунктов к другим положительным, от одних отрицательных к другим отрицательным, от положительных к отрицательным, и наоборот. Но результаты большей частью получались такой сложности, что мы были вынуждены решать ту же задачу в более упрощстановке.
Как указано было еще в первой лекции и упомянуто опять в начале этой, функциональная мозаика коры не только постоянно может пополняться, но и подлежит частой переделке, т. е. одни и те же пункты коры связываются то с одной, то с другой физиологической деятельностью организма. К последнему пункту относится у нас только одно исследование (опыты С. С. Фридемана).
Одни и те же агенты делались сначала условныйи пищевыми раздражителями, а затем переделывались кислотные, и наоборот - прежние кислотные раздражители превращались в пищевые. У двух собак первоначальный условный пищевой раздражитель был превращен в условный кислотный, а у третьей кислотный в пищевой. Производилось это, ечно, так, что условный раздражитель сопровождался теперь другим безусловным раздражителем. Общий ход дела представляется в следующем виде. Условный раздражитель быстро, даже в течение одного опыта, теряет свой прежний слюноотделительный эффект и становится нулевым на более или менее значительный срок и затем снова приобретает слюногонная действие - и это уже был другой рефлекс, как показывало резкое изменение в составе слюны подчелюстной железы. Вся эта процедура требует около 30 сочетаний условного раздражителя с новым безусловным раздражителем. Когда затем у первых двух собак после довольно длинного периода с условными кислотными рефлексами условные раздражители еще раз подвергались превращению опять в пищевые, это превращение произошло очень быстро, через небольшое число раз. Значит, первая связь до известной степени уцелела, несмотря на существование второй связи.
В настоящем исследовании главный интерес для нас привязывался, однако, к другому пункту. Мы желали знать, сохранится ли дифференцировка, выработанная для условного раздражителя на одном рефлексе, при превращении его в условный раздражитель другого рефлекса. У всех трех собак результат получился тождественный. Я опишу его более подробно для одной из собак. Условным пищевым раздражителем был тон ми (2600 колебаний в секунду). От него была выработана и хорошо концентрирована дифференцировка на соседний низкий тон (2324 колебания в секунду). Затем условный пищевой раздражитель был превращен в условный кислотный. Та же дифференцировка, испробованная в первый раз на кислотном рефлексе, оказалась полной и в такой же степени концентрированной. Теперь была выработана новая более тонкая дифференцировка на полутон выше (2760 колебаний в секунду). Когда тон ми снова был сделан пищевым условным раздражителем, то и эта новая дифференцировка также вполне сохранилась.
На протяжении предыдущих лекций мы достаточно познакомились с фактом, что одни и те же пункты коры, смотря по условиям, являются местом то раздражительного процесса, то тормозного, вызывают то одну работу организма, то другую. И это изменение их физиологической роли происходит довольно легко, т. e. более или менее скоро они меняют свое состояние (прошу припомнить опыты с отрицательной индукцией) и свои связи (как показывают только что сообщенные опыты).
Но мы имеем также и факты, когда определенная роль известных пунктов и отделов коры или даяе временные отношения процессов в них делаются чрезвычайно упорными, трудно или совсем не поддающимися переменам. С этим мы встретились в работе о следовых положительных и отрицательных условных рефлексах (опыты Фролова). Тут некоторые явления носили такой исключительный характер, что считаю необходимым описать их подробно.
У одной собаки был выработан условный следовой кислотный рефлекс на тон органной трубы в 1740 колебаний в секунду, причем тон продолжался 15 секунд и по прекращении его, спустя 30 секунд, производилось вливание собаке в рот кислоты. Этот следовой рефлекс повторялся 994 раза в течение 1 года и 9 месяцев. Латентный период, считая от начала тона, составлял около 25 секунд, или, считая иначе, 10 секунд от начала следов тона. Когда одни опыты с этим рефлексом были покончены, было решено тот же тон переделать для других опытов из следового условного раздражителя в наличный условный, т. е. получать от него эффект не по прекращении, а еще во время его действия.
Для этого тон на пятнадцатой секунде его продолжения сопровождался вливанием кислоты. После -вадцатикратного повторения такой процедуры не только не появилось условного рефлекса в течение 15 секунд, но после вливания кислоты слюноотделение начиналось лишь спустя 10 секунд, т. е. спустя те же 25 секунд от начала тона. Примененные вместо тона другие звуковые раздражители ничего не изменили в положении дела. Когда же тон и эти раздражители изолированно без вливания кислоты применялись 45 секунд, то спустя около 25 секунд от начала их слюноотделение начиналось и быстро достигало значительной величины (10 капель за остальные 20 секунд). После этого вливание кислоты начали производить спустя только 2 секунды от начала звучания тона. И, однако, после очень большого количества (многих десятков раз) таких сочетаний слюноотделение как во время вливания кислоты, так и при одном звучании тона без кислоты начиналось только около двадцатойдвадцать пятой секунды от начала тона.
Вот последний относящийся сюда опыт.
Мы видим, что из расположенных между положительным до и отрицательным ми промежуточные тон ре и полутоны до диез и ми бемоль оказались разного значения. Полутон до до диез имел хотя и уменьшенный, но положительный эффект, т. е. принадлежал к району положительного до. Тон ре и полутон ми бемоль, повидимому, были одинаковы по действию, давши нуль. Но испытание их на последовательное торможение обнаружило их разницу. 2 минуты спустя после применения полутона ми бемоль условное механическое раздражение потеряло в своем обыкновенном эффекте 50%, т. е. было заторможена, после же тона ре в тех же условиях времени осталось без изменения. Следовательно, полутон ми безоль бесспорно находился в районе отрицательного ми, тон же ре или был совершенно индифферентным, или заторможен в гораздо меньшей степени. Последнее надо было решить более тонкими опытами, и потому вопрос об истинно индифферентных пунктах подвергается дальнейшему исследованию.
В этих же опытах исследовалось также взаимодействие пунктов при непосредственном переходе c раздражением от одних положительных пунктов к другим положительным, от одних отрицательных к другим отрицательным, от положительных к отрицательным, и наоборот. Но результаты большей частью получались такой сложности, что мы были вынуждены решать ту же задачу в более упрощстановке.
Как указано было еще в первой лекции и упомянуто опять в начале этой, функциональная мозаика коры не только постоянно может пополняться, но и подлежит частой переделке, т. е. одни и те же пункты коры связываются то с одной, то с другой физиологической деятельностью организма. К последнему пункту относится у нас только одно исследование (опыты С. С. Фридемана).
Одни и те же агенты делались сначала условныйи пищевыми раздражителями, а затем переделывались кислотные, и наоборот - прежние кислотные раздражители превращались в пищевые. У двух собак первоначальный условный пищевой раздражитель был превращен в условный кислотный, а у третьей кислотный в пищевой. Производилось это, ечно, так, что условный раздражитель сопровождался теперь другим безусловным раздражителем. Общий ход дела представляется в следующем виде. Условный раздражитель быстро, даже в течение одного опыта, теряет свой прежний слюноотделительный эффект и становится нулевым на более или менее значительный срок и затем снова приобретает слюногонная действие - и это уже был другой рефлекс, как показывало резкое изменение в составе слюны подчелюстной железы. Вся эта процедура требует около 30 сочетаний условного раздражителя с новым безусловным раздражителем. Когда затем у первых двух собак после довольно длинного периода с условными кислотными рефлексами условные раздражители еще раз подвергались превращению опять в пищевые, это превращение произошло очень быстро, через небольшое число раз. Значит, первая связь до известной степени уцелела, несмотря на существование второй связи.
В настоящем исследовании главный интерес для нас привязывался, однако, к другому пункту. Мы желали знать, сохранится ли дифференцировка, выработанная для условного раздражителя на одном рефлексе, при превращении его в условный раздражитель другого рефлекса. У всех трех собак результат получился тождественный. Я опишу его более подробно для одной из собак. Условным пищевым раздражителем был тон ми (2600 колебаний в секунду). От него была выработана и хорошо концентрирована дифференцировка на соседний низкий тон (2324 колебания в секунду). Затем условный пищевой раздражитель был превращен в условный кислотный. Та же дифференцировка, испробованная в первый раз на кислотном рефлексе, оказалась полной и в такой же степени концентрированной. Теперь была выработана новая более тонкая дифференцировка на полутон выше (2760 колебаний в секунду). Когда тон ми снова был сделан пищевым условным раздражителем, то и эта новая дифференцировка также вполне сохранилась.
На протяжении предыдущих лекций мы достаточно познакомились с фактом, что одни и те же пункты коры, смотря по условиям, являются местом то раздражительного процесса, то тормозного, вызывают то одну работу организма, то другую. И это изменение их физиологической роли происходит довольно легко, т. e. более или менее скоро они меняют свое состояние (прошу припомнить опыты с отрицательной индукцией) и свои связи (как показывают только что сообщенные опыты).
Но мы имеем также и факты, когда определенная роль известных пунктов и отделов коры или даяе временные отношения процессов в них делаются чрезвычайно упорными, трудно или совсем не поддающимися переменам. С этим мы встретились в работе о следовых положительных и отрицательных условных рефлексах (опыты Фролова). Тут некоторые явления носили такой исключительный характер, что считаю необходимым описать их подробно.
У одной собаки был выработан условный следовой кислотный рефлекс на тон органной трубы в 1740 колебаний в секунду, причем тон продолжался 15 секунд и по прекращении его, спустя 30 секунд, производилось вливание собаке в рот кислоты. Этот следовой рефлекс повторялся 994 раза в течение 1 года и 9 месяцев. Латентный период, считая от начала тона, составлял около 25 секунд, или, считая иначе, 10 секунд от начала следов тона. Когда одни опыты с этим рефлексом были покончены, было решено тот же тон переделать для других опытов из следового условного раздражителя в наличный условный, т. е. получать от него эффект не по прекращении, а еще во время его действия.
Для этого тон на пятнадцатой секунде его продолжения сопровождался вливанием кислоты. После -вадцатикратного повторения такой процедуры не только не появилось условного рефлекса в течение 15 секунд, но после вливания кислоты слюноотделение начиналось лишь спустя 10 секунд, т. е. спустя те же 25 секунд от начала тона. Примененные вместо тона другие звуковые раздражители ничего не изменили в положении дела. Когда же тон и эти раздражители изолированно без вливания кислоты применялись 45 секунд, то спустя около 25 секунд от начала их слюноотделение начиналось и быстро достигало значительной величины (10 капель за остальные 20 секунд). После этого вливание кислоты начали производить спустя только 2 секунды от начала звучания тона. И, однако, после очень большого количества (многих десятков раз) таких сочетаний слюноотделение как во время вливания кислоты, так и при одном звучании тона без кислоты начиналось только около двадцатойдвадцать пятой секунды от начала тона.
Вот последний относящийся сюда опыт.
 В опыте условное раздражение всякий раз подкреплялось кислотой.
Наконец на тон fis тонвариатора был выработан пищевой рефлекс, но и на нем латентный период упорно держался на 24 28 секундах.
И лишь когда условным кислотным раздражением было взято кожно-механическое раздраодкрепляемое через 2 секунды от его начала, на двадцать четвертой пробе получилось условное слюноотделение за 30 секунд в 12 капель с латентным периодом в 2 секунды.
То же упорство выработанного тормозного процесса наблюдалось и у другой собаки. У нее из следов тона органной трубы в 1740 колебаний в секунду был сделан условный тормоз к ударам метронома (104 удара в минуту) как положительному условному раздражителю. Даже следы тона спустя 60 секунд по прекращении его были действительным условным тормозом. Какое торможение, конечно, должно было вырабатываться очень постепенно, начиная с более ранних следов, и очень медленно. A вся выработка и работа с таким условным торможением заняла более двух лет. Нужно еще прибавить, что к этому условному тормозу были присоединены и его дифференцировки. Следовательно, торможение было, так сказать, очень напряженное. Затем собака должна была служить для другой темы, ради которой было приступлено к выработке положительного условного раздражителя из звука телесронной пластинки, приводимой в колебание переменным током частоты, почти совпадающей с числом колебаний тона. Положительный условный рефлекс необычно долго не проявлялся. Тогда новый агент был применен перед ударами метронома - и тогда обнаружилось его резкое условное тормозящее действие. Имея пример стойкости торможения на первой собаке, здесь мы дальнейшую процедуру варьировали таким образом, что стали сопровождать безусловным раздражителем тормозную комбинацию звука телефонной пластинки с ударами метронома, т. е. действовали точно обратно тому, как ранее поступали при образовании тормозной комбинации из следов тона с ударами метронома. И, действительно, теперь довольно скоро тормозная комбинация стала положительным раздражителем, а еще несколько времени спустя и звук мембраны также получил положительное условное действие. Однако сам тон органной трубы не сделался от этого положительным раздражителем, и его пришлось лишать тормозного действия особо и так же, как было поступлено с звуком телефонной пластинки. Но на этот раз цель была достигнута не так скоро.
Описанные случаи стойкости тормозного процесса должны считаться чрезвычайными. Вероятно, помимо напряженности процесса здесь существовали и еще особые основания для нее в условиях данных опытов. Может быть, объяснению их послужат и наши опыты, относящиеся к гипнотизму и сну, о которых речь впереди.
Рядом с этими случаями стойкости тормозного процесса в отдельных пунктах коры можно поставить случай стойкости в отдельных пунктах и процесса раздражения. У собаки (опыты Б. Н. Бирмана) был выработан условный пищевой рефлекс на тон фисгармонии в 256 колебаний в секунду. От него были отдифференцированы 22 тона вверх до тона 768 колебаний в секунду и вниз до тона 85 колебаний в секунду, т. е., как обыкновенно, тон 256 колебаний в секунду сопровождался едой, а все остальные нет. Положительный тон поименялся в каждом опыте по нескольку раз, а из тормозных то те, то другие. Таким образом положительный пункт был как бы окружен тормозным, но это повело не к уменьшению его эффекта: а, наоборот, к резкому увеличению. Затем у собаки был произведен глубокий физиологический сон. Среди этого сна, когда резкий свисток или сильный стук кулаком в дверь экспериментальной комнаты, где стояла собака, не будили собаку, положительный тон сейчас же делал собаку бодрой, причем получался полный условный слюноотделительный эффект. Так как в последующих лекциях будет доказано нашими опытами, что сон есть торможение, то в описанном опыте мы имеем пример стойкости в отдельных пунктах коры процесса раздражения при сильном натиске торможения.
Если, с одной точки зрения, кору больших полушарий можно рассматривать как мозаику, состоящую из бесчисленной массы отдельных пунктов с определенной физиологической ролью в данный момент, то с другой - мы имеем в ней сложнейшую динамическую систему, постоянно стремящуюся к объединению (интеграции) и к стереотипности объединенной деятельности. Всякое новое местное воздействие на эту систему дает себя знать более или менее во всей системе. Мы выработали известное количество условных рефлексов. Прибавление новых положительных и особенно тормозных большей частью сейчас же отзывается на состоянии прежних (опыты Анрепа и других). Мало этого. Если рефлексы остаются те же и мы меняем только раз установленный и долго поддерживаемый порядок их, размер эффекта условных рефлексов резко меняется в сторону уменьшения его, т. e. в сторону преобладания тормозного процесса.
Вот пример этого, взятый из работы Д. И. Соловейчика. У собаки через каждые 10 минут применяются следующие условные пищевые раздражители: удары метронома, вспыхивание электрической лампочки, свисток и механическое раздражение кожи, в приведенном порядке. В каждом опыте этот ряд повторяется.
В опыте условное раздражение всякий раз подкреплялось кислотой.
Наконец на тон fis тонвариатора был выработан пищевой рефлекс, но и на нем латентный период упорно держался на 24 28 секундах.
И лишь когда условным кислотным раздражением было взято кожно-механическое раздраодкрепляемое через 2 секунды от его начала, на двадцать четвертой пробе получилось условное слюноотделение за 30 секунд в 12 капель с латентным периодом в 2 секунды.
То же упорство выработанного тормозного процесса наблюдалось и у другой собаки. У нее из следов тона органной трубы в 1740 колебаний в секунду был сделан условный тормоз к ударам метронома (104 удара в минуту) как положительному условному раздражителю. Даже следы тона спустя 60 секунд по прекращении его были действительным условным тормозом. Какое торможение, конечно, должно было вырабатываться очень постепенно, начиная с более ранних следов, и очень медленно. A вся выработка и работа с таким условным торможением заняла более двух лет. Нужно еще прибавить, что к этому условному тормозу были присоединены и его дифференцировки. Следовательно, торможение было, так сказать, очень напряженное. Затем собака должна была служить для другой темы, ради которой было приступлено к выработке положительного условного раздражителя из звука телесронной пластинки, приводимой в колебание переменным током частоты, почти совпадающей с числом колебаний тона. Положительный условный рефлекс необычно долго не проявлялся. Тогда новый агент был применен перед ударами метронома - и тогда обнаружилось его резкое условное тормозящее действие. Имея пример стойкости торможения на первой собаке, здесь мы дальнейшую процедуру варьировали таким образом, что стали сопровождать безусловным раздражителем тормозную комбинацию звука телефонной пластинки с ударами метронома, т. е. действовали точно обратно тому, как ранее поступали при образовании тормозной комбинации из следов тона с ударами метронома. И, действительно, теперь довольно скоро тормозная комбинация стала положительным раздражителем, а еще несколько времени спустя и звук мембраны также получил положительное условное действие. Однако сам тон органной трубы не сделался от этого положительным раздражителем, и его пришлось лишать тормозного действия особо и так же, как было поступлено с звуком телефонной пластинки. Но на этот раз цель была достигнута не так скоро.
Описанные случаи стойкости тормозного процесса должны считаться чрезвычайными. Вероятно, помимо напряженности процесса здесь существовали и еще особые основания для нее в условиях данных опытов. Может быть, объяснению их послужат и наши опыты, относящиеся к гипнотизму и сну, о которых речь впереди.
Рядом с этими случаями стойкости тормозного процесса в отдельных пунктах коры можно поставить случай стойкости в отдельных пунктах и процесса раздражения. У собаки (опыты Б. Н. Бирмана) был выработан условный пищевой рефлекс на тон фисгармонии в 256 колебаний в секунду. От него были отдифференцированы 22 тона вверх до тона 768 колебаний в секунду и вниз до тона 85 колебаний в секунду, т. е., как обыкновенно, тон 256 колебаний в секунду сопровождался едой, а все остальные нет. Положительный тон поименялся в каждом опыте по нескольку раз, а из тормозных то те, то другие. Таким образом положительный пункт был как бы окружен тормозным, но это повело не к уменьшению его эффекта: а, наоборот, к резкому увеличению. Затем у собаки был произведен глубокий физиологический сон. Среди этого сна, когда резкий свисток или сильный стук кулаком в дверь экспериментальной комнаты, где стояла собака, не будили собаку, положительный тон сейчас же делал собаку бодрой, причем получался полный условный слюноотделительный эффект. Так как в последующих лекциях будет доказано нашими опытами, что сон есть торможение, то в описанном опыте мы имеем пример стойкости в отдельных пунктах коры процесса раздражения при сильном натиске торможения.
Если, с одной точки зрения, кору больших полушарий можно рассматривать как мозаику, состоящую из бесчисленной массы отдельных пунктов с определенной физиологической ролью в данный момент, то с другой - мы имеем в ней сложнейшую динамическую систему, постоянно стремящуюся к объединению (интеграции) и к стереотипности объединенной деятельности. Всякое новое местное воздействие на эту систему дает себя знать более или менее во всей системе. Мы выработали известное количество условных рефлексов. Прибавление новых положительных и особенно тормозных большей частью сейчас же отзывается на состоянии прежних (опыты Анрепа и других). Мало этого. Если рефлексы остаются те же и мы меняем только раз установленный и долго поддерживаемый порядок их, размер эффекта условных рефлексов резко меняется в сторону уменьшения его, т. e. в сторону преобладания тормозного процесса.
Вот пример этого, взятый из работы Д. И. Соловейчика. У собаки через каждые 10 минут применяются следующие условные пищевые раздражители: удары метронома, вспыхивание электрической лампочки, свисток и механическое раздражение кожи, в приведенном порядке. В каждом опыте этот ряд повторяется.
 В опыте на другой день порядок тех же раздражителей несколько меняется.
В опыте на другой день порядок тех же раздражителей несколько меняется.
 Это уменьшение величины условных рефлексов при изменении их порядка иногда, как в приведенном опыте, наступает сейчас же, иногда же оно резче обнаруживается на другой день уже при восстановлении старого порядка и продолжается то много дней, то быстро сменяется нормальной величиной.
Уменьшение рефлексов при повторном ряде в первом опыте довольно частое явление, о котором речь будет в следующей дей лекции.
Это уменьшение величины условных рефлексов при изменении их порядка иногда, как в приведенном опыте, наступает сейчас же, иногда же оно резче обнаруживается на другой день уже при восстановлении старого порядка и продолжается то много дней, то быстро сменяется нормальной величиной.
Уменьшение рефлексов при повторном ряде в первом опыте довольно частое явление, о котором речь будет в следующей дей лекции.
Лекция четырнадцатая. Переход корковой клетки под влияниием условного раздражителя в тормозное состояние
Мм. гг.! Условный раздражитель, очевидно адресующийся к корковой клетке, естественно прежде всего должен давать нам сведения об ее свойствах. Характернейшим свойством этой клетки является более или менее быстрый перевод ее под влиянием условного раздражителя в тормозное состояние. В ранних лекциях о внутреннем торможении мы видели, что, как только положительный условный раздражитель остается без подкрепления, без сопровождения безусловным раздражителем, он делается тормозным, т. е. клетка под его действием переходит в тормозное состояние. Как понимать этот факт? Какое его ближайшее основание? Существует ли этот переход только при указанном условии? Какое значение имеет при этом безусловный рефлекс? Ответом на эти вопросы и будет настоящая лекция. Факт перехода корковой клетки в тормозное состояние общее, чем можно было бы предполагать на основании сообщенного доселе о возникновении внутреннего торможения. Клетка переходит в тормозное состояние и при подкреплении. Развитие в ней тормозного состояния без подкрепления есть частный случай более общего отношения. Клетка под влиянием раздражения постоянно, хотя иногда и медленно, стремится к переходу в тормозное состояние. Безусловный раздражитель только задерживает этот переход. Вот самый обычный, постоянно повторяющийся факт. Мы имеем, положим, отставленный на 30 секунд условный рефлекс, т. е. условный раздражитель один, до присоединения безусловного раздражителя, продолжается 30 секунд. Пусть, когда рефлекс вполне выработается, так называемый нами латентный период, т. е. время от начала условного раздражения до начала слюноотделения, составляет 5 секунд. Этот период некоторое время (это время разных собак чрезвычайно разнится) остается почти неизменным. А затем он начинает прогрессивно расти. Наконец дело доходит до того, что за 30 секунд изолированного условного раздражения не наступает ни малейшего слюноотделения. Но стоит экстреннотодвинуть присоединение безусловного раздражителя на 5-10 секунд, и мы видим опять условное слюноотделение. Продолжая опыт попрежнему, т. е. применяя отставленный на 30 секунд рефлекс, мы снова на один раз отодвигаем безусловный раздражитель на те же 5 10 секунд, и теперь уже и за этот срок нет условного слюнаотделения. Надо отодвигать еще дальше. А кончается тем, что мы таким образом уже совсем не можем видеть условного эффекта. Это постепенное исчезание условного слюноотделения при одном и том же отставлении происходит у разных собак в чрезвычайно разные сроки: у одних через недели, даже дни, у других через года. При наших обычных условных раздражителях оно особенно быстро наступает при кожно-температурных (как холодовом, так и тепловом), далее при кожно-механических и большей части зрительных. Всего позже оно дает себя знать при звуковых раздражителях и особенно не сплошных, а более или менее прерывистых. Вот пример разницы между различными агентами как условными раздражителями в отношении скорости перехода корковой клетки под их действием в тормозное состояние. У собаки (опыты Шишло) вырабатывается первый условный рефлекс на механическое раздражение кожи. Еда присоединяется спустя 10 секунд от начала раздражения и только изредка отодвигается на 30 секунд. Рефлекс появился на двадцать седьмом разе. К сто семьдесят девятому разу он (в течение недель) установился на величине 8 капель в минуту. После этого было приступлено к выработке второго рефлекса, также пищевого, на приложение к коже теплоты 45° С. Рефлекс образовался более скоро: на двенадцатом разе при отставлении на 30 секунд получились 4 капли. Несмотря на постоянное подкрепление также через 10 секунд от начала температурного раздражения, он быстро начинает падать и на тридцать третьем разе при отставлении на минуту дает только каплю. Что исчезновение условного рефлекса, несмотря на подкрепление, есть выражение настоящего тормозного состояния клетки под влиянием повторяющегося условного раздражителя, доказывается следующим. Когда мы, благодаря только что указанной разнице в раздражителях, уже не имеем условного слюноотделения при одном из раздражителей, то применение других деятельных раздражителей сейчас же после него сопровождается также их временным ослаблением. Точно так же исключение из ряда раздражителей раздражителя, потерявшего свои положительный эффект, ведет за собой увеличение действия остальных. Значит, этот раздражитель действительно приобрел тормозящее действие. Не следует смешивать описываемое явление тем, что мы в отделе внутреннем торможении назвали запаздыванием. Запаздывание есть тот так называемый нами латентный период, который устанавливается вскоре после выработки каждого условного рефлекса и держится затем без изменения более или менее значительный срок. Для данного же явления характерна именно его неуклонная прогрессивность. Тормозное состояние клетки под действием условного раздражителя наступает тем скорее, чем более отставлен рефлекс, т. е. чем длительнее изолированное условное раздражение. У данной собаки рефлекс, отставленный на 10 секунд, держится долгое время без изменения его величины и может служить для многих опытов; у нее же, отставленный на 30 секунд, быстро делается негодным вследствие превращения в тормозной. У собаки (опыты Петровой) между другими условными пищевыми рефлексами имеется рефлекс на удары метронома. Пока условный раздражитель применялся изолированно, т. е. до присоединения безусловного, 10 секунд, он оставался постоянным. Вот пример. Как только изолированное условное раздражение продолжалось 30 секунд, рефлекс становился не постоянным в величине и в течение опыта при повторении делался нулевым.
Как только изолированное условное раздражение продолжалось 30 секунд, рефлекс становился не постоянным в величине и в течение опыта при повторении делался нулевым.
 Приведенные примеры разделены промежутком в один день.
Ввиду разницы в этом отношении отдельных животных выгодно, даже часто необходимо, пользоваться различно отставленными рефлексами у разных животных. Понятно также, что сильно отставленные рефлексы, запаздывающие, вырабатываются трудно и получались в начале нашей работы далеко не у всех собак.
Тоже обычный факт, что условные раздражители при повторениях в течение отдельного опыта быстро теряют в их положительном эффекте, часто даже только при однократном повторении. Это и выступало в последнем опыте предшествующей лекции. Что в этом случае имеет определяющее значение именно повторение, а не какое-либо другое обстоятельство, например постепенное насыщение животного в течение опыта при пищевых условных рефлексах, доказывается тем, что можно повторять только один из условных раздражителей - и эффект лишь от этого раздражителя резко упадет, тогда как другие рядом с ним могут сохранить свое обычное действие до конца опыта. Пример этого будет приведен позже.
Так как существенный интерес экспериментатора - при многих опытах иметь постоянную величину условного рефлекса, то часто приходится бороться с этим стремлением условных раздражителей уклоняться в сторону торможения. Сначала эмпирически, а потом и сознательно, мы должны были выработать большой ряд мер борьбы с этой помехой в нашей работе. Понятно, что наиболее достигающие цели меры состояли из применения условий, обратных тем, которые вызывалиослабление условных раздражителей .
На первом месте в этом отношении надо поставить резкое укорочение продолжительности изолированного условного раздражения. Если, как наиболее обычнос, рефлекс был отставлен на 30 секунд, мы начинаем подкреплять его через 3--5 секунд после начала действия условного раздражителя. Конечно, на этот период условный раздражитель большей частью делается, так сказать, бесполезным для работы, так как никакого слюноотделения за такой короткий срок тел более не может быть. Но ведь это есть нарочитый период восстановления, можно сказать, лечения условного раздражителя. Зато потом, когда мы возвращаемся к обычному отставлению, мы снова имеем, как в начале работы, большой и постоянный рефлекс. При этом частное правило: после коротко отставленного рефлекса полезно переходить к обычному не сразу, этапами, только постепенно увеличивая отставление. Действительность приема, т. е. как надолго он помогает, зависит как от степени и продолжительности ослабленного состояния условного раздражителя, так и от величины периода с коротким отставлением. Небольшой период восстановляет слабо и очень кратковременно, и наоборот. Точно так же, если ослабление условного раздражителя было слишком значительное и давнее, то даже и обсуждаемый прием, вообще очень действительный, на втот раз отказывает в своей помощи, как увидим сейчас.
Далее отчетливо помогает применение условных раздражителей в течение каждого отдельного опыта без повторений или возможно ограничивая число повторений. Точно так же во многих случаях оказывается полезным простой перерыв в работе с условными рефлексами, даже только на несколько дней.
Затем следует ряд более косвенных мер: введение более сильных агентов в число условных раздражителей, вообще увеличение числа условных раздражителей, влияние положительной индукции и, наконец, усиление безусловных раздражителей. Но в данном случае нас преимущественно интересуют, так сказать, прямые вышеперечисленные меры.
Но есть случаи, когда ни одна из приведенных мер более уже не помогает.
Все положительные условные рефлексы постепенно исчезают у животного. Животное становится вялым на станке во время опыта и даже отказывается от еды, подаваемой ему вслед за условными раздражителями. И это происходит с собаками, которые служили для многих и очень точных опытов в течение годов.
Что это значит? Как быть с такой собакой? В раннее время нашей работы мы бы ее бросили, как уже негодную для нас. Теперь она для нас - вопрос, требующий решения. Решение оказалось очень простым. Стоило бросить, не применять старые условные раздражители, заменить их новыми агентами, и казавшееся неодолимым затруднение исчезло. Новые рефлексы образуются чрезвычайно быстро, что и понятно, так как все посторонние рефлексы, обыкновенно в начале работы с нашими собаками сильно мешающие, давно отпали. Вновь образованные рефлексы скоро достигают максимальной величины и становятся постоянными; короче говоря, собака совершенно возвращается к прежнему, выгодному для нашей работы, состоянию. Ввиду особенно важного значения такого факта, я приведу подробно историю одной из таких собак (опыты Подкопаева).
С этой собакой началась работа с июня 1921 г. У нее постепенно были выработаны разнообразные положительные и отрицательные пищевые условные рефлексы. Кожно-механических рефлексов было несколько (с разных мест кожи), обозначаемых разными номерами. Все рефлексы отличались постоянством величины их эффекта, а также однообразием латентного периода. Собака служила в течение годов для разработки многих тем.
Bo Вот опыт из 1922 года (30 VIII) с положительными рефлексами.
Приведенные примеры разделены промежутком в один день.
Ввиду разницы в этом отношении отдельных животных выгодно, даже часто необходимо, пользоваться различно отставленными рефлексами у разных животных. Понятно также, что сильно отставленные рефлексы, запаздывающие, вырабатываются трудно и получались в начале нашей работы далеко не у всех собак.
Тоже обычный факт, что условные раздражители при повторениях в течение отдельного опыта быстро теряют в их положительном эффекте, часто даже только при однократном повторении. Это и выступало в последнем опыте предшествующей лекции. Что в этом случае имеет определяющее значение именно повторение, а не какое-либо другое обстоятельство, например постепенное насыщение животного в течение опыта при пищевых условных рефлексах, доказывается тем, что можно повторять только один из условных раздражителей - и эффект лишь от этого раздражителя резко упадет, тогда как другие рядом с ним могут сохранить свое обычное действие до конца опыта. Пример этого будет приведен позже.
Так как существенный интерес экспериментатора - при многих опытах иметь постоянную величину условного рефлекса, то часто приходится бороться с этим стремлением условных раздражителей уклоняться в сторону торможения. Сначала эмпирически, а потом и сознательно, мы должны были выработать большой ряд мер борьбы с этой помехой в нашей работе. Понятно, что наиболее достигающие цели меры состояли из применения условий, обратных тем, которые вызывалиослабление условных раздражителей .
На первом месте в этом отношении надо поставить резкое укорочение продолжительности изолированного условного раздражения. Если, как наиболее обычнос, рефлекс был отставлен на 30 секунд, мы начинаем подкреплять его через 3--5 секунд после начала действия условного раздражителя. Конечно, на этот период условный раздражитель большей частью делается, так сказать, бесполезным для работы, так как никакого слюноотделения за такой короткий срок тел более не может быть. Но ведь это есть нарочитый период восстановления, можно сказать, лечения условного раздражителя. Зато потом, когда мы возвращаемся к обычному отставлению, мы снова имеем, как в начале работы, большой и постоянный рефлекс. При этом частное правило: после коротко отставленного рефлекса полезно переходить к обычному не сразу, этапами, только постепенно увеличивая отставление. Действительность приема, т. е. как надолго он помогает, зависит как от степени и продолжительности ослабленного состояния условного раздражителя, так и от величины периода с коротким отставлением. Небольшой период восстановляет слабо и очень кратковременно, и наоборот. Точно так же, если ослабление условного раздражителя было слишком значительное и давнее, то даже и обсуждаемый прием, вообще очень действительный, на втот раз отказывает в своей помощи, как увидим сейчас.
Далее отчетливо помогает применение условных раздражителей в течение каждого отдельного опыта без повторений или возможно ограничивая число повторений. Точно так же во многих случаях оказывается полезным простой перерыв в работе с условными рефлексами, даже только на несколько дней.
Затем следует ряд более косвенных мер: введение более сильных агентов в число условных раздражителей, вообще увеличение числа условных раздражителей, влияние положительной индукции и, наконец, усиление безусловных раздражителей. Но в данном случае нас преимущественно интересуют, так сказать, прямые вышеперечисленные меры.
Но есть случаи, когда ни одна из приведенных мер более уже не помогает.
Все положительные условные рефлексы постепенно исчезают у животного. Животное становится вялым на станке во время опыта и даже отказывается от еды, подаваемой ему вслед за условными раздражителями. И это происходит с собаками, которые служили для многих и очень точных опытов в течение годов.
Что это значит? Как быть с такой собакой? В раннее время нашей работы мы бы ее бросили, как уже негодную для нас. Теперь она для нас - вопрос, требующий решения. Решение оказалось очень простым. Стоило бросить, не применять старые условные раздражители, заменить их новыми агентами, и казавшееся неодолимым затруднение исчезло. Новые рефлексы образуются чрезвычайно быстро, что и понятно, так как все посторонние рефлексы, обыкновенно в начале работы с нашими собаками сильно мешающие, давно отпали. Вновь образованные рефлексы скоро достигают максимальной величины и становятся постоянными; короче говоря, собака совершенно возвращается к прежнему, выгодному для нашей работы, состоянию. Ввиду особенно важного значения такого факта, я приведу подробно историю одной из таких собак (опыты Подкопаева).
С этой собакой началась работа с июня 1921 г. У нее постепенно были выработаны разнообразные положительные и отрицательные пищевые условные рефлексы. Кожно-механических рефлексов было несколько (с разных мест кожи), обозначаемых разными номерами. Все рефлексы отличались постоянством величины их эффекта, а также однообразием латентного периода. Собака служила в течение годов для разработки многих тем.
Bo Вот опыт из 1922 года (30 VIII) с положительными рефлексами.
 О пыт из 1923 года (6 VIII): положительные рефлексы и отрицательный (условное торможение).
О пыт из 1923 года (6 VIII): положительные рефлексы и отрицательный (условное торможение).
 О п ы т из 1924 года (12 VI): положительные рефлексы и отрицательный (условное торможение с другим агентом).
О п ы т из 1924 года (12 VI): положительные рефлексы и отрицательный (условное торможение с другим агентом).
 К концу этого года рефлексы часто становились нулевыми, и собака брала подаваемую ей после условного раздражения еду не сразу. Были пущены в ход разные наши приемы для устранения нарастающего торможения: применение вместо кожномеханических раздражений главным образов звуковых, вместо отставленных рефлексов почти совпадающие, отдыхи от опытов даже на большие сроки (полтора месяца), усиление безусловного раздражителя и ведение опыта вместо станка на полу. Но все эти приемы имели только кратковременное действие. Собака делалась на станке все более и более вялой и часто совершенно отказывалась от еды после условных раздражителей. Так прошел весь 1925 год. В конце этого года были отменены все старые условные раздражители и начата выработка рефлексов на новые агенты. Это быстро и резко изменило положение дела. Собака на станке снова стала бодрой и еду брала сейчас же, как она подавалась после новых условных раздражителей. Появились постоянные и значительные величины для условного раздражения. Латентный период вернулся к его первоначальному размеру.
Вот один из ранних опытов 21 I 1926 этой новой фазы в работе с собакой. Рефлексы отставлены пока на 15 секунд, и сделана попытка вставить среди новых раздражителей один из старых - удары метронома.
К концу этого года рефлексы часто становились нулевыми, и собака брала подаваемую ей после условного раздражения еду не сразу. Были пущены в ход разные наши приемы для устранения нарастающего торможения: применение вместо кожномеханических раздражений главным образов звуковых, вместо отставленных рефлексов почти совпадающие, отдыхи от опытов даже на большие сроки (полтора месяца), усиление безусловного раздражителя и ведение опыта вместо станка на полу. Но все эти приемы имели только кратковременное действие. Собака делалась на станке все более и более вялой и часто совершенно отказывалась от еды после условных раздражителей. Так прошел весь 1925 год. В конце этого года были отменены все старые условные раздражители и начата выработка рефлексов на новые агенты. Это быстро и резко изменило положение дела. Собака на станке снова стала бодрой и еду брала сейчас же, как она подавалась после новых условных раздражителей. Появились постоянные и значительные величины для условного раздражения. Латентный период вернулся к его первоначальному размеру.
Вот один из ранних опытов 21 I 1926 этой новой фазы в работе с собакой. Рефлексы отставлены пока на 15 секунд, и сделана попытка вставить среди новых раздражителей один из старых - удары метронома.
 Разница в величине условных раздражителей отчасти зависит от различной стадии выработки этих новых рефлексов. Достигнутый результат оказывается прочным и дальше.
Общий смысл приведенных фактов бросается в глаза. Ясно, что изолированное условное раздражение, хотя и сопровождаемое потом безусловным раздражителем, действуя на корковую клетку, приводит ее в тормозное состояние тем скорее, чем оно более продолжительное время каждый раз действует на нее, чем чаще повторяется такое действие и в течение более значительного периода времени.
Таким образом не оказывается существенной разницы между развитием тормозного процесса, как мы изучили его в ранних лекциях о разных случаях внутреннего торможения и как он проявился в только что описанных условиях. Разница эта, очевидно, только количественная, но большей частью значительная. В тех случаях, когда условный раздражитель оставался без подкрепления, тормозной процесс выступал очень скоро; здесь же при подкреплении появление тормозного процесса обыкновенно сильно отодвигалось во времени, а иногда и чрезвычайно опазбывало, так что развития его можно было бы даже и не подозревать. Только в некоторых случаях быстрота развития тормозного процесса в обоих рядах опытов почти или вполне сравнивалась. Когда известный положительный условный раздражитель уже обнаружил наклонность к превращению в тормозной, то после небольшого перерыва в работе или даже в начале каждого опыта, в первый раз, имея значительный эффект, он при повторении быстро теряет его, становясь почти или совершенно тормозным, несмотря на подкрепление каждый раз.
Теперь перед нами встает интересный вопрос: каким образом безусловный раздражитель, присоединяемый к условному, обыкновенно отсрочивает развитие тормозного процесса? В лекции об условиях образования условных рефлексов мы видели, что рефлекс не образуется, если безусловный раздражитель предшествует индифферентному агенту, из которого мы намереваемся выработать условный раздражитель. Факт всего естественнее понимать как следствие внешнего торможения, т. е. что сильный пункт безусловного раздражителя ведет к торможению слабейшего пункта индифферентного раздражителя. Тогда позволительно спросить себя: что же делается с пунктом выработанного условного раздражителя, когда к нему присоединяется безусловный раздражитель? Не происходит ли и теперь то же самое? Ввиду этого соображения мы выработанный условный раздражитель, как мы выражаемся, покрывали безусловным раздражителем, т. е. начинали с безусловного раздражителя, и, когда действие его обнаруживалось, присоединяли условной. Такую процедуру продолжали недели, а иногда и месяцы. Опыты были проделаны на многих собаках. Результат получился отчетливый. После этого условный рефлекс не оставался в прежней силе, а довольно часто ослаблялся или исчезал совершенно. Чем были сильнее и более практиковались условные раздражители, тем медленнее происходила при этом потеря ими их действия. Слабые или недавно образованные рефлексы теряли свое действие быстро.
Вот один из таких опытов.
У собаки (опыты Соловейчика) имеются многое и вообще очень большие условные пищевые рефлексы, до 20 капель за 30 секунд. На шипение вырабатывается новый рефлекс. Рефлекс образовался очень скоро. Сочетаний безусловным было всего 11. С шестого сочетания рефлекс стал постоянным и представлял следующие величины по порядку: 10, 8, 13, 9, 9 и 10.5 капли за 30 секунд. Затем этот рефлекс покрывается безусловным рефлексом в течение 32 дней, всего 54 раза.
Вот как шел после этого опыт при первой пробе шипения на его условное действие.
Разница в величине условных раздражителей отчасти зависит от различной стадии выработки этих новых рефлексов. Достигнутый результат оказывается прочным и дальше.
Общий смысл приведенных фактов бросается в глаза. Ясно, что изолированное условное раздражение, хотя и сопровождаемое потом безусловным раздражителем, действуя на корковую клетку, приводит ее в тормозное состояние тем скорее, чем оно более продолжительное время каждый раз действует на нее, чем чаще повторяется такое действие и в течение более значительного периода времени.
Таким образом не оказывается существенной разницы между развитием тормозного процесса, как мы изучили его в ранних лекциях о разных случаях внутреннего торможения и как он проявился в только что описанных условиях. Разница эта, очевидно, только количественная, но большей частью значительная. В тех случаях, когда условный раздражитель оставался без подкрепления, тормозной процесс выступал очень скоро; здесь же при подкреплении появление тормозного процесса обыкновенно сильно отодвигалось во времени, а иногда и чрезвычайно опазбывало, так что развития его можно было бы даже и не подозревать. Только в некоторых случаях быстрота развития тормозного процесса в обоих рядах опытов почти или вполне сравнивалась. Когда известный положительный условный раздражитель уже обнаружил наклонность к превращению в тормозной, то после небольшого перерыва в работе или даже в начале каждого опыта, в первый раз, имея значительный эффект, он при повторении быстро теряет его, становясь почти или совершенно тормозным, несмотря на подкрепление каждый раз.
Теперь перед нами встает интересный вопрос: каким образом безусловный раздражитель, присоединяемый к условному, обыкновенно отсрочивает развитие тормозного процесса? В лекции об условиях образования условных рефлексов мы видели, что рефлекс не образуется, если безусловный раздражитель предшествует индифферентному агенту, из которого мы намереваемся выработать условный раздражитель. Факт всего естественнее понимать как следствие внешнего торможения, т. е. что сильный пункт безусловного раздражителя ведет к торможению слабейшего пункта индифферентного раздражителя. Тогда позволительно спросить себя: что же делается с пунктом выработанного условного раздражителя, когда к нему присоединяется безусловный раздражитель? Не происходит ли и теперь то же самое? Ввиду этого соображения мы выработанный условный раздражитель, как мы выражаемся, покрывали безусловным раздражителем, т. е. начинали с безусловного раздражителя, и, когда действие его обнаруживалось, присоединяли условной. Такую процедуру продолжали недели, а иногда и месяцы. Опыты были проделаны на многих собаках. Результат получился отчетливый. После этого условный рефлекс не оставался в прежней силе, а довольно часто ослаблялся или исчезал совершенно. Чем были сильнее и более практиковались условные раздражители, тем медленнее происходила при этом потеря ими их действия. Слабые или недавно образованные рефлексы теряли свое действие быстро.
Вот один из таких опытов.
У собаки (опыты Соловейчика) имеются многое и вообще очень большие условные пищевые рефлексы, до 20 капель за 30 секунд. На шипение вырабатывается новый рефлекс. Рефлекс образовался очень скоро. Сочетаний безусловным было всего 11. С шестого сочетания рефлекс стал постоянным и представлял следующие величины по порядку: 10, 8, 13, 9, 9 и 10.5 капли за 30 секунд. Затем этот рефлекс покрывается безусловным рефлексом в течение 32 дней, всего 54 раза.
Вот как шел после этого опыт при первой пробе шипения на его условное действие.
 Таким образом значение присоединения безусловного раздражителя в отношении выработанного условного раздражения состоит в том, что оно прекращает деятельное состояние (состояние раздражения) клетки, делая ее на время невосприимчивой к своему раздражителю. Но это еще разъясняет деталей вопроса: как именно происходит при этом количественная разница между развитием торможения при повторении условного раздражителя без подкрепления и при подкреплении. Если сложить все время действия изолированного положительного условного раздражителя при подкреплении до превращения его в тормозной и время действия этого же раздражителя без подкрепления, например при выработке дифференцировки, также до полного развития торможения, то последнее время в подавляюшем числе случаев окажется несоизмеримо меньше первого. Следовательно, здесь должно иметь определяющее значение не простое сокращение всей продолжительности раздраженного состояния клетки, а что-то другое. Может быть, дело в том, что клетка каждый отдельный раз при раздражении без подкрепления остается более продолжительное время в состоянии раздражения, так как в этом случае и по окончании раздражения раздражительный процесс в клетке продолжается, тогда как при подкреплении он кончается с началом подкрепления. Может быть, также имеет важность специально то, что один раз раздражительный процесс обрывается сразу, а в другой раз затухает только постепенно. Возможны и другие предположения. Следовательно, требуется добавочное исследование.
Итак, основной факт, постоянно повторявшийся перед нами, это переход корковой клетки под влиянием условного раздражителя рано или поздно в тормозное состояние. Ввиду всего сообщенного всего естественнее, конечно, рассматривать этот переход в связи с функциональным разрушением клетки при работе, вызываемой раздражением. Это разрушение вообще связано, очевидно, с продолжительностью работы. Но, с другой стороны, ясно, что торможение не есть само разрушиние клетки, так как торможение, возникающее в работавшей клетке, распространяется на клетки, не работавшие, не разрушавшиеся функционально. Быстрота, с которой корковая клетка без подкрепления переходит в тормозное состояние, совпадает, с одной стороны, с ее высшей реактивностью, с другой - с ее, так сказать, требовательностью относительно ее питания. Давно известно, что прекращение кровообращения в коре скорее, чем во всех других тканях тела, ведет к окончательному необратимому нарушению ее функции. И это вполне согласуется физиологической ролью коры как сигнального аппарата. В художественное совершенство машины превращается только что установленный факт, что сигнализируемый безусловный раздражитель на время его действия делает корковую клетку невозбудимой. Позволю себе повторить раньше мной употребленную фразу. Образцово бдительнейший сигнальщик исполнил свою ответственную роль, и его отдых, восстановление должны быть сейчас же ему обеспечены ради такой же исправности будущей его работы.
Остается еще вопрос: есть ли такой минимальный срок действия изолированного условного раздражителя, который не влечет за собой в клетке нарастающей тенденции к тормозному состоянию? Изучением этого пункта мы заняты в настоящее время. И у нас уже есть опыты, находящиеся, может быть, в связи с этим пунктом. У собак, имевших долгое время отставленные на 30 секунд и еще хорошо сохранившиеся условные рефлексы, мы все эти рефлексы делали почти совпадающими, т. е. подкрепляли через 1-2 секунды после начала условного раздражителя. Это, казалось бы, небольшое изменение процедуры применения уже выработанных условных рефлексов сейчас же начинает отражаться на их как секреторном, так и двигательнсм компонентах, а также на установившемся балансе между положительными и отрицательными условными раздражителями. Положительные условные рефлексы резко усиливаются, а отрицательные значительно нарушаются, т. ерет перевес раздражительный процесс (опыты Петровой и Крепса).
Вот как опыты велись у одной собаки (опыты Крепса). У У этой собаки, ввиду ее особенности (сильной тормозимости), работа производилась на полу, а не на станке. Между другими условными рефлексами у нее имелся положительный пищевой рефлекс на удары метронома --- 132 удара в минуту с дифференцировкой на 144 удара в минуту. Дифференцировка была полной и прочной. Ранее также был выработан, но давно не применялся условный тормоз - вспыхивание лампочки - к положительному метроному. Условное торможение тоже было полным. Рефлексы постоянно отставлялись на 30 секунд. Теперь рефлексы были переделаны на совпадающие: еда подавалась собаке через секунду после начала действия условного раздражителя. Это повлекло за собой то, что условный тормоз не удалось восстановить в течение 36 дней, несмотря на 100 повторений тормозной комбинации. Точно так же и испытанная после того дифференцировка, полная перед переходом к совпадающему рефлексу, растормозилась и в течение 13 дней при 33 повторениях продолжала быть неудовлетворительной (3 капли при 6-7 каплях на положительный раздражитель). Тогда решено было вернуться к отставленным на 30 секунд рефлексам, и в первый же день дифференцировка снова стала полной. Теперь примененный условный тормоз так же быстро начал вырабатываться и к четвертому дню оказался полным. Приведу пример с условным тормозом.
Таким образом значение присоединения безусловного раздражителя в отношении выработанного условного раздражения состоит в том, что оно прекращает деятельное состояние (состояние раздражения) клетки, делая ее на время невосприимчивой к своему раздражителю. Но это еще разъясняет деталей вопроса: как именно происходит при этом количественная разница между развитием торможения при повторении условного раздражителя без подкрепления и при подкреплении. Если сложить все время действия изолированного положительного условного раздражителя при подкреплении до превращения его в тормозной и время действия этого же раздражителя без подкрепления, например при выработке дифференцировки, также до полного развития торможения, то последнее время в подавляюшем числе случаев окажется несоизмеримо меньше первого. Следовательно, здесь должно иметь определяющее значение не простое сокращение всей продолжительности раздраженного состояния клетки, а что-то другое. Может быть, дело в том, что клетка каждый отдельный раз при раздражении без подкрепления остается более продолжительное время в состоянии раздражения, так как в этом случае и по окончании раздражения раздражительный процесс в клетке продолжается, тогда как при подкреплении он кончается с началом подкрепления. Может быть, также имеет важность специально то, что один раз раздражительный процесс обрывается сразу, а в другой раз затухает только постепенно. Возможны и другие предположения. Следовательно, требуется добавочное исследование.
Итак, основной факт, постоянно повторявшийся перед нами, это переход корковой клетки под влиянием условного раздражителя рано или поздно в тормозное состояние. Ввиду всего сообщенного всего естественнее, конечно, рассматривать этот переход в связи с функциональным разрушением клетки при работе, вызываемой раздражением. Это разрушение вообще связано, очевидно, с продолжительностью работы. Но, с другой стороны, ясно, что торможение не есть само разрушиние клетки, так как торможение, возникающее в работавшей клетке, распространяется на клетки, не работавшие, не разрушавшиеся функционально. Быстрота, с которой корковая клетка без подкрепления переходит в тормозное состояние, совпадает, с одной стороны, с ее высшей реактивностью, с другой - с ее, так сказать, требовательностью относительно ее питания. Давно известно, что прекращение кровообращения в коре скорее, чем во всех других тканях тела, ведет к окончательному необратимому нарушению ее функции. И это вполне согласуется физиологической ролью коры как сигнального аппарата. В художественное совершенство машины превращается только что установленный факт, что сигнализируемый безусловный раздражитель на время его действия делает корковую клетку невозбудимой. Позволю себе повторить раньше мной употребленную фразу. Образцово бдительнейший сигнальщик исполнил свою ответственную роль, и его отдых, восстановление должны быть сейчас же ему обеспечены ради такой же исправности будущей его работы.
Остается еще вопрос: есть ли такой минимальный срок действия изолированного условного раздражителя, который не влечет за собой в клетке нарастающей тенденции к тормозному состоянию? Изучением этого пункта мы заняты в настоящее время. И у нас уже есть опыты, находящиеся, может быть, в связи с этим пунктом. У собак, имевших долгое время отставленные на 30 секунд и еще хорошо сохранившиеся условные рефлексы, мы все эти рефлексы делали почти совпадающими, т. е. подкрепляли через 1-2 секунды после начала условного раздражителя. Это, казалось бы, небольшое изменение процедуры применения уже выработанных условных рефлексов сейчас же начинает отражаться на их как секреторном, так и двигательнсм компонентах, а также на установившемся балансе между положительными и отрицательными условными раздражителями. Положительные условные рефлексы резко усиливаются, а отрицательные значительно нарушаются, т. ерет перевес раздражительный процесс (опыты Петровой и Крепса).
Вот как опыты велись у одной собаки (опыты Крепса). У У этой собаки, ввиду ее особенности (сильной тормозимости), работа производилась на полу, а не на станке. Между другими условными рефлексами у нее имелся положительный пищевой рефлекс на удары метронома --- 132 удара в минуту с дифференцировкой на 144 удара в минуту. Дифференцировка была полной и прочной. Ранее также был выработан, но давно не применялся условный тормоз - вспыхивание лампочки - к положительному метроному. Условное торможение тоже было полным. Рефлексы постоянно отставлялись на 30 секунд. Теперь рефлексы были переделаны на совпадающие: еда подавалась собаке через секунду после начала действия условного раздражителя. Это повлекло за собой то, что условный тормоз не удалось восстановить в течение 36 дней, несмотря на 100 повторений тормозной комбинации. Точно так же и испытанная после того дифференцировка, полная перед переходом к совпадающему рефлексу, растормозилась и в течение 13 дней при 33 повторениях продолжала быть неудовлетворительной (3 капли при 6-7 каплях на положительный раздражитель). Тогда решено было вернуться к отставленным на 30 секунд рефлексам, и в первый же день дифференцировка снова стала полной. Теперь примененный условный тормоз так же быстро начал вырабатываться и к четвертому дню оказался полным. Приведу пример с условным тормозом.
 При переходе еще раз к совпадающим рефлексам условный тормоз уже только после троекратного применения их почти нацело растормозился.
Нужно ли видеть в описанном факте указание, что при стремительной смене условного раздражителя безусловным совсем не будет нарастающего торможения при повторении таких условных рефлексов, или это временное явление, которое, хотя и поздно, все же сменится торможением. - остается пока для нас вопросом.
Но рядом с функциональным разрушением клетки идет, конечно, и ее восстановление, а с ним в некоторых случаях на основании связи истощения клетки с торможением должно исчезать и торможение. Так можно понимать факт, что после угашения условный рефлекс через известный срок сам собой возвращается к нормальному размеру своего положительного действия. С этой точки зрения легко представить себе, что и медленно нарастающее торможение многократновторяемых, хотя и постоянно подкрепляемых, условных рефлексов при переходе к короткому отставлению и при полном перерыве опытов на известное время сменяется возвратом условных рефлексов к норме. С одной стороны, укорачивая время изолированного условного раздражения, с другой - в течение известного срока совсем не вызывая процесса раздражения, мы тем ограничиваем разрушительный процесс в клетке и даем перевес восстановительному.
Мы приступили сейчас к специальному изучению восстановления условных рефлексов после угасания и при нарастающем торможении их, несмотря на подкрепление.
Вот один из относящихся сюда опытов (А. Д. Сперанского). У собаки среди многих положительных и отрицательных условных пищевых рефлексов имелись положительное рефлексы: на удары метронома, усиление общего освещения комнаты, свисток и появление перед глазами собаки фигуры круга. На звуковые раздражения рефлексы были постоянно больше (10--12 капель за 30 секунд), чем на зрительные (6-8 капель). Раздражения применялись в только что приведенном порядке и через один и тот же промежуток -- 10 минут. В каждом опыте ряд раздражителей повторяется один, два раза. После обычного опыта в указанном виде на следующий день весь опыт ведется только с одним из раздражений - ударами метронома. Раздражения, всякий раз подкрепляемые и применяемые также через десятиминутные промежутки, повторяются 12 раз. Первые дза раздражения дают 12 и 11 капель слюны, последние два - по 9 капель. Таким образом повторение раздражения повело к уменьшению его эффекта на 25%. На следующий за этим день опыт ведется так же, с тем только различием, что промежуток между раздражениями очень укорачивается, составляя всего 1.5 минуты. Это было возможно потому, что у данной собаки слюноотделение после еды чрезвычайно быстро обрывалось. Полный перерыв слюноотделения был вполне гарантирован, как показывали постоянные и многочисленные наблюдения. Теперь дело пошло совсем иначе. Рефлекс, начинаясь с 11 капель, при третьем раздражении составлял только 4.5 капли (уменьшение на 60 %), а к восьмому раздражению снизился до 2 капель (уменьшение на 88%). А затем он представляет довольно правильные волны с колебаниями от 2 капель до 5 и 7, дошедши к два цать второму повторению до капли и, наконец, до нуля, причем в последние три раза собака уже не брала подаваемой ей еды. На примененное же теперь через тот же промежуток усиленное освещение комнаты собака дала 2.5 капли слюны и начала есть, а на том же станке и тот же хлеб (большую порцию) без условного раздражителя съела с большой жадностью. В ближайший день, при восстановлении первоначального порядка раздражителей и при промежутке между ними в 10 минут, метроном на первом месте дал 6.5 капли, при повторении на пятом месте после трех других раздражителей только 1.5 капли и, наконец, на девятом месте опять после других раздражителей оказался нулевым, между тем как другие раздражители, хотя тоже уменьшенные против обычного размера, при повторении совершенно не обнаружили такого исключительного падения. Наконец еще на следующий день метроном достиг нормальной величины своего действия и не представлял уже при повторении такой наклонности к падению.
Опыт чрезвычайно содержательный. Мы видим, как один и тот же условный раздражитель, постоянно подкрепляемый и много раз повторяемый через большие промежутки, теряет в своем эффекте только очень мало. Он же при малых промежутках сперва уменьшается стремительно, затем эффект принимает волнообразный характер, и все кончается нулем эффекта, как секреторного, так и двигательного (собака более не берет еды вслед за условным раздражителем). Другой же, обыкновенно более слабый, раздражитель сейчас же вызывает и ту и другую реакцию. Что в этом результате не играет роли насыщение животного, доказывает последний факт и то, что собака на станке же без условного раздражителя с жадностью съедает большую порцию той же еды. Относя эффект ударов метронома к состоянию корковой клетки, надо, следовательно, заключить, что она при частом ее раздражении, функционально истрачиваясь и не успевая восстановляться, переходит в полное тормозное состояние, предварительно представляя колебания, род борьбы между процессами раздражения и торможения. Что здесь мы имеем дело с обычным торможением, указано выше, в начале этой лекции, и вполне подтверждается здесь. Другой раздражитель, более слабый, после полного отказа от действия сильного хотя и действует, но в уменьшенном размере. То же повторяется и на другой день. Клетка, к которой адресовались удары метронома, все еще остается ослабленной и быстро при немногих повторениях идет к полному торможению. И это ее состояние, как это постоянно наблюдается при торможении, иррадиирует на другие клетки, пункты приложения других условных раздражителей, что выражается в уменьшении их эффекта. Но что это есть иррадиированное, так сказать, заимствованное, торможение, a не обусловленное собственной работой, собственным истощением, доказывается тем, что эти другие раздражители не обнаруживают такого быстрого необычного уменьшения их эффекта при повторении, как это происходит с ударами метронома. Еще через день метрономная клетка уже почти вполне восстановляется.
Помимо вопросов: в какой срок при разных раздражителях и когда именно, при полном ли покое клеток или же и во время их работы, а также в какой мере при подкреплении условных рефлексов безусловными и т. д. происходит восстановление, перед нами стоят более глубокие и более трудные вопросы. олько часть случаев наступления исчезания торможения могла бы быть просто понимаема в связи с разрушением и восстановлением клеток. Но как представлять себе другую категорию случаев торможений постоянных или длительных, которыми переполнена деятельность коры? Почему выработанный условный отрицательный раздражитель вызывает в клетке прямо тормозной процесс без предварительного процесса раздражения? Почему клетка, являющаяся пунктом приложения такого раздражителя, не подвергаясь недели, а иногда и месяцы раздражению, не становится сама по себе снова положительно раздражимой?
При переходе еще раз к совпадающим рефлексам условный тормоз уже только после троекратного применения их почти нацело растормозился.
Нужно ли видеть в описанном факте указание, что при стремительной смене условного раздражителя безусловным совсем не будет нарастающего торможения при повторении таких условных рефлексов, или это временное явление, которое, хотя и поздно, все же сменится торможением. - остается пока для нас вопросом.
Но рядом с функциональным разрушением клетки идет, конечно, и ее восстановление, а с ним в некоторых случаях на основании связи истощения клетки с торможением должно исчезать и торможение. Так можно понимать факт, что после угашения условный рефлекс через известный срок сам собой возвращается к нормальному размеру своего положительного действия. С этой точки зрения легко представить себе, что и медленно нарастающее торможение многократновторяемых, хотя и постоянно подкрепляемых, условных рефлексов при переходе к короткому отставлению и при полном перерыве опытов на известное время сменяется возвратом условных рефлексов к норме. С одной стороны, укорачивая время изолированного условного раздражения, с другой - в течение известного срока совсем не вызывая процесса раздражения, мы тем ограничиваем разрушительный процесс в клетке и даем перевес восстановительному.
Мы приступили сейчас к специальному изучению восстановления условных рефлексов после угасания и при нарастающем торможении их, несмотря на подкрепление.
Вот один из относящихся сюда опытов (А. Д. Сперанского). У собаки среди многих положительных и отрицательных условных пищевых рефлексов имелись положительное рефлексы: на удары метронома, усиление общего освещения комнаты, свисток и появление перед глазами собаки фигуры круга. На звуковые раздражения рефлексы были постоянно больше (10--12 капель за 30 секунд), чем на зрительные (6-8 капель). Раздражения применялись в только что приведенном порядке и через один и тот же промежуток -- 10 минут. В каждом опыте ряд раздражителей повторяется один, два раза. После обычного опыта в указанном виде на следующий день весь опыт ведется только с одним из раздражений - ударами метронома. Раздражения, всякий раз подкрепляемые и применяемые также через десятиминутные промежутки, повторяются 12 раз. Первые дза раздражения дают 12 и 11 капель слюны, последние два - по 9 капель. Таким образом повторение раздражения повело к уменьшению его эффекта на 25%. На следующий за этим день опыт ведется так же, с тем только различием, что промежуток между раздражениями очень укорачивается, составляя всего 1.5 минуты. Это было возможно потому, что у данной собаки слюноотделение после еды чрезвычайно быстро обрывалось. Полный перерыв слюноотделения был вполне гарантирован, как показывали постоянные и многочисленные наблюдения. Теперь дело пошло совсем иначе. Рефлекс, начинаясь с 11 капель, при третьем раздражении составлял только 4.5 капли (уменьшение на 60 %), а к восьмому раздражению снизился до 2 капель (уменьшение на 88%). А затем он представляет довольно правильные волны с колебаниями от 2 капель до 5 и 7, дошедши к два цать второму повторению до капли и, наконец, до нуля, причем в последние три раза собака уже не брала подаваемой ей еды. На примененное же теперь через тот же промежуток усиленное освещение комнаты собака дала 2.5 капли слюны и начала есть, а на том же станке и тот же хлеб (большую порцию) без условного раздражителя съела с большой жадностью. В ближайший день, при восстановлении первоначального порядка раздражителей и при промежутке между ними в 10 минут, метроном на первом месте дал 6.5 капли, при повторении на пятом месте после трех других раздражителей только 1.5 капли и, наконец, на девятом месте опять после других раздражителей оказался нулевым, между тем как другие раздражители, хотя тоже уменьшенные против обычного размера, при повторении совершенно не обнаружили такого исключительного падения. Наконец еще на следующий день метроном достиг нормальной величины своего действия и не представлял уже при повторении такой наклонности к падению.
Опыт чрезвычайно содержательный. Мы видим, как один и тот же условный раздражитель, постоянно подкрепляемый и много раз повторяемый через большие промежутки, теряет в своем эффекте только очень мало. Он же при малых промежутках сперва уменьшается стремительно, затем эффект принимает волнообразный характер, и все кончается нулем эффекта, как секреторного, так и двигательного (собака более не берет еды вслед за условным раздражителем). Другой же, обыкновенно более слабый, раздражитель сейчас же вызывает и ту и другую реакцию. Что в этом результате не играет роли насыщение животного, доказывает последний факт и то, что собака на станке же без условного раздражителя с жадностью съедает большую порцию той же еды. Относя эффект ударов метронома к состоянию корковой клетки, надо, следовательно, заключить, что она при частом ее раздражении, функционально истрачиваясь и не успевая восстановляться, переходит в полное тормозное состояние, предварительно представляя колебания, род борьбы между процессами раздражения и торможения. Что здесь мы имеем дело с обычным торможением, указано выше, в начале этой лекции, и вполне подтверждается здесь. Другой раздражитель, более слабый, после полного отказа от действия сильного хотя и действует, но в уменьшенном размере. То же повторяется и на другой день. Клетка, к которой адресовались удары метронома, все еще остается ослабленной и быстро при немногих повторениях идет к полному торможению. И это ее состояние, как это постоянно наблюдается при торможении, иррадиирует на другие клетки, пункты приложения других условных раздражителей, что выражается в уменьшении их эффекта. Но что это есть иррадиированное, так сказать, заимствованное, торможение, a не обусловленное собственной работой, собственным истощением, доказывается тем, что эти другие раздражители не обнаруживают такого быстрого необычного уменьшения их эффекта при повторении, как это происходит с ударами метронома. Еще через день метрономная клетка уже почти вполне восстановляется.
Помимо вопросов: в какой срок при разных раздражителях и когда именно, при полном ли покое клеток или же и во время их работы, а также в какой мере при подкреплении условных рефлексов безусловными и т. д. происходит восстановление, перед нами стоят более глубокие и более трудные вопросы. олько часть случаев наступления исчезания торможения могла бы быть просто понимаема в связи с разрушением и восстановлением клеток. Но как представлять себе другую категорию случаев торможений постоянных или длительных, которыми переполнена деятельность коры? Почему выработанный условный отрицательный раздражитель вызывает в клетке прямо тормозной процесс без предварительного процесса раздражения? Почему клетка, являющаяся пунктом приложения такого раздражителя, не подвергаясь недели, а иногда и месяцы раздражению, не становится сама по себе снова положительно раздражимой?
Лекция пятнадцатая. Внутреннее торможение и сон - один и тот же процесс в своей физико-химической основе
Мм. гг.! В прошлой лекции мы пришли к очень важному положению, что корковая клетка под влиянием условных раздражений непременно рано или поздно, а при частых повторениях их и очень быстро, приходит в тормозное состояние. А это всего законнее надо было понимать так, что эта клетка как, так сказать, сторожевой пункт организма владеет высшей реактивностью, а следовательно стремительной функциональной разрушаемостью, быстрой утомляемостью. Наступающее тогда торможение, не будучи само утомлением, является в роли охранителя клетки, предупреждающего дальнейшее чрезмерное, опасное разрушение этой исключительной клетки. За время тормозного периода, оставаясвободной от работы, клетка восстановляет свой нормальный состав. Это касается всех клеток коры, и, следовательно, при условии множества работавших клеток коры, вся кора должна приходить в то же тормозное состояние, которое мы видели в отдельных клетках коры, когда на них действовали наши условные раздражители. А это и есть каждодневный факт, есть сон наш и всех животных. Вся наша двадцатипятилетняя работа над большими полушариями является сплошным и постоянным доказательством этого заключения. Сейчас в физиологии больших полушарий, как мы ее изучаем нашим методом, это одно из наиболее прочных положений. Сонливость и сон наших экспериментальных животных сопровождали нашу работу с самого ее начала, и до сих пор мы постоянно имеем с ними дело. Таким образом нами собран огромный фактический материал, который в разные фазы нашего исследования, конечно, давал основание для многих предположений, несколько разнящихся друг от друга. Ное несколько лет тому назад все эти предположения слились в окончательное, гармонирующее решительно со всеми нашими фактами, положение, что сон и то, что мы называем внутренним торможением, есть один и тот же процесс. Основное условие появления и развития как этого торможения, так и сна - совершенно одно и то же. Это более или менее продолжительное или много раз повторяющееся изолированное условное раздражение, т. е. раздражение корковой клетки. При всех случаях внутреннего торможения, с которыми мы знакомились в четвертой, пятой, шестой седьмой лекциях, мы постоянно встречались с сонливостью сном наших животных. Если мы производим угасание, то иного животного и при первом случае с ним, помимо прекращения условного слюноотделения и соответствующей двигательной реакции, уже замечается большая вялость животного сравнительно с состоянием до угасания. При повторении угасания в течение ряда дней, хотя бы при чередовании с подкрепляемыми условными раздражителями, почти всегда дело кончается явной сонливостью и сном на станке, хотя до этого с животным этого не случалось. То же самое еще резче наблюдается при выработке дифференцировок. Мы, например, имеем у животного ряд условных раздражителей и между другими определенный тон. Животное все время остается на станке бодрым. Затем мы переходим к дифференцировке одного из близких тонов. Вместе с начинающим развиваться дифференцировочным торможением появляется сонливость, все усиливающаяся и кончающаяся глубоким сном с полным расслаблением скелетной мускулатуры и храпом животного, так что при следующих положительных условных раздражителях и подаче еды приходится расталкивать животное или даже вкладывать ему пищу в рот для начала акта еды. С тем же самым встречаемся и при образовании сильно запаздывающего отставленного рефлекса (на 3 минуты), так что раньше, когда мы еще мало были знакомы с делом, из-за возникающего при этом сна у иных животных нельзя было добиться выработки нужного нам рефлекса. Наконец то же имеет место и при выработке условного торможения. Надо заметить, что при разных случаях внутреннего торможения степень вмешательства сна несколько разлимна. Всего меньше он дает себя знать именно при условном торможении. Это все случаи, где внутреннее торможение развивается. быстро вследствие того, что условный раздражитель не сопровождается безусловным раздражителем, но то же самое происходит и при медленно нарастающем торможении подкрепляемых, но много раз повторенных условных рефлексов, торможении, о котором специально была речь в прошлой лекции. И тут через. месяцы и годы дело кончается разными переходными состояниями между бодрым состоянием и сном (что составит содержание следующей лекции) и полным сном, смотря по животному. В этом отношении животные так же разнятся между собой, как и в отношении скорости наступления торможения. Случаями перехода торможения в сон так переполнена наша работа с условными рефлексами, что является излишним приводить отдельные примеры. Таким образом действительно торможение тесно связано со сном, прямо переходит в сон, если не принимаются соответствующие меры. Интересно, что как для внутреннего торможения почти нет раздражителя, который бы не подвергся ему при определенных условиях, так и для сна. Сильнейший электрический ток, приложенный к коже в качестве условного пищевого раздражителя, в опытах Ерофеевой обнаружил после многих месяцев нарастающее торможение при подкрепляемых рефлексах, а в опытах Петровой демонстративно вызывал сон. С другой стороны, различные внешние агенты в роли условных раздражителей совершенно одинаково располагаются в отношении скорости развития при них как внутреннего торможения, так и сна. В прошлой лекции было сообщено, что всего легче развивается торможение при температурных раздражениях и всего труднее при звуковых. Строго параллельно с этим сон быстро наступает при температурных условных рефлексах и позже и реже при звуковых. При температурных условных раздражениях вмешательство сна так настойчиво и так нарушает работу, что в первую пору наших исследований трудно было найти сотрудников, соглашавшихся работать с этими раздражителями. Наконец, как раньше было указано, для торможения и для наступления сна определяющее значение имеет продолжительность условного раздражения. У иной собаки, пока условный раздражитель отставлен на 10-15 секунд, за длинный период работы сохраняется совершенно бодрое состояние. Лишь только рефлекс у нее отставляется на 30 секунд и больше, сейчас же появляются сонливость и сон. Эта форма опыта часто поистине поразительна. Кажется совершенно неожиданным этот быстрый переход от полной бодрости к настоящему сну при таком, повидимому, малозначительном изменении условий опыта. Таких примеров при разной длине отставления и с разной степенью отчетливости рассеяно по нашей работе множество. Все без исключения приемы, которые были приведены в прошлой лекции как задерживающие наступление нарастающего торможения давно повторяющихся, хотя и постоянно подкрепляемых условных рефлексов, так и устраняющие торможение, служат одинаково и для борьбы со сном. После всего только что приведенного поднимается естественнейший вопрос: если сон так совпадает в его появлении и в его исчезании с внутренним торможением, то каким образом последнее может быть важнейшим фактором бодрого состояния, основанием тончайшего уравновешивания организма с окружающей средой? Сообщенные в ранних лекциях факты должны, по моему мнению, совершенно устранить кажущуюся, может быть, на первый взгляд трудность ответа. Внутреннее торможение в бодром состоянии есть раздробленный сон, сон отдельных групп клеток, как сон есть внутреннее торможение, иррадиированное, распространившееся сплошь на всю массу полушарий и на лежащие ниже отделы головного мозга. Все дело, следовательно, состоит в пространственном ограничении торможения, в заключении его в определенные рамки. А делает это, конечно, противоположный нервный процесс, как это мы видели в лекциях о мозаичности коры и об анализаторной деятельности ее. При угасании сон не наступает, не допускается только тогда, когда после достигнутого угасания мы систематически применяем подкрепляемые условные раздражители не повторяем угасание слишком часто. При дифференцировках развивающееся торможение, сначала сопровождаемое сном, остается, так сказать, деловым торможением, без осложнения неуместным сном, только тогда, когда вперемежку с неподкрепляемым агентом применяется, и обыкновенно чаще, наш условный раздражитель, положительно действующий агент, т. е. процесс раздражения постоянно противодействует распространению процесса торможения. То же самое - при условном торможении и при запаздывании. Во всех этих случаях, если опыт ведется целесообразно, сонливость и сон являются только как фазовое явление, когда еще не установилось точное разграничение между районами раздражительного и тормозного процессов. Как только условия опыта дают перевес торможению, наступает сон. Вот резкий пример. В лекции о мозаике я упоминал о собаке, у которой на определенный тон был выработан положительный условный рефлекс, a 20 соседних с ним тонов вверх и вниз были втдифференцированы, т. е. были отрицательными условными раздражителями. Собака вообще была очень не склонна ко сну, когда соблюдалось определенное отношение при применении положительных и отрицательных раздражителей, и всегда тогда давала полные эффекты на все положительные раздражители. Но как только несколько раз подряд пускались в действие отдифференцированные тоны, собака скоро впадала в такой глубокий сон, что ее не будили очень сильные посторонние раздражители. Наоборот, сон не наступает, несмотря и на частое применение отрицательных условных раздражений, если они чередуются с положительными. В этом отношении очень поучительны упоминаемые ранее опыты с мозаикой и особенно опыты с кожно-механической мозаикой. Несмотря на то, что условное кожно-механическое раздражение очень располагает к сонливости и сну, тем не менее собака Купалова. на которой более двух лет практикуется исключительно кожно-механическая мозаика, никакой наклонности ко сну не обнаруживает, очевидно потому, что тормозной процесс постоянно отмежевывается, заключается узкие рамки раздражительным процессом. Обширно иррадиирующему торможению мы противодействуем еще и тем, что увеличиваем число положительных условных раздражителей, ограничивающих распространение торможения из его исходников. Приведу более сложный, отчасти сюда относящийся, пример из работы Петровой. У собаки было приступлено сразу к выработке сильно отставленного (на 3 минуты) пищевого рефлекса на удары метронома. Собака очень скоро стала впадать в сонливое состояние и кончила полным сном. Очевидно, торможение, которое должно было развиться на первом периоде действия метронома, как очень удаленном от времени присоединения безусловного раздражителя, взяло перевес, не встречая должного противодействия со стороны еще не проявившегося процесса раздражения на втором периоде действия метронома как непосредственно предшествующем действию безусловного раздражителя. Тогда было введено еще 5 новых агентов для выработки из них условных раздражителей, причем еда присоединялась к ним спустя только 5 секунд после их начала. Сонливое состоянтро исчезло, и все рефлексы легло образовались, затем началось постепенное, каждый день на 5 секунд, отодвигание акта еды от начала условных раздражителей. Соответственно удлинялся и латентный период рефлексов, и в конце, без малейшего вмешательства сна, получилось 6 рефлексов с латентным периодом в 1--1.5 минуты, т. е. с предварительным тормозным процессом. Таким образом процесс раздражения, первоначально возникший в 6 пунктах коры и только постепенно давая место процессу торможения, ограничил его во времени и в пространстве, не допустивши сна. Сюда же надо отнести и влияние на некоторых, правда немногих, собак, особенно наклонных торможению и сонливости, ограничения движений в нашем станке. Тогда иногда целесообразно вести опыты, по крайней мере временно, на полу, на свободе. Надо думать, что раздражения, идущие при движении из двигательного аппарата и с кожи, образуют переменно возникающие очаги возбуждения в коре полушарий, которые также до известной степени противодействуют распространению торможения, хотя тут есть и другой момент, вероятно большего значения, о чем речь впереди. То, что мы видели на корковой клетке относительно появления в ней торможения в случаях условного раздражения, повторяется на ней и при раздражении ее всяким агентом, не имеющим никакого условного специального физиологического значения. Как уже упомянуто раньше, между рефлексами имеется исследовательский рефлекс. Этот рефлекс имеет своими пунктами приложения клетки как больших полушарий, так и ниже лежащих отделов головного мозга. При наличии больших полушарий он, очевидно, происходит при участии клеток больших полушарий. Это несомненно доказывается его высочайшей тонкостью, когда он появляется при всяком малейшем колебании окружающей среды, что возможно только при высшей анализаторной функции больших полушарий и что совершенно недоступно для ниже лежащих отделов мозга. Исследовательский рефлекс, как все мы знаем, при повторении непременно слабеет и, наконец, совершенно исчезает, несмотря на продолжение существования агента, его вызвавшего. Специальные опыты в нашей лаборатории Н. А. Попова показали, что исчезание исследовательского рефлекса основано на развитии торможения и вполне аналогично в отношении подробностей угасанию условных рефлексов. Если повторяемый через короткие промежутки времени в течение опыта данный агент исследовательского рефлекса перестает вызывать соответствующую двигательную реакцию, то при большем промежутке в том же опыте эта реакция восстанавливается, как это делается с угашенным условным рефлексом. Исчезнувший в опыте от повторения определенный исследовательский рефлекс на время появляется, если сейчас же после его исчезания применяется новый посторонний раздражитель, т. е. производится другой исследовательский рефлекс. Следовательно, он растормаживается, как и задержанный условный рефлекс. Если исследовательский рефлекс на определенный агент повторяется в течение ряда дней, то он хронически исчезает как систематически неподкрепляемый условный рефлекс. Наконец исчезнувший исследовательский рефлекс временио восстанавливается под влиянием возбуждающих веществ (кофеина), как это наблюдается, например, при условных дифференцированных агентах. Это торможение исследовательского рефлекса непременно, и даже легче, чем торможение условного рефлекса, ведет к сонливости и сну. Привожу опыты из работы С. И. Чечулина, в которой торможение и сон при исследовательском рефлексе комбинировались с применением условных раздражителей. У собаки выработан пищевой условный рефлекс на свисток. В качестве возбудителей исследовательского рефлекса употребляются впервые применяемые агенты: шипение, бульканье, механическое раздражение кожи и другие. Затем, начиная с двадцать первой минуты, применяется в течение 30 секунд и повторяется через каждые 2 минуты звук бульканья. При первых трех повторениях наступают движения в сторону звука, постепенно ослабляющиеся. С четвертого обнаруживается сонливость. До восьмого повторения сон прерывается на разные моменты раздражения. При восьмом и девятом нег более никаких движений на раздражитель, сон сплошной. В 43 минуты сперва одно бульканье применяется 10 секунд, а затем на 30 секунд к нему присоединяется свисток. Нет никакой - ни двигательной, ни секреторной реакции, продолжается сон. Подача еды пробуждает собаку, собака начинает есть, но и после еды остается сонливой. Опыт с условным раздражителем продолжается следующим образом.
Затем, начиная с двадцать первой минуты, применяется в течение 30 секунд и повторяется через каждые 2 минуты звук бульканья. При первых трех повторениях наступают движения в сторону звука, постепенно ослабляющиеся. С четвертого обнаруживается сонливость. До восьмого повторения сон прерывается на разные моменты раздражения. При восьмом и девятом нег более никаких движений на раздражитель, сон сплошной. В 43 минуты сперва одно бульканье применяется 10 секунд, а затем на 30 секунд к нему присоединяется свисток. Нет никакой - ни двигательной, ни секреторной реакции, продолжается сон. Подача еды пробуждает собаку, собака начинает есть, но и после еды остается сонливой. Опыт с условным раздражителем продолжается следующим образом.
 Собака до этого опыта в станке никогда нпала. В следующих опытах новые агенты, вызывающиеследовательский рефлекс, то повторялись до наступления сна, то только до тех пор, пока не исчезали на них движения, но еще до появления сонливости. 21 день спустя после вышеприведенного опыта ставится опыт c механическим раздражением кожи. Опыт идет следующим образом.
Собака до этого опыта в станке никогда нпала. В следующих опытах новые агенты, вызывающиеследовательский рефлекс, то повторялись до наступления сна, то только до тех пор, пока не исчезали на них движения, но еще до появления сонливости. 21 день спустя после вышеприведенного опыта ставится опыт c механическим раздражением кожи. Опыт идет следующим образом.
 С двадцать пятой минуты начинается механическое раздражение кожи, продолжающееся 30 секунд каждый раз и повторяемое через минуту. При первых трех повторениях -- движения головой в сторону раздражаемого места. При четвертом и пятом повторениях движений больше нет. В 32.5 минуты на 10 секунд применяется одно механическое раздражение кожи, a затем в продолжение 30 секунд к нему присоединяется свисток. На пятнадцатой секунде от начала свистка начинается слюноотделение, и за 30 секунд присоединения свистка выделяются 2 капли. Дальше опыт идет так.
С двадцать пятой минуты начинается механическое раздражение кожи, продолжающееся 30 секунд каждый раз и повторяемое через минуту. При первых трех повторениях -- движения головой в сторону раздражаемого места. При четвертом и пятом повторениях движений больше нет. В 32.5 минуты на 10 секунд применяется одно механическое раздражение кожи, a затем в продолжение 30 секунд к нему присоединяется свисток. На пятнадцатой секунде от начала свистка начинается слюноотделение, и за 30 секунд присоединения свистка выделяются 2 капли. Дальше опыт идет так.
 В начале опыта условный раздражителреплялся то 5 секунд спустя, то 30, чтобы сохранить нормальную величину его эффекта до конца опыта.
Мы видим, что при повторениях агента, вызывающего исследовательский рефлекс, его двигательный эффект постепенно все уменьшается, a затем при продолжающихся 11овторениях то прямо наступают сонливость и сон, все углубляющиеся, то перед этим агент остается некоторое время как бы без всякого действия. Однако присоединяемый к нему условный раздражитель (во втором из только что приведенных опытов) свидетельствует, что он в этот период производит тормозящее действие. Что тормозящее действие на условный раздражитель не основано теперь на механизме так называемого нами внешнего торможения, доказывается тем, что (как это мы имели в шестой лекции при изучении действия исследовательского рефлекса на запахдывающий условный рефлекс) исследовательский рефлекс, когда он оказывается очень малым, растормаживает, а не тормозит условный рефлекс. Следователь действует уже торможение, развивающееся при повторении исследовательского рефлекса и переходящее потом сон, при котором условный рефлекс и совсем исчезает (в первом из только что приведенных опытов).
То же мы видим в особенности резко выраженным на щенках (опыт И. С. Розенталя). При однообразном повторении какогонибудь раздражителя, при отсутствии вообще колебаний в окружающей среде, щенки часто поразительно точно и быстро впадают в сон. А с другой стороны, разве это не общеизвестная истина, хотя до сих пор остававшаяся без научного освещения, что все люди, а особенно не имеющие сильной внутренней жизни, при однообразных раздражениях, как бы ни было неуместно и несвоевременно, неодолимо впадают в сонливость и сон. Значит, определенные корковые клетки, реагирующие на данный длительно продолжающийся внешний агент, истрачиваясь, переходят в тормозное состояние, а при отсутствии противодействия со стороны других деятельных пунктов коры тормозной процесс, распространяясь, обусловливает сон. Чрезвычайно быстрая истощаемость корковой клетки с переходом клетки в тормозное состояние резко контрастирует с выносливостью клеток низших отделов головного мозга при тех же условиях. Опыты Зеленого в нашей лаборатории показали, что в то время как у нормальной собаки на определенный звук исследовательский рефлекс быстро исчезает, тот же звук при той же обстановке у собаки без больших полушарий вызывает этот рефлекс стереотипно огромное число раз.
Возвращаюсь к условным рефлексам.
Что основание для развития торможения, а за ним и для наступления сна дает истощение корковой клетки и вообще ее ослабление, доказывается и следующими постоянно перед нами повторявшимися фактами. Когда мы хирургачески, операцией на полушарии, нарушали какой-нибудь анализатор, то положительные условные раздражители, к нему относящиеся, или почти нельзя было применять изолированными даже очень короткое время, - они быстро превращались в тормозные, - или даже они совсем теряли их положительное действие, становясь только отрицательными, тормозящими. Это в особенности легко и постоянно наблюдается при нарушении кожного анализатора.
Когда у собаки удаляются gyri coronarius и ectosylvius, то у нее на значительный срок (часто на многое недели) исчезают положительные условные кожно-механические рефлексы с конечностей, с плечевого и тазового поясов туловища, заменяясь отрицательными тормозными. Последнее доказывается тем, что положительные условные раздражители изугих анализаторов хорошо действуют до применения кожно-механического раздражения, теряя свое действие после него. Вместе с этим эти кожномеханические раздражения чрезвычайно легко и скоро вызывают сон у таких собак, которые раньше при тех же раздражениях никогда не спали во время нашей работы с ними. Факт часто принимает следующую выразительную форму. Условный кожномеханический раздражитель на поврежденном мозговой операцией отделе кожной поверхности ведет к тормовению и сонливости, а тот же раздражитель на неповрежденных местах дает положительный эффект и оставляет животное совершенно бодрым (опыты Н. И. Красногорского, И. П. Разенкова и В. М. Архангельского).
Сюда же справедливо отнести и факт, наблюдавшийся в лаборатории в наш голодный период, несколько лет тому назад. На истощенных животных нельзя было вестиисследование с условными рефлексами, так как все положительные условные раздражители чрезвычайно быстро становились у них отрицательными, и вместе с тем собаки начинали спать именно в связи с применением условных раздражителей. Очевидно, общее истощение животного особенно давало себя знать на корковых клетках (опыт Фролова, Розенталя и др.).
В перечисленных до сих пор многочисленных случаях мы постоянно видели, что торможение переходит в сон, но можно наблюдать и обратное - переход сна в торможение. Мы выработали отставленный на 3 минуты условный рефлекс. Отдельный опыт иногда идет следующим образом. Мы ставим животное в станок. Оно бодро. Но как только применяется условный раздражитель, животное сейчас же становится сонным, слюноотделительный эффект отсутствует все 3 минуты, и при подаче еды после этого животное не сразу и вяло ест подаваемую пищу. Мы повторяем в опыте через обыкновенные промежутки условный раздражитель несколько раз. С каждым разом наша собака при раздражении держится все бодрее, в конце 3 минут раздражения появляется слюноотделение. С повторением это слюноотделение все растет. Наконец трехминутный период раздражения делится приблизительно на две половины: в первую нет слюноотделения, хотя животное остается совершенно бодрым, во вторую обильное слюноотделение, и собака стремительно и с жадностью ест подаваемую ей еду. Здесь иррадиированное торможение - сон, наступившее в силу преобладания сначала тормозного процесса при первой половине действия условного раздражителя, постепенно переходит в ограниченное, концентрированное торможение под давлением все усиливающегося процесса раздражения, связанного со второй половиной действия того же условного раздражителя. В подобных случаях приходится видеть и чистую замену торможения сном. Как при продолжительном отставлении на 3 минуты, так и при более коротком, на 30 секунд, иногда наблюдается следующее отношение. Животное, постоянно устающееся на станке во время опыта бодрым, точно каждый раз с началом действия повторяющегося условного раздражителя засыпает: закрываются глаза, опускается голова, все туловище расслабленно виснет на лямках, временами даже слышится храп. Но проходит определенный срок, при длинном отставлении 1.5 2 минуты, при коротком 25 25 секунд, животное быстро само собой просыпается, появляется слюноотделение и обнаруживается резкая пищевая двигательная реакция. Ясно, что в данном случае концентрированное торможение хронически заменяется разлитым сном. Наконец можно иногда видеть, как суммирование двух торможений дает сонливость.
У одной собаки (опыт Фурсикова) имелся хорошо выработанный условный рефлекс, отставленный на 3инуты. Первые 2 минуты не было слюноотделения, затем оно начиналось, достигая максимума к концу третьей минуты. В данном опыте вместе с условным раздражителем применяется посторонний агент слабое шипение. Оно растормаживает первую половину рефлекса, причем на шипение была заметна небольшая двигательная реакция, исследовательский рефлекс. Условный рефлекс был подкреплен. При повторении той же комбинации ориентировочной реакции на шипение больше нет, а условный рефлекс совершенно исчезает, и у животного замечается резкая сонливость. Рефлекс опять подкрепляется. Результат, очевидно, надо понимать так. Исследовательский рефлекс на шипение сорого раза уже гаснет, т. е. теперь на шипение развивается торможение, как об этом сказано раньше в настоящей лекции. Это торможение, складываясь с торможением первой фазы запаздывающего рефлекса, усиливает торможение настолько, что деятельная фаза рефлекса отпадает, уступая место сонливости животного. Что это толкование хода опыта верно, свидетельствует продолжение опыта. При следующем повторении условного раздражения без присоединения шипения получается правильный запаздывающий рефлекс с двумя фазами. Повторенная еще раз комбинация метронома с шипением опять производит сонливость при исчезании условного рефлекса.
Вот подлинные цифры этого опыта.
В начале опыта условный раздражителреплялся то 5 секунд спустя, то 30, чтобы сохранить нормальную величину его эффекта до конца опыта.
Мы видим, что при повторениях агента, вызывающего исследовательский рефлекс, его двигательный эффект постепенно все уменьшается, a затем при продолжающихся 11овторениях то прямо наступают сонливость и сон, все углубляющиеся, то перед этим агент остается некоторое время как бы без всякого действия. Однако присоединяемый к нему условный раздражитель (во втором из только что приведенных опытов) свидетельствует, что он в этот период производит тормозящее действие. Что тормозящее действие на условный раздражитель не основано теперь на механизме так называемого нами внешнего торможения, доказывается тем, что (как это мы имели в шестой лекции при изучении действия исследовательского рефлекса на запахдывающий условный рефлекс) исследовательский рефлекс, когда он оказывается очень малым, растормаживает, а не тормозит условный рефлекс. Следователь действует уже торможение, развивающееся при повторении исследовательского рефлекса и переходящее потом сон, при котором условный рефлекс и совсем исчезает (в первом из только что приведенных опытов).
То же мы видим в особенности резко выраженным на щенках (опыт И. С. Розенталя). При однообразном повторении какогонибудь раздражителя, при отсутствии вообще колебаний в окружающей среде, щенки часто поразительно точно и быстро впадают в сон. А с другой стороны, разве это не общеизвестная истина, хотя до сих пор остававшаяся без научного освещения, что все люди, а особенно не имеющие сильной внутренней жизни, при однообразных раздражениях, как бы ни было неуместно и несвоевременно, неодолимо впадают в сонливость и сон. Значит, определенные корковые клетки, реагирующие на данный длительно продолжающийся внешний агент, истрачиваясь, переходят в тормозное состояние, а при отсутствии противодействия со стороны других деятельных пунктов коры тормозной процесс, распространяясь, обусловливает сон. Чрезвычайно быстрая истощаемость корковой клетки с переходом клетки в тормозное состояние резко контрастирует с выносливостью клеток низших отделов головного мозга при тех же условиях. Опыты Зеленого в нашей лаборатории показали, что в то время как у нормальной собаки на определенный звук исследовательский рефлекс быстро исчезает, тот же звук при той же обстановке у собаки без больших полушарий вызывает этот рефлекс стереотипно огромное число раз.
Возвращаюсь к условным рефлексам.
Что основание для развития торможения, а за ним и для наступления сна дает истощение корковой клетки и вообще ее ослабление, доказывается и следующими постоянно перед нами повторявшимися фактами. Когда мы хирургачески, операцией на полушарии, нарушали какой-нибудь анализатор, то положительные условные раздражители, к нему относящиеся, или почти нельзя было применять изолированными даже очень короткое время, - они быстро превращались в тормозные, - или даже они совсем теряли их положительное действие, становясь только отрицательными, тормозящими. Это в особенности легко и постоянно наблюдается при нарушении кожного анализатора.
Когда у собаки удаляются gyri coronarius и ectosylvius, то у нее на значительный срок (часто на многое недели) исчезают положительные условные кожно-механические рефлексы с конечностей, с плечевого и тазового поясов туловища, заменяясь отрицательными тормозными. Последнее доказывается тем, что положительные условные раздражители изугих анализаторов хорошо действуют до применения кожно-механического раздражения, теряя свое действие после него. Вместе с этим эти кожномеханические раздражения чрезвычайно легко и скоро вызывают сон у таких собак, которые раньше при тех же раздражениях никогда не спали во время нашей работы с ними. Факт часто принимает следующую выразительную форму. Условный кожномеханический раздражитель на поврежденном мозговой операцией отделе кожной поверхности ведет к тормовению и сонливости, а тот же раздражитель на неповрежденных местах дает положительный эффект и оставляет животное совершенно бодрым (опыты Н. И. Красногорского, И. П. Разенкова и В. М. Архангельского).
Сюда же справедливо отнести и факт, наблюдавшийся в лаборатории в наш голодный период, несколько лет тому назад. На истощенных животных нельзя было вестиисследование с условными рефлексами, так как все положительные условные раздражители чрезвычайно быстро становились у них отрицательными, и вместе с тем собаки начинали спать именно в связи с применением условных раздражителей. Очевидно, общее истощение животного особенно давало себя знать на корковых клетках (опыт Фролова, Розенталя и др.).
В перечисленных до сих пор многочисленных случаях мы постоянно видели, что торможение переходит в сон, но можно наблюдать и обратное - переход сна в торможение. Мы выработали отставленный на 3 минуты условный рефлекс. Отдельный опыт иногда идет следующим образом. Мы ставим животное в станок. Оно бодро. Но как только применяется условный раздражитель, животное сейчас же становится сонным, слюноотделительный эффект отсутствует все 3 минуты, и при подаче еды после этого животное не сразу и вяло ест подаваемую пищу. Мы повторяем в опыте через обыкновенные промежутки условный раздражитель несколько раз. С каждым разом наша собака при раздражении держится все бодрее, в конце 3 минут раздражения появляется слюноотделение. С повторением это слюноотделение все растет. Наконец трехминутный период раздражения делится приблизительно на две половины: в первую нет слюноотделения, хотя животное остается совершенно бодрым, во вторую обильное слюноотделение, и собака стремительно и с жадностью ест подаваемую ей еду. Здесь иррадиированное торможение - сон, наступившее в силу преобладания сначала тормозного процесса при первой половине действия условного раздражителя, постепенно переходит в ограниченное, концентрированное торможение под давлением все усиливающегося процесса раздражения, связанного со второй половиной действия того же условного раздражителя. В подобных случаях приходится видеть и чистую замену торможения сном. Как при продолжительном отставлении на 3 минуты, так и при более коротком, на 30 секунд, иногда наблюдается следующее отношение. Животное, постоянно устающееся на станке во время опыта бодрым, точно каждый раз с началом действия повторяющегося условного раздражителя засыпает: закрываются глаза, опускается голова, все туловище расслабленно виснет на лямках, временами даже слышится храп. Но проходит определенный срок, при длинном отставлении 1.5 2 минуты, при коротком 25 25 секунд, животное быстро само собой просыпается, появляется слюноотделение и обнаруживается резкая пищевая двигательная реакция. Ясно, что в данном случае концентрированное торможение хронически заменяется разлитым сном. Наконец можно иногда видеть, как суммирование двух торможений дает сонливость.
У одной собаки (опыт Фурсикова) имелся хорошо выработанный условный рефлекс, отставленный на 3инуты. Первые 2 минуты не было слюноотделения, затем оно начиналось, достигая максимума к концу третьей минуты. В данном опыте вместе с условным раздражителем применяется посторонний агент слабое шипение. Оно растормаживает первую половину рефлекса, причем на шипение была заметна небольшая двигательная реакция, исследовательский рефлекс. Условный рефлекс был подкреплен. При повторении той же комбинации ориентировочной реакции на шипение больше нет, а условный рефлекс совершенно исчезает, и у животного замечается резкая сонливость. Рефлекс опять подкрепляется. Результат, очевидно, надо понимать так. Исследовательский рефлекс на шипение сорого раза уже гаснет, т. е. теперь на шипение развивается торможение, как об этом сказано раньше в настоящей лекции. Это торможение, складываясь с торможением первой фазы запаздывающего рефлекса, усиливает торможение настолько, что деятельная фаза рефлекса отпадает, уступая место сонливости животного. Что это толкование хода опыта верно, свидетельствует продолжение опыта. При следующем повторении условного раздражения без присоединения шипения получается правильный запаздывающий рефлекс с двумя фазами. Повторенная еще раз комбинация метронома с шипением опять производит сонливость при исчезании условного рефлекса.
Вот подлинные цифры этого опыта.
 При этом случае считаю не бесполезным ради ясности нарочито обратить ваше внимание на следующее. Очевидно, этот опыт вместе с вышеприведенными опытами Чечулина прибавляет лишнюю новую фазу в действии посторонних агентов на условный рефлекс. Если взять сильный внешний агент, то он, как вы помните из шестой лекции запаздывающем рефлексе, вызываемым им исследовательским рефлексом сперва тормозит весь запаздывающий рефлекс, затем, когда исследовательский рефлекс при повторении значительно слабеет, он только растормаживает первую фазу запаздывающего рефлекса, и, наконец, как вы узнаете теперь, он еще раз тормозит этот рефлекс, но уже на другом основании, сам становясь первичным возбудителем тормозного процесса в коре полушарий. Слабый же внешний агент, как это оказалось в опыте Фурсиковусловливая с самого начала слабый и коротко продолжающийся исследовательский рефлекс, начинает при первом применении прямо с растормаживающего действия на запаздывающий рефлекс и кончает тем же вторым торможением.
За тождество процессов торможения и сна говорят также и их общие свойства. В прошлых лекциях мы даже слишком много видели фактов, бесспорно доказывающих движение тормозного процесса по массе больших полушарий, причем это движение оказывалось очень медленным, измерялось минутами, и даже многими, и, кроме того, очень варьировало в отношении скорости у разных животных и при разных условиях. Очевидно, что и сон - тоже движущийся процесс. Все мы знаем, как постепенно овладевают нами сонливость и сон и как часто они трудно и медленно отступают, рассеиваются. Имеются и научные сведения о постепенном исчезании при засыпании работы разных органов чувств и других более сложных умственных деятельностей. С другой стороны, общеизвестны очень большие разницы между разными людьми в отношении скорости засыпания и пробуждения, а также разницы этой скорости при разных условиях. То же самое наблюдалось нами у собак.
Да: Далее в нашей работе мы постоянно видели, как торможение, если оно сначала развивается трудно, от практики, в связи с повторением и применением разных случаев торможения, делается все легче и легче воспроизводящимся процессам. Точно так же условные раздражители, когда они при соответствующих условиях обусловливают сон, а также индифферентные раздражители вызывают сон быстрее и быстрее, чем чаще они повторяются.
Интересно особенно следующее. Как показано ранее, торможение индуцирует возбуждение. У некоторых собак, у которых, как сообщено это несколько раньше, при запаздывающем рефлексе условный раздражитель вместо торможения в начальной фазе раздражения заменяется сном, наступлению сна иногда предшествует на очень короткое время небольшое общее возбуждение животного. А еще рельефнее и постояннее это явление выступает при засыпании животного под влиянием продолжительного или повторяющегося индифферентного раздражения. Это часто наблюдалось в вышеупомянутых опытах Розенталя. Когда индифферентный раздражитель явно начинает делать щенка сонливым, то, перед тем как ему окончательно заснуть, он приходит на некоторое время в возбужденное состояние: начинает беспокойно двигаться, чесаться, лаять беспричинно в воздух. То же мне приходилось замечать и на засыпающих детях. Совершенно своеобразная и неожиданная картина. Можно с правом видеть в ней явление индукции. Так же могло быть понимаемо и изместное возбуждение при начале наркотизации.
Мне кажется, что совокупность приведенных фактов должна считаться достаточной для полного доказательства истинности нашего положения, что сон и внутреннее торможение - один и тот же процесс. Я лично сейчас не знаю, не вижу ни одного факта в нашей работе, который бы противоречил серьезно нашему заключению. Надо сожалеть, что у нас до сих пор нет хорошего графического способа для изображения сна. Только изредка мы пользовались для этой цели записыванием положения головы животного. Конечно, сообщение наших опытов, относящихся ко сну, сопровождаемое тем или другим изображением сна, очень бы прибавило в ваших глазах убедительности наших доводов.
Очевидно, с нашим заключением вполне гармонируют и все подробности нашего нормального обычного сна. Наша дневная работа, у одних очень однообразная, у других, наоборот, очень разнообразная, одинаково в конце должна обусловливать наступление сна. Продолжительное раздражение одних и те же пунктов коры ведет к очень глубокому торможению их, которое, естественно, сильно иррадиируя, захватывает полушария и спускается на ниже лежащие отделы головного мозга. С другой стороны, при разнообразной деятельности, хотя отдельные пункты коры не доходят до значительной степени тормозного состояния, зато их большое количество создает и без большой иррадиации распространенное тормозное состояние, также спускающееся ниже. Конечно, очень большое количество быстро сменяющихся раздражений может часто долго сопротивляться общему захватыванию полушарий тормозным процессом и отсрочивать наступление сна. И обратно: строго заведенный порядок в смене бодрствования и сна, установленный ритм, может увеличивать настойчивость сна и без достаточного утомления клеток коры. Для того и другого мы имели в наших опытах достаточно примеров аналогичных отношений раздражительного и тормозного процессов.
При этом случае считаю не бесполезным ради ясности нарочито обратить ваше внимание на следующее. Очевидно, этот опыт вместе с вышеприведенными опытами Чечулина прибавляет лишнюю новую фазу в действии посторонних агентов на условный рефлекс. Если взять сильный внешний агент, то он, как вы помните из шестой лекции запаздывающем рефлексе, вызываемым им исследовательским рефлексом сперва тормозит весь запаздывающий рефлекс, затем, когда исследовательский рефлекс при повторении значительно слабеет, он только растормаживает первую фазу запаздывающего рефлекса, и, наконец, как вы узнаете теперь, он еще раз тормозит этот рефлекс, но уже на другом основании, сам становясь первичным возбудителем тормозного процесса в коре полушарий. Слабый же внешний агент, как это оказалось в опыте Фурсиковусловливая с самого начала слабый и коротко продолжающийся исследовательский рефлекс, начинает при первом применении прямо с растормаживающего действия на запаздывающий рефлекс и кончает тем же вторым торможением.
За тождество процессов торможения и сна говорят также и их общие свойства. В прошлых лекциях мы даже слишком много видели фактов, бесспорно доказывающих движение тормозного процесса по массе больших полушарий, причем это движение оказывалось очень медленным, измерялось минутами, и даже многими, и, кроме того, очень варьировало в отношении скорости у разных животных и при разных условиях. Очевидно, что и сон - тоже движущийся процесс. Все мы знаем, как постепенно овладевают нами сонливость и сон и как часто они трудно и медленно отступают, рассеиваются. Имеются и научные сведения о постепенном исчезании при засыпании работы разных органов чувств и других более сложных умственных деятельностей. С другой стороны, общеизвестны очень большие разницы между разными людьми в отношении скорости засыпания и пробуждения, а также разницы этой скорости при разных условиях. То же самое наблюдалось нами у собак.
Да: Далее в нашей работе мы постоянно видели, как торможение, если оно сначала развивается трудно, от практики, в связи с повторением и применением разных случаев торможения, делается все легче и легче воспроизводящимся процессам. Точно так же условные раздражители, когда они при соответствующих условиях обусловливают сон, а также индифферентные раздражители вызывают сон быстрее и быстрее, чем чаще они повторяются.
Интересно особенно следующее. Как показано ранее, торможение индуцирует возбуждение. У некоторых собак, у которых, как сообщено это несколько раньше, при запаздывающем рефлексе условный раздражитель вместо торможения в начальной фазе раздражения заменяется сном, наступлению сна иногда предшествует на очень короткое время небольшое общее возбуждение животного. А еще рельефнее и постояннее это явление выступает при засыпании животного под влиянием продолжительного или повторяющегося индифферентного раздражения. Это часто наблюдалось в вышеупомянутых опытах Розенталя. Когда индифферентный раздражитель явно начинает делать щенка сонливым, то, перед тем как ему окончательно заснуть, он приходит на некоторое время в возбужденное состояние: начинает беспокойно двигаться, чесаться, лаять беспричинно в воздух. То же мне приходилось замечать и на засыпающих детях. Совершенно своеобразная и неожиданная картина. Можно с правом видеть в ней явление индукции. Так же могло быть понимаемо и изместное возбуждение при начале наркотизации.
Мне кажется, что совокупность приведенных фактов должна считаться достаточной для полного доказательства истинности нашего положения, что сон и внутреннее торможение - один и тот же процесс. Я лично сейчас не знаю, не вижу ни одного факта в нашей работе, который бы противоречил серьезно нашему заключению. Надо сожалеть, что у нас до сих пор нет хорошего графического способа для изображения сна. Только изредка мы пользовались для этой цели записыванием положения головы животного. Конечно, сообщение наших опытов, относящихся ко сну, сопровождаемое тем или другим изображением сна, очень бы прибавило в ваших глазах убедительности наших доводов.
Очевидно, с нашим заключением вполне гармонируют и все подробности нашего нормального обычного сна. Наша дневная работа, у одних очень однообразная, у других, наоборот, очень разнообразная, одинаково в конце должна обусловливать наступление сна. Продолжительное раздражение одних и те же пунктов коры ведет к очень глубокому торможению их, которое, естественно, сильно иррадиируя, захватывает полушария и спускается на ниже лежащие отделы головного мозга. С другой стороны, при разнообразной деятельности, хотя отдельные пункты коры не доходят до значительной степени тормозного состояния, зато их большое количество создает и без большой иррадиации распространенное тормозное состояние, также спускающееся ниже. Конечно, очень большое количество быстро сменяющихся раздражений может часто долго сопротивляться общему захватыванию полушарий тормозным процессом и отсрочивать наступление сна. И обратно: строго заведенный порядок в смене бодрствования и сна, установленный ритм, может увеличивать настойчивость сна и без достаточного утомления клеток коры. Для того и другого мы имели в наших опытах достаточно примеров аналогичных отношений раздражительного и тормозного процессов.
Лекция шестнадцатая. Переходные фазы между бодрым состоянием и полным сном животного (гипнотические фазы)
Мм. гг.! Прошлый год я привел внушительное количество фактов, доказывающих, что сон есть внутреннее торможение, сплошное (a не раздробленное, постоянно перемежающееся с процессом раздражения), распространившееся по всей массе полушарий и спустившиеся также на некоторые нижележащие отделы головного мозга. Надо было ожидать, что, так как распространение торможения происходит постепенно, будет иметься различная экстенсивность сна, постепенное захватывание им то больших, то меньших районов. Следовательно, должны быть разные переходные формы к полному сну. Это и есть на самом деле, и мы наблюдали и производили их. При наших опытах мы имели дело не только с обыкновенной формой сна, характеризующейся при отсутствии нормальной деятельности полушарий расслаблением скелетной мускулатуры (закрытые глаза, низко спущенная голова, полусогнутые конечности и пассивно повисшес туловище на лямках, петлях для ног), но и с совершенно другой формой, что касается состояния скелетной мускулатуры. При ней деятельность полушарий также отсутствует, все условные раздражители остаются без действия и на всякие посторонние раздражители, если они не достигают большой силы, тоже нет никакой реакции, но животное сохраняет вполне активную позу. Оно стоит с открытыми неподвижными глазами, с поднятой головой, на вытянутых конечностях, отнюдь не опираясь на петли для ног, стоит неподвижно минутами и часами. При изменении положения его конечностей оно удерживает то, которое им придано. Отдергивание лапы при прикосновении к подошвенной стороне ее принимает характер контрактуры. При подаче еды животное тоже не реагирует, оставаясь неподвижным, и еды не берет. Такая форма торможения встречается довольно редко, и мы пока не знаем, с какими специальными условиями в обстановке нашего экспериментирования или с какой особенностью нервной системы она связана. Наш сотрудник Н. А. Рожанский, тщательно наблюдавший у собак переход бодрого состояния в сонное, пришел к заключению, что описанное состояние существует всегда при этом, но обыкак очень летучее, кратковременное явление. Физиологическое понимание этого состояния, как мне кажется, не представляет особенных затруднений. Перед нами заторможенная деятельность только больших полушарий, но торможение не спустилось ниже на центры, заведующие уравновешиванием, установкой тела в пространстве (центры Магнуса и Клейна), т. е. каталептическое состояние. Следовательно, в этой форме разграничительная линия между заторможенным отделом мозга и свободным от него лежит прямо под большими полушариями. Но она может разделять и большие районы самих полушарий. С этой невой формой мы встречаемся чаще и даже можем производить ее нарочно. Мы наблюдали ее впервые при следующих условиях (опыты Л. Н. Воскресенского). Одна из собак, с которой ранее работали без всякой помехи со стороны сна, вследствие того, что она в станке и одна в экспериментальной комнате была часто оставляема часами без всяких воздействий, начала впадать в сонное состояние. Очевидно, однообразные раздражения обстановки, как это указано в прошлой лекции, обусловили, наконец, сильное сплошное торможение мозга. Тормозящее действие обстановки сделалось таким сильным, что одно введение собаки в экспериментальную комнату сейчас же, видимо, делало ее другой, а тем более постановка в станок. Нужно было животное нарочно всячески раздражать, чтобы оно не оказалось заснувшим при окончании приготовлений к опыту. Когда затем экспериментатор уходил за дверь, чтобы оттуда вести опыт, и сейчас же, не теряя минуты, начинал действовать условными пищевыми раздражителями, нормальный условный рефлекс был налицо: текла слюна, и собака тотчас принималась за подаваемую ей еду. Но если по выходе из комнаты пропускалось 4 минут, получался совершенно особенный результат: слюноотделительный эффект на условный раздражитель был, слюнная реакция при подаче еды увеличивалась, но животное еды не брало, и надо было ему вводить ее насильно. В это время расслабленной скелетной мускулатуры еще не было. Если по выходе из комнаты ждать 10 минут до применения условного раздражителя, то теперь уже на него не было никакого эффекта и животное оказывалось вполне спящим, с расслабленной мускулатурой и храпом. Какое другое может быть объяснение этого факта, кроме того, что при первоначальном распространении торможения одно время оно захватило только двигательную область коры, оставляя пока нетронутыми все остальные районы коры, откуда условные раздражения шли к органу (железе), не связанному с двигательной областью. И только несколько позже сплошное торможение оказалось на всей массе полушарий и спустилось на нижележащие отделы мозга, и вместе с этим наступил полный сон. В данном случае эта стадия развивающегося сонного состояния получилась под влиянием индифферентных, долго действовавших на большие полушария раздражений. Обычно мы имеем ее при действии как отрицательных условных раздражителей, многократно повторяемых в одном и том же опыте, так и положительных условных раздражителей, специально слабых, тоже при частом и сплошном их применении. Привожу два примера. Первый относится к собаке, упоминавшейся уже в ранних лекциях, у которой тон в 256 колебаний в секунду был условным пищевым раздражителем, а по 10 тонов вверх и вниз от него были отдифференцированы (опыты Бирмана).
 Другой пример беру из работы (Розенталя) над собакой, у которой сначала было много и постоянных условных пищевых рефлексов. Но когда у нее был выработан еще один рефлекс на появление перед ее глазами экрана из серой бумаги и он повторялся много раз и часто сплошь один за другим в одном и том же опыте, то развилось следующее состояние собаки: часто на условные раздражители слюнная реакция была, и значительная, но к подаваемой еде собака не прикасалась.
Вот такой пример.
Другой пример беру из работы (Розенталя) над собакой, у которой сначала было много и постоянных условных пищевых рефлексов. Но когда у нее был выработан еще один рефлекс на появление перед ее глазами экрана из серой бумаги и он повторялся много раз и часто сплошь один за другим в одном и том же опыте, то развилось следующее состояние собаки: часто на условные раздражители слюнная реакция была, и значительная, но к подаваемой еде собака не прикасалась.
Вот такой пример.
 При этом собака только малоподвижна, но еще нет никакого видимого сна. Без применения условных раздражителей и на том же станке собака с жадностью ест ту же еди.
Сюда же относится и следующее случайное наблюдение. Одна из собак, давно служившая для опытов с условными рефлексами, никогда не представлявшая при условное пищевом рефлексе разъединения секреторной и двигательной реакций и никогда не спавшая в станке, была впервые поставлена перед многолюдной аудиторией для демонстрирования некоторых опытов с этими рефлексами. Необычная и очень сложная обстановка, видимо, оказала сильное действие на животное: оно оцепенело и слегка дрожало. При пробе условного раздражителя секреторный эффект получился, как обыкновенно, но поданную еду собака не взяла, а спустя довольно короткое время тут же в аудитории и в станке заснула, и с полным расслаблением скелетной мускулатуры. На этот раз, очевидно, сильный, необыкновенный раздражитель произвел прямо торможение больших полушарий сперва частичное, только в районе двигательной области, а потом и полное с переходом на нижележащий отдел мозга. Этот случай надо считать совершенно тождественным c обычной формой опыта с так называемым гипнозом животных, когда стремительное обездвиживание животного и укладывание его на спину также ведет к торможению в различной степени его распространенности, то лишь к каталепсии и даже частичной (неподвижность туловища при движении глаз и всей головы), то к полному сну. И у нас в лаборатории раз, - когда попалось очень непокорное животное, сильно сопротивлявшееся его снаряжению к опыту, - стремительное ограничение руками, при причинении ему значительного механического раздражения, сейчас же повело к полному сну животного в станке.
Таким образом как частичный сон, так и полный производится как слабыми, долго продолжающимися общими раздражениями, так и короткими, но сильными, как тормозными условными раздражителями, так и положительными. К некоторым подробностям, сюда относящимся, я еще не раз возвращусь в следующих лекциях.
Но рядом с различной экстенсивностью разлитого торможения наш фактический материал познакомил нас с различными вариациями, различными этапами самого процесса торможения, с различной, может быть позволительно сказать, интенсивностью разлитого торможения, сна.
Но прежде чем обратиться к этому предмету, я должен предварительно коснуться пункта, имеющего существенное значение для многих наших опытов относительно вариации разлитого тормозного процесса. В восьмой лекции я ставил вопрос: на чем основано в одновременном комплексном раздражителе замаскирование раздражителя из одного анализатора раздражителем из другого анализатора, и высказал предположение, что, может быть, оно связано с различной силой употребляемых нами агентов, принадлежащих к разным анализаторам. Там проектированные опыты были исполнены в течение этих наших лекций и вполне подтвердили то предположение. Когда мы нарочито чрезвычайно изменили относительную интенсивность наших обычных раздражителей: звуковые очень ослабили, а другие изи оставили, как они обыкновенно употреблялись, или усилили, то в эффекте комплексных раздражителей теперь, наоборот, оказались менее участвующими звуковые агенты, чем другие, т. е. при отдельных пробах условное действие принадлежало гораздо более вторым, чем первым.
Вот наши опыты. У одной собаки одновременное комплексное раздражение состояло из нашего обыкновенного кожно-механического раздражения и из звукового, чрезвычайно ослабленного (спыты В. В. Рикмана). Прочно выработанный раздражитель давал 4 4.5 капли слюны за 30 секунд изолированного действия. Отдельно испытанный звуковой компонент давал от 0.5 до 1.5 капли, а кожно-механический - от 2.5 до 5 капель. У другой собаки комплексное, также одновременное, раздражение составляли ритмическое вспыхивание лампочки в 400 свечей и звучание чрезвычайно заглушенного тона. Выработанный комплексный раздражитель давал 7-8 капель слюны за 30 секунд. Отдельно примененный световой раздражитель давал 5 капель, звуковой - 2.5. Точно так же комбинированное раздражение температурой 0°, приложенной к коже, и очень слабым звуком при разложении на компоненты показало гораздо большее действие на стороне холода сравнительно с звуком (опыты Гентта (W. Horsley Gantt) и Купалова).
Таким образом различие в величине эффекта наших обычных условных раздражителей, принадлежащих к различным анализаторам, обусловлено различной силой этих раздражителей, а не связано с качеством клеток разных анализаторов.
С этим положением мы можем приступить к нашей очередной теме об этапах разлитого тормозного процесса. Поводом к этому исследованию послужил случай патологического состояния нервной системы собаки под влиянием одного нашего приема не оперативного, a функционального характера. Экспериментально нами производимые патологическизменения нервной системы сами по себе займут наше внимание в следующей лекции. Здесь я опишу только исходный патологический опыт, из которого вытекло наше дальнейшее исследование на здоровых животных.
У собаки (опыты Разенкова) были выработаны положительные условные пищевые рефлексы на свисток, удары метронома, ритмическое механическое раздражение кожи (24 прикосновения в минуту), вспыхивание электрической лампочки, a также несколько дифференцировок, между ними дифференцировка на другую частоту прикосновений (12 в минуту) при механическом раздражении кожи на одном и том же месте.
Вот нормальное действие положительных условных раздражителей.
При этом собака только малоподвижна, но еще нет никакого видимого сна. Без применения условных раздражителей и на том же станке собака с жадностью ест ту же еди.
Сюда же относится и следующее случайное наблюдение. Одна из собак, давно служившая для опытов с условными рефлексами, никогда не представлявшая при условное пищевом рефлексе разъединения секреторной и двигательной реакций и никогда не спавшая в станке, была впервые поставлена перед многолюдной аудиторией для демонстрирования некоторых опытов с этими рефлексами. Необычная и очень сложная обстановка, видимо, оказала сильное действие на животное: оно оцепенело и слегка дрожало. При пробе условного раздражителя секреторный эффект получился, как обыкновенно, но поданную еду собака не взяла, а спустя довольно короткое время тут же в аудитории и в станке заснула, и с полным расслаблением скелетной мускулатуры. На этот раз, очевидно, сильный, необыкновенный раздражитель произвел прямо торможение больших полушарий сперва частичное, только в районе двигательной области, а потом и полное с переходом на нижележащий отдел мозга. Этот случай надо считать совершенно тождественным c обычной формой опыта с так называемым гипнозом животных, когда стремительное обездвиживание животного и укладывание его на спину также ведет к торможению в различной степени его распространенности, то лишь к каталепсии и даже частичной (неподвижность туловища при движении глаз и всей головы), то к полному сну. И у нас в лаборатории раз, - когда попалось очень непокорное животное, сильно сопротивлявшееся его снаряжению к опыту, - стремительное ограничение руками, при причинении ему значительного механического раздражения, сейчас же повело к полному сну животного в станке.
Таким образом как частичный сон, так и полный производится как слабыми, долго продолжающимися общими раздражениями, так и короткими, но сильными, как тормозными условными раздражителями, так и положительными. К некоторым подробностям, сюда относящимся, я еще не раз возвращусь в следующих лекциях.
Но рядом с различной экстенсивностью разлитого торможения наш фактический материал познакомил нас с различными вариациями, различными этапами самого процесса торможения, с различной, может быть позволительно сказать, интенсивностью разлитого торможения, сна.
Но прежде чем обратиться к этому предмету, я должен предварительно коснуться пункта, имеющего существенное значение для многих наших опытов относительно вариации разлитого тормозного процесса. В восьмой лекции я ставил вопрос: на чем основано в одновременном комплексном раздражителе замаскирование раздражителя из одного анализатора раздражителем из другого анализатора, и высказал предположение, что, может быть, оно связано с различной силой употребляемых нами агентов, принадлежащих к разным анализаторам. Там проектированные опыты были исполнены в течение этих наших лекций и вполне подтвердили то предположение. Когда мы нарочито чрезвычайно изменили относительную интенсивность наших обычных раздражителей: звуковые очень ослабили, а другие изи оставили, как они обыкновенно употреблялись, или усилили, то в эффекте комплексных раздражителей теперь, наоборот, оказались менее участвующими звуковые агенты, чем другие, т. е. при отдельных пробах условное действие принадлежало гораздо более вторым, чем первым.
Вот наши опыты. У одной собаки одновременное комплексное раздражение состояло из нашего обыкновенного кожно-механического раздражения и из звукового, чрезвычайно ослабленного (спыты В. В. Рикмана). Прочно выработанный раздражитель давал 4 4.5 капли слюны за 30 секунд изолированного действия. Отдельно испытанный звуковой компонент давал от 0.5 до 1.5 капли, а кожно-механический - от 2.5 до 5 капель. У другой собаки комплексное, также одновременное, раздражение составляли ритмическое вспыхивание лампочки в 400 свечей и звучание чрезвычайно заглушенного тона. Выработанный комплексный раздражитель давал 7-8 капель слюны за 30 секунд. Отдельно примененный световой раздражитель давал 5 капель, звуковой - 2.5. Точно так же комбинированное раздражение температурой 0°, приложенной к коже, и очень слабым звуком при разложении на компоненты показало гораздо большее действие на стороне холода сравнительно с звуком (опыты Гентта (W. Horsley Gantt) и Купалова).
Таким образом различие в величине эффекта наших обычных условных раздражителей, принадлежащих к различным анализаторам, обусловлено различной силой этих раздражителей, а не связано с качеством клеток разных анализаторов.
С этим положением мы можем приступить к нашей очередной теме об этапах разлитого тормозного процесса. Поводом к этому исследованию послужил случай патологического состояния нервной системы собаки под влиянием одного нашего приема не оперативного, a функционального характера. Экспериментально нами производимые патологическизменения нервной системы сами по себе займут наше внимание в следующей лекции. Здесь я опишу только исходный патологический опыт, из которого вытекло наше дальнейшее исследование на здоровых животных.
У собаки (опыты Разенкова) были выработаны положительные условные пищевые рефлексы на свисток, удары метронома, ритмическое механическое раздражение кожи (24 прикосновения в минуту), вспыхивание электрической лампочки, a также несколько дифференцировок, между ними дифференцировка на другую частоту прикосновений (12 в минуту) при механическом раздражении кожи на одном и том же месте.
Вот нормальное действие положительных условных раздражителей.
 Следовательно, на основании предыдущего, раздражители по силе располагаются, идя от сильного к слабым, так: свисток, удары метронома, механическое раздражение кожи и вспыхивание лампочки.
Затем ставится такой опыт, что среди других раздражений применяется дифференцированное механическое раздражение (12 прикосновений в минуту) в течение 30 секунд, и оно непосредственно сменяется на положительное механическое раздражение (24 прикосновения в минуту) тоже в течение 30 секунд.
На другой день после этого опыта и в следующие 9 дней все условные рефлексы исчезают, только очень изредка проявляясь и в минимальном размере. За этим периодом наступает совершенно особенный. Вот он.
Следовательно, на основании предыдущего, раздражители по силе располагаются, идя от сильного к слабым, так: свисток, удары метронома, механическое раздражение кожи и вспыхивание лампочки.
Затем ставится такой опыт, что среди других раздражений применяется дифференцированное механическое раздражение (12 прикосновений в минуту) в течение 30 секунд, и оно непосредственно сменяется на положительное механическое раздражение (24 прикосновения в минуту) тоже в течение 30 секунд.
На другой день после этого опыта и в следующие 9 дней все условные рефлексы исчезают, только очень изредка проявляясь и в минимальном размере. За этим периодом наступает совершенно особенный. Вот он.
 Получилось, как видите, совершенно обратное тому, что имелось раньше в норме: сильные раздражители совсем не действуют или едва, а слабые производят эффект, даже больший сравнительно с нормой. Это состояние полушарий, по примеру Н. Е. Введенского, мы назвали парадоксальной фазой. Парадоксальная фаза держалась 14 дней и перешла в следующую, такого вида:
Получилось, как видите, совершенно обратное тому, что имелось раньше в норме: сильные раздражители совсем не действуют или едва, а слабые производят эффект, даже больший сравнительно с нормой. Это состояние полушарий, по примеру Н. Е. Введенского, мы назвали парадоксальной фазой. Парадоксальная фаза держалась 14 дней и перешла в следующую, такого вида:
 Эту фазу мы назвали уравнительной фазой, так как при ней все раздражители оказывались равными по эффекту. Уровнительная фаза продолжалась 7 дней сменилась еще на новую фазу, где очень усилились средние раздражители, сильный несколько упал, а слабый совсем остался без эффекта. 7 дней спустя вернулась норма. В этой работе, как и в наших дальнейших опытах относительно этого предмета, применялись ради большей бесспорности одни и те же раздражители, но разной интенсивности, так что было очевидно, что все дело в отношении клеток именно к разной силе раздражения.
Таким образом мы в наших опытах впервые констатировали, что клетки полушарий переживают ряд переходных особенных состояний между нормально возбудимым и полным тормозным, обнаруживающихся в необычных отношениях этих клеток к различной силе раздражений.
Ознакомившись с этими переходными состояниями в явно патологическом случае, мы поставили себе дальше вопрос: не существуют ли эти состояния и в норме при переходе от бодрости ко сну и обратно? Представлялось вероятным, что в описанном случае патологическое состояние заключалось только в фиксировании надолго этих состоянии, когда в норме они могли быть скоро проходящими, не бросающимися в глаза, как это оказалось с каталепсией. Наше исследование направилось по этой дороге и привело к утвердительному ответу. Вот ряд наших опытов.
Вот собака, уже упоминавшаяся в этой лекции раньше, у которой, кроме одного положительно действующего тона, было 20 отдифференцированных соседних тонов. Среди многих других положительных условных рефлексов у нее были выработаны дополнительно рефлексы на тихий и громкий треск, резкоотличавшиеся по величине эффекта.
Вот их нормальные отношения.
Эту фазу мы назвали уравнительной фазой, так как при ней все раздражители оказывались равными по эффекту. Уровнительная фаза продолжалась 7 дней сменилась еще на новую фазу, где очень усилились средние раздражители, сильный несколько упал, а слабый совсем остался без эффекта. 7 дней спустя вернулась норма. В этой работе, как и в наших дальнейших опытах относительно этого предмета, применялись ради большей бесспорности одни и те же раздражители, но разной интенсивности, так что было очевидно, что все дело в отношении клеток именно к разной силе раздражения.
Таким образом мы в наших опытах впервые констатировали, что клетки полушарий переживают ряд переходных особенных состояний между нормально возбудимым и полным тормозным, обнаруживающихся в необычных отношениях этих клеток к различной силе раздражений.
Ознакомившись с этими переходными состояниями в явно патологическом случае, мы поставили себе дальше вопрос: не существуют ли эти состояния и в норме при переходе от бодрости ко сну и обратно? Представлялось вероятным, что в описанном случае патологическое состояние заключалось только в фиксировании надолго этих состоянии, когда в норме они могли быть скоро проходящими, не бросающимися в глаза, как это оказалось с каталепсией. Наше исследование направилось по этой дороге и привело к утвердительному ответу. Вот ряд наших опытов.
Вот собака, уже упоминавшаяся в этой лекции раньше, у которой, кроме одного положительно действующего тона, было 20 отдифференцированных соседних тонов. Среди многих других положительных условных рефлексов у нее были выработаны дополнительно рефлексы на тихий и громкий треск, резкоотличавшиеся по величине эффекта.
Вот их нормальные отношения.
 Опыт ведется дальше следующим образом. Повторным применением отдифференцированных тонов мы доводим собаку до явного засыпания и пускаем в действие слабый треск. Секреторного эффекта нет. Подачей еды мы пробуждаем собаку, она начинает есть. Через некоторое время слабый треск повторяется. Теперь он уже действует, но еще мало. Собака опять подкармливается. В третий раз слабый треск вызывает нормальный эффект и даже иногда несколько больший. Происходит опять подкрепление. В следующий раз применяется сильный треск. Его эффект более или менее уступает по величине эффекту перед этим действовавшего слабого треска. И только несколько позже при полном восстановлении бодрого состояния сильный треск достигает его полного нормального эффекта, и с этим вообще появляются обычные количественные отношения между этими двумя. раздражителями.
Вот подлинные цифры одного из таких опытов.
Опыт ведется дальше следующим образом. Повторным применением отдифференцированных тонов мы доводим собаку до явного засыпания и пускаем в действие слабый треск. Секреторного эффекта нет. Подачей еды мы пробуждаем собаку, она начинает есть. Через некоторое время слабый треск повторяется. Теперь он уже действует, но еще мало. Собака опять подкармливается. В третий раз слабый треск вызывает нормальный эффект и даже иногда несколько больший. Происходит опять подкрепление. В следующий раз применяется сильный треск. Его эффект более или менее уступает по величине эффекту перед этим действовавшего слабого треска. И только несколько позже при полном восстановлении бодрого состояния сильный треск достигает его полного нормального эффекта, и с этим вообще появляются обычные количественные отношения между этими двумя. раздражителями.
Вот подлинные цифры одного из таких опытов.
 Затем собака усыпляется применением отдифференцированпых тснов.
Затем собака усыпляется применением отдифференцированпых тснов.
 Иногда при таких опытах наблюдается при повторении этих раздражений, вместо первоначального преобладания в отношении эффекта слабого треска перед сильным, их равенство. Очевидно, при постепенном рассеивании сонного состояния под влиянием повторяющегося короткого акта еды корковая клетка, возвращаясь к бодрому состоянию, проходит и теперь как парадоксальную, так и уравнительную фазы.
Следовательно, то же самое, что патологическом случае, только то, что там держалось днями, здесь сменяется минутами (опыт Бирмана).
У другой нашей собаки, вследствие продолжительной работы с ней, развилось легкое дремотное состояние на станке, отразившееся на условных рефлексах в том, что ранее отчетливо отличавшиеся различные раздражители в отношении величины их условного эффекта теперь все сравнялись. Благодаря впрыскиваниям соответствующей дозы кофеина собака приходила в первоначальное бодрое состояние, а с ним восстановлялись и нормальные отношения между условными раздражителями (опыты Н. В. Зимкина).
Иногда при таких опытах наблюдается при повторении этих раздражений, вместо первоначального преобладания в отношении эффекта слабого треска перед сильным, их равенство. Очевидно, при постепенном рассеивании сонного состояния под влиянием повторяющегося короткого акта еды корковая клетка, возвращаясь к бодрому состоянию, проходит и теперь как парадоксальную, так и уравнительную фазы.
Следовательно, то же самое, что патологическом случае, только то, что там держалось днями, здесь сменяется минутами (опыт Бирмана).
У другой нашей собаки, вследствие продолжительной работы с ней, развилось легкое дремотное состояние на станке, отразившееся на условных рефлексах в том, что ранее отчетливо отличавшиеся различные раздражители в отношении величины их условного эффекта теперь все сравнялись. Благодаря впрыскиваниям соответствующей дозы кофеина собака приходила в первоначальное бодрое состояние, а с ним восстановлялись и нормальные отношения между условными раздражителями (опыты Н. В. Зимкина).
 На другой день за 18 минут до опыта собаке впрыскивается под кожу 8 куб. см 2% раствора Coffeini puri: собака совершенно бодра.
На другой день за 18 минут до опыта собаке впрыскивается под кожу 8 куб. см 2% раствора Coffeini puri: собака совершенно бодра.
 У собаки, которая уже упоминалась в этой лекции как представлявшая стадию разъединения секреторной реакции от двигательной, часто в этой стадии наблюдалось, что из всех раздражителей только самый слабый (вспыхивание лампочки) действовал иногда всего более положительно, вызывая полный нормальный рефлекс: текла слюна, и собака ела подаваемую ей еду. Таким образом в таких случаях с определенной степенью экстенсивности разлитого торможения существовала и парадоксальная фаза (опыты Розенталя).
При явном сонливом состоянии, но еще до наступления полного сна, замечалось еще следующее совершенно особенное явление. Когда условный положительный раздражитель терял совсем, или почти, свое действие, хорошо выработанный отрицательный раздражитель, наоборот, получал отчетливое положительное действие. Вот пример (опыты Шишло).
У собаки были выработаны пищевые положительные условные рефлексы на механическое раздражение кожи плеча и бедра и на приложение к коже тепла 45 50 С, а также постоянный отрицательный условный рефлекс на механическое раздражение кожи на спине. Эффект положительного механического раздражения кожи достигал в норме 15-18 капель в минуту. Температурный условный раздражитель довольно скоре начал вызывать сонливость и сон. В данном опыте сперва применяется температурный раздражитель. Развивается сонливость. Дальше опыт идет следующим образом.
У собаки, которая уже упоминалась в этой лекции как представлявшая стадию разъединения секреторной реакции от двигательной, часто в этой стадии наблюдалось, что из всех раздражителей только самый слабый (вспыхивание лампочки) действовал иногда всего более положительно, вызывая полный нормальный рефлекс: текла слюна, и собака ела подаваемую ей еду. Таким образом в таких случаях с определенной степенью экстенсивности разлитого торможения существовала и парадоксальная фаза (опыты Розенталя).
При явном сонливом состоянии, но еще до наступления полного сна, замечалось еще следующее совершенно особенное явление. Когда условный положительный раздражитель терял совсем, или почти, свое действие, хорошо выработанный отрицательный раздражитель, наоборот, получал отчетливое положительное действие. Вот пример (опыты Шишло).
У собаки были выработаны пищевые положительные условные рефлексы на механическое раздражение кожи плеча и бедра и на приложение к коже тепла 45 50 С, а также постоянный отрицательный условный рефлекс на механическое раздражение кожи на спине. Эффект положительного механического раздражения кожи достигал в норме 15-18 капель в минуту. Температурный условный раздражитель довольно скоре начал вызывать сонливость и сон. В данном опыте сперва применяется температурный раздражитель. Развивается сонливость. Дальше опыт идет следующим образом.
 То же мы не один раз наблюдали при некоторых патологических состояниях полушарии. Это состояние мы назвали ультрапарадоксальной фазой.
Наблюдая столько различных состояний клеток полушарий при переходе животного от бодрого состояния к полному сну и зная, что сон есть сплошное и разлитое внутреннее торможение, мы должны были ожидать, что некоторые из этих состояний мы встретим и в так называемом нами последовательном торможении, с которым мы основательно познакомились в ранних лекциях о внутреннем торможении. Как кажется, это и было, пока в единственном случае, который мы успели исследовать, именно при условном торможении (опыты Быкова).
У собаки имелись пять положительных условных раздражителей: удары метронома, громкий тон, он же сильно заглушенный, появление перед глазами собаки круга из картона и механическое раздражение кожи. Условное торможение было выработано на комбинацию механического раздражения кожи со звуком бульканья. Условные раздражители по величине слюноотделительного эффекта, средней из многих опытов, располагались в том порядке, как они перечислены: 22 капли, 18.5, 16.5, 13.5 и 10 капель за 30 секунд. Когда условное торможение только что было окончательно выработано, испытывались все условные раздражители. 10 минут спустя после того, как было применено условное торможение, удары метронома давали 16.5 капли, сильный тон 16, заглушенный тон 20 капель и круг 18 капель. Принимая во внимание возможное участие в факте как движения торможения, так и индукции, единственное, что можно было отнести на счет интересующего нас пункта, это то, что заглушенный тон действовал больше нормы, когда эффект сильного был менее нормы. Так как это происходило в одном и том же пункте коры, то в этом можно было видеть обнаружение парадоксальной фазы. Сейчас мы продолжаем это исследование дальше на других видах внутреннего торможения.
Затем мы остановились на следующем пункте. В лекции о взаимной индукции мы сделали предположение: не есть ли так называемое нами внешнее торможение явление отрицательной индукции, т. е. торможение, индуцированное на периферии района, находящегося в состоянии раздражения? Иначе говоря, не есть ли внешнее и внутреннее торможение в своей физико-химической основе один и тот же процесрассчитывали получить, может быть, некоторое подтверждение этого предположения при исследовании вопроса: не обусловливает ли внешнее торможение те же состояния корковой клетки, с которыми мы только что познакомились в случаях внутреннего торможения? Так как для этого исследования нужен был более или менее продолжительный период внешнего торможения, то мы производили его введением в рот животного различных отвергаемых веществ, при которых внешнее торможение дает себя знать очень долго.
Опыты произведены на двух собаках с пищевыми рефлексами.
У одной собаки (опыты Пророкова) после вливания ей в рот раствора соды, когда вызванное этим слюноотделение прекратилось, испытанные сейчас же как сильные, так и слабые условные раздражители были одинаково сильно заторможены, но позднее, в ближайшие 15--20 минут слабые уже действовали как в норме, так и даже больше и были или равны по действию с сильными, или даже в значительной степе-и превосходили их, если они были теперь еще очень ослаблены. Вот пример. в 9 часов 41 минуту влит раствор соды.
То же мы не один раз наблюдали при некоторых патологических состояниях полушарии. Это состояние мы назвали ультрапарадоксальной фазой.
Наблюдая столько различных состояний клеток полушарий при переходе животного от бодрого состояния к полному сну и зная, что сон есть сплошное и разлитое внутреннее торможение, мы должны были ожидать, что некоторые из этих состояний мы встретим и в так называемом нами последовательном торможении, с которым мы основательно познакомились в ранних лекциях о внутреннем торможении. Как кажется, это и было, пока в единственном случае, который мы успели исследовать, именно при условном торможении (опыты Быкова).
У собаки имелись пять положительных условных раздражителей: удары метронома, громкий тон, он же сильно заглушенный, появление перед глазами собаки круга из картона и механическое раздражение кожи. Условное торможение было выработано на комбинацию механического раздражения кожи со звуком бульканья. Условные раздражители по величине слюноотделительного эффекта, средней из многих опытов, располагались в том порядке, как они перечислены: 22 капли, 18.5, 16.5, 13.5 и 10 капель за 30 секунд. Когда условное торможение только что было окончательно выработано, испытывались все условные раздражители. 10 минут спустя после того, как было применено условное торможение, удары метронома давали 16.5 капли, сильный тон 16, заглушенный тон 20 капель и круг 18 капель. Принимая во внимание возможное участие в факте как движения торможения, так и индукции, единственное, что можно было отнести на счет интересующего нас пункта, это то, что заглушенный тон действовал больше нормы, когда эффект сильного был менее нормы. Так как это происходило в одном и том же пункте коры, то в этом можно было видеть обнаружение парадоксальной фазы. Сейчас мы продолжаем это исследование дальше на других видах внутреннего торможения.
Затем мы остановились на следующем пункте. В лекции о взаимной индукции мы сделали предположение: не есть ли так называемое нами внешнее торможение явление отрицательной индукции, т. е. торможение, индуцированное на периферии района, находящегося в состоянии раздражения? Иначе говоря, не есть ли внешнее и внутреннее торможение в своей физико-химической основе один и тот же процесрассчитывали получить, может быть, некоторое подтверждение этого предположения при исследовании вопроса: не обусловливает ли внешнее торможение те же состояния корковой клетки, с которыми мы только что познакомились в случаях внутреннего торможения? Так как для этого исследования нужен был более или менее продолжительный период внешнего торможения, то мы производили его введением в рот животного различных отвергаемых веществ, при которых внешнее торможение дает себя знать очень долго.
Опыты произведены на двух собаках с пищевыми рефлексами.
У одной собаки (опыты Пророкова) после вливания ей в рот раствора соды, когда вызванное этим слюноотделение прекратилось, испытанные сейчас же как сильные, так и слабые условные раздражители были одинаково сильно заторможены, но позднее, в ближайшие 15--20 минут слабые уже действовали как в норме, так и даже больше и были или равны по действию с сильными, или даже в значительной степе-и превосходили их, если они были теперь еще очень ослаблены. Вот пример. в 9 часов 41 минуту влит раствор соды.
 Обыкновенный эффект электрического звонка - около 8 капель за 30 секунд, эффект механического раздражения кожи около 4 капель.
У другой собаки (опыты П. К. Анохина) результат опытов частью согласовался результатом, полученным на первой собаке, частью представлялся особенным. После введения ответгаемых веществ в рот собаки и прекращения вызванного ими слюноотделения сейчас же и дальше до окончания опыта все раздражители, ранее постоянно различавшиеся между собой по величине эффекта (сильный звонок был первым раздражителем по эффекту, свет лампы - последним), сравнивались в этом отношении. Но рядом с этим происходило ступенеобразное падение рефлексов в течение опыта. Вот как шел опыт.
Влит раствор соды. Слюноотделение продолжалось 10 минут.
Обыкновенный эффект электрического звонка - около 8 капель за 30 секунд, эффект механического раздражения кожи около 4 капель.
У другой собаки (опыты П. К. Анохина) результат опытов частью согласовался результатом, полученным на первой собаке, частью представлялся особенным. После введения ответгаемых веществ в рот собаки и прекращения вызванного ими слюноотделения сейчас же и дальше до окончания опыта все раздражители, ранее постоянно различавшиеся между собой по величине эффекта (сильный звонок был первым раздражителем по эффекту, свет лампы - последним), сравнивались в этом отношении. Но рядом с этим происходило ступенеобразное падение рефлексов в течение опыта. Вот как шел опыт.
Влит раствор соды. Слюноотделение продолжалось 10 минут.
 Хотя полученный на обеих собаках результат в общем и благоприятен предположению, что внутреннее и внешнее торможение в своей основе является одним и тем же процессом, но сложность явлений требует повторения и варьирования этих опытов при более строгом внимании к другим возможным толкованиям фактов.
Наконец нам представлялось интересным исследовать: как относятся наши условные рефлексы при действии снотворных средств от начала этого действия до полного усыпления животного и обратно до возврата к бодрому состоянию. Мы употребляли с этой целью уретан и хлоралгидрат. Здесь почти исключительно преобладал другой ход явлений, именно постепенное падение всех рефлексов, так что слабые раздражители естественно ранее делались недействительными, чем сильные. Мы назвали такое состояние клеток наркотической фазой. Привожу один из этих опытов (С. И. Лебединской).
У собаки имелись следующие положительные условные раздражители: сильный звонок, удары метронома, тихий звонок, механическое раздражение кожи и прерывистое вспыхивание электрической лампочки перед глазами собаки. По эффекту раздражители располагались, как они перечислены. В 10 часов 9 минут введено в rectum 2.0 г хлоралгидрата в 150 куб. см воды. Собака стоит в станке. Опыт идет дальше так.
Хотя полученный на обеих собаках результат в общем и благоприятен предположению, что внутреннее и внешнее торможение в своей основе является одним и тем же процессом, но сложность явлений требует повторения и варьирования этих опытов при более строгом внимании к другим возможным толкованиям фактов.
Наконец нам представлялось интересным исследовать: как относятся наши условные рефлексы при действии снотворных средств от начала этого действия до полного усыпления животного и обратно до возврата к бодрому состоянию. Мы употребляли с этой целью уретан и хлоралгидрат. Здесь почти исключительно преобладал другой ход явлений, именно постепенное падение всех рефлексов, так что слабые раздражители естественно ранее делались недействительными, чем сильные. Мы назвали такое состояние клеток наркотической фазой. Привожу один из этих опытов (С. И. Лебединской).
У собаки имелись следующие положительные условные раздражители: сильный звонок, удары метронома, тихий звонок, механическое раздражение кожи и прерывистое вспыхивание электрической лампочки перед глазами собаки. По эффекту раздражители располагались, как они перечислены. В 10 часов 9 минут введено в rectum 2.0 г хлоралгидрата в 150 куб. см воды. Собака стоит в станке. Опыт идет дальше так.

 Мы видим, что при развитии снотворного действия все раздражители постепенно теряли в их эффекте, a при возврате к бодрому состоянию также все и постепенно достигали нормальной величины их эффекта. Единственное резкое исключение из 20 раздражений представил тихий звонок, произведший в 11 часов 53 минуты несоответственно большое действие.
Итак, на разных здоровых животных и при разных условиях мы получили много разных состояний полушарий в отношении реакции их на условные раздражители. Поднимался вопрос: насколько все эти состояния, включая сюда и наркотическую фазу, свойственны каждому животному и при обычных условиях жизни? При решении этого вопроса мы оказались в счастливом положении. Среди наших собак имелась одна (на ней были поставлены опыты, приведенные в конце четырнадцатой лекции), принадлежащая к резкому нервному типу, о котором будет сообцена в следующей лекции. Эта собака отличалась часто прямо изумительной при определенных условиях стереотипностью ее высшей нервной деятельности, в виде условных рефлексов. Заслуженно собака много раз получала отс название "живого инструмента". У нее, как было сообщено раньше, было 10 условных рефлексов: 6 положительных - на звонок, удары метронома, свисток, усиление общего освещения комнаты, появление перед ее глазами формы круга и игрушечной лошади, и 4 отрицательных - на другую частоту ударов метронома, уменьшение общего освещения комнаты, форму квадрата игрушечного зайчика приблизительно того же размера и цвета, как лошадка. В последнее время по некоторым основаниям нпотреблялся звонок, а из отрицательных мы пользовались почти только дифференцировкой ударов метронома. Эвукздражители обычно, а в ранних опытах постоянно, производили гораздо больший (на 30-50 %) слюноотделительный эффект, чем световые. После долгой лабораторной службы собаки (больше 2 лет) положительные условные раздражители стали обнаруживать тенденцию к уменьшению размера их действия и к изменению количественных отношений между ними, как это часто происходит с нашими собаками при долговременном пользовании одними и теми же условными раздражителями. И теперь-то мы могли видеть на нашей собаке в отчетливой форме все те состояния полушарий, которые описаны раньше в этой лекции как вариации наступающего разлитого тормозного процесса. Каждое такое состояние полушарий или резко выступало в продолжение целого опыта, или же то само собой, то под влиянием наших мер переходило в другие (опыты Сперанского). Единственно, что у нее не наблюдалось, - это ультрапарадоксальная фаза. Но не представлялось и случая для этого, так как собака никогда не становилась отчетливо сонливой.
Вот опыты из разных дней и разных периодов.
Мы видим, что при развитии снотворного действия все раздражители постепенно теряли в их эффекте, a при возврате к бодрому состоянию также все и постепенно достигали нормальной величины их эффекта. Единственное резкое исключение из 20 раздражений представил тихий звонок, произведший в 11 часов 53 минуты несоответственно большое действие.
Итак, на разных здоровых животных и при разных условиях мы получили много разных состояний полушарий в отношении реакции их на условные раздражители. Поднимался вопрос: насколько все эти состояния, включая сюда и наркотическую фазу, свойственны каждому животному и при обычных условиях жизни? При решении этого вопроса мы оказались в счастливом положении. Среди наших собак имелась одна (на ней были поставлены опыты, приведенные в конце четырнадцатой лекции), принадлежащая к резкому нервному типу, о котором будет сообцена в следующей лекции. Эта собака отличалась часто прямо изумительной при определенных условиях стереотипностью ее высшей нервной деятельности, в виде условных рефлексов. Заслуженно собака много раз получала отс название "живого инструмента". У нее, как было сообщено раньше, было 10 условных рефлексов: 6 положительных - на звонок, удары метронома, свисток, усиление общего освещения комнаты, появление перед ее глазами формы круга и игрушечной лошади, и 4 отрицательных - на другую частоту ударов метронома, уменьшение общего освещения комнаты, форму квадрата игрушечного зайчика приблизительно того же размера и цвета, как лошадка. В последнее время по некоторым основаниям нпотреблялся звонок, а из отрицательных мы пользовались почти только дифференцировкой ударов метронома. Эвукздражители обычно, а в ранних опытах постоянно, производили гораздо больший (на 30-50 %) слюноотделительный эффект, чем световые. После долгой лабораторной службы собаки (больше 2 лет) положительные условные раздражители стали обнаруживать тенденцию к уменьшению размера их действия и к изменению количественных отношений между ними, как это часто происходит с нашими собаками при долговременном пользовании одними и теми же условными раздражителями. И теперь-то мы могли видеть на нашей собаке в отчетливой форме все те состояния полушарий, которые описаны раньше в этой лекции как вариации наступающего разлитого тормозного процесса. Каждое такое состояние полушарий или резко выступало в продолжение целого опыта, или же то само собой, то под влиянием наших мер переходило в другие (опыты Сперанского). Единственно, что у нее не наблюдалось, - это ультрапарадоксальная фаза. Но не представлялось и случая для этого, так как собака никогда не становилась отчетливо сонливой.
Вот опыты из разных дней и разных периодов.


 Когда рефлексы очень падали и искажались, мы поднимали и исправляли их, как указано было в четырнадцатой лекции, тем, что действовали условными раздражителями до присоединения безусловного более короткое время. Отсюда в приведенных там опытах различная продолжительность изолированного условного раздражения. Переход из одной фазы в другую в двух последних опытах, надо думать, происходил под влиянием повторяющегося акта еды. Но мы сами располагали двумя специальными приемами, которыми сейчас же могли сменять фазы. Это - применение всегда полной дифференцировки (на другую частоту ударов метронома) как агента, концентрирующего торможение или индуцирующего процесс раздражения, а также применение социального раздражителя, которым являлось присутствие в экспериментальной комнате хозяина собаки - экспериментатора.
Вот примеры.
Когда рефлексы очень падали и искажались, мы поднимали и исправляли их, как указано было в четырнадцатой лекции, тем, что действовали условными раздражителями до присоединения безусловного более короткое время. Отсюда в приведенных там опытах различная продолжительность изолированного условного раздражения. Переход из одной фазы в другую в двух последних опытах, надо думать, происходил под влиянием повторяющегося акта еды. Но мы сами располагали двумя специальными приемами, которыми сейчас же могли сменять фазы. Это - применение всегда полной дифференцировки (на другую частоту ударов метронома) как агента, концентрирующего торможение или индуцирующего процесс раздражения, а также применение социального раздражителя, которым являлось присутствие в экспериментальной комнате хозяина собаки - экспериментатора.
Вот примеры.
 Полная тормозная фаза (нет ни секреторной, ни двигательной реакции) после применения дифференцировки переходит сперва в парадоксальную, а потом в нормальную фазу.
Полная тормозная фаза (нет ни секреторной, ни двигательной реакции) после применения дифференцировки переходит сперва в парадоксальную, а потом в нормальную фазу.
 Присутствие экспериментатора в комнате с собакой сейчас же превращает полную тормозную фазу в нормальную.
Вопрос: можно ли перечисленные переходные фазы в состоянии полушарий расположить в один ряд, и если можно, то в какой именно, - остается пока для нас полностью нерешенным. Если взять все наши случаи, преемственность фаз оказывается довольно разнообразной. Таким образом остается неясным: есть ли эти состояния строго последовательные или параллельные. Мы также не можем точно указать, почему данная фаза прямо переходит то в ту, то в другую. Следовательно, требуется дальнейшее исследование.
Едва ли можно сомневаться в том, чтописанные в этой лекции состояния полушарий есть то, что называется гипнозом в его разнообразных стадиях и чертах. Подробно явлениях человеческого гипноза в связи c полученными нами фактами будет речь в последней лекции.
Присутствие экспериментатора в комнате с собакой сейчас же превращает полную тормозную фазу в нормальную.
Вопрос: можно ли перечисленные переходные фазы в состоянии полушарий расположить в один ряд, и если можно, то в какой именно, - остается пока для нас полностью нерешенным. Если взять все наши случаи, преемственность фаз оказывается довольно разнообразной. Таким образом остается неясным: есть ли эти состояния строго последовательные или параллельные. Мы также не можем точно указать, почему данная фаза прямо переходит то в ту, то в другую. Следовательно, требуется дальнейшее исследование.
Едва ли можно сомневаться в том, чтописанные в этой лекции состояния полушарий есть то, что называется гипнозом в его разнообразных стадиях и чертах. Подробно явлениях человеческого гипноза в связи c полученными нами фактами будет речь в последней лекции.
Лекция семнадцатая разные типы нервной системы. Патологические состояния больших полушарий как результат функциональных воздействий на них
Мм. гг.! До сих пор мы занимались нормальной деятельпостью больших полушарий. Но опыты, которым мы подвергали наших животных, иначе говоря, нервные задачи, которые мы им ставили, не имея, конечно, сначала никакого представления о возможных пределах, так сказать, выносливости их мозга, производили у них иногда хронические нарушения нормальной деятельности полушарий. Я разумею здесь исключительно функциональные нарушения и производимые также только функционально, а не оперативно. Эти нарушения в одних случаях изглаживались постепенно сами собой, под влиянием одного отдыха вследствие прекращения соответствующих опытов, в другие разы оказывались столь упорными, что с нашей стороны требовались специальные лечебные меры. Таким образом перед нашими глазами физиология больших полушарий переходила в патологию и терапию их. Патологические состояния полушарий у разных наших животных под влиянием одних и тех же вредных условий проявлялись очень развод. Одни животные заболевали серьезно и надолго, другие легко и кратковременно, а третьи почти без следа переносили те же самые воздействия. У одних собак отклонения от нормы происходили в одну сторону, у других - в другую. Так как это разнообразие, очевидно, было связано с разницей характеров, типов нервной системы у разных животных, то, прежде чем обратиться к патологическим состояниям полушарий, необходимостановиться на типах нервной системы наших собак. В настоящее время при нашем изучении больших полушарий уже намечаются до некоторой степени точные критерии, при помощи которых можно будет достигать строго научной характеристики нервной системы того или другого из наших животных. Тогда, между прочим, откроется важная возможность строго научного экспериментального исследования на животных вопроса о наследственной передаче разных сторон нервной деятельности. Сейчас, однако, я опишу эти типы так, как они представляются обыденному наблюдению. Особенно выделились своей определенностью, своей резкостью, два типа, надо думать, крайние. Один тип давно познакомил нас с собой. О нем я уже не раз упоминал в моих ранних статьях и докладах как о типе, относительно которого при использовании его для наших опытов произошло вначале недоразумение. Когда в нашей ранней работе, по незнанию дела, мы испытывали порядочные затруднения со стороны сонливости животных, при некоторых условных раздражителях и при некоторых наших приемах, мы думали избавиться от этого зла выбором для наших опытов особенно подвижных на свободе собак. Это - животные в высшей степени суетливые, все обнюхивающие, все рассматривающие, быстро реагирующие на малейшие звуки, при знакомстве с людыхи (а знакомятся они очень легко) надоедливые своей навязчивостью, которых ни окриками, ни легкими ударами не удается скоро усмирить. И именно эти-то животные, поставленные в станок, ограниченные в их движениях и особенно оставленные одни в экспериментальной комнате, несмотря на применение условных раздражителей с подкармливанием или с вливанием кислоты, очень быстро становятся сонливыми, и условные рефлексы у них очень уменьшаются или и совсем исчезают. Наши повторяющиеся агенты, когда из них еще не успели выработаться прочные условные раздражители, демонстративно сейчас же вызывают сонливость, если с начала опыта собака еще бодра. Некоторые из этих собак даже при опытах не в станке, а на свободе, если экспериментатор остается к ним безучастным, так сказать, не развлекает их, тоже быстро начинают закрывать глаза, покачиваются и, наконец, укладываются на полу. И это часто сейчас же, как только собака после условного раздражителя была подкормлена. Сначала мы должны были отказываться от работы над такими животными. Но затем постепенно приспособились к ним. Если на таких собаках сразу вырабатывать условные рефлексы на много разных агентов, не повторять одних и тех же раздражителей в течение опыта, не делать больших промежутков между отдельными раздражителями, иметь у них не только положительные, но и тормозные рефлексы, короче говоря, поддерживать для них большое и, так сказать, деловое разнообразие опыта, они становятся вполнеудовлетворительными экспериментальными объектами. Остается вопросом, который мы в настоящее время решаем: представляет ли собой этот тип сильную или слабую нервную систему! Мне предварительно, впредь до более научно обоснованное системы, казалось, что типы собак, как я имел случай с ними ознакомиться при опытах в лаборатории, в некоторой степени отвечают древней классификации так называемых темпераментов. Только что описанные собаки должны быть признаны тогда за подлинных сангвиников. При быстрой смене раздражений они энергичны и деловиты, при малейшем однообразии обстановки они вялы, сонливы и, следовательно, бездеятельны. Другой тип собак, тоже очень ярко очерченный, должен быть поставлен на противоположном конце классической серии темпераментов. Во всякой новой, а тем более в несколько особенной обстановке они в высшей степени сдержанны в своих движениях, постоянно тормозят их: тихо идут около стены на не вполне разгибающихся ногах, часто при малейшем постороннем движении или звуке совсем прилегают всем туловищем к полу. Окрик или угрожающее движение человека моментально делает их неподвижными в пассивной распластанной позе. Всякий, увидавший их, сейчас же признает их за очень трусливых животных. Естественно, что они очень медленно осваиваются с обстановкой наших опытов, с разными манипуляциями на них. Но когда все это делается, наконец, для них обычным, они становятся для нашего исследования образцовыми объектахи. К такому типу принадлежит хваленое в конце прошлой лекции животное. Собаки этого типа в станке обыкновенно не спят. Если обстановка опытов остается более или менее однообразной, все условные рефлексы у них, а в особенности тормозные, оказываются в высшей степени стойкими и регулярными. Мы имеем сейчас в лаборатории и исследуем крайнего представителя этого типа. Эта собака-самка родилась у нас, никогда и ни от кого не получала никаких неприятностей. Когда ей было около года, ее начали приводить в лабораторию, где в станки в течение опыта только несколько раз подкармливали при образовании условных рефлексов. Какой она оказалась в лаборатории при первом ее появлении, такой она остается и теперь, 5 лет спустя. До сих пор она нисколько не привыкает к лаборатории. Проходя в сопровождении своего хозяина-экспериментатора в опытную комнату, как всегда с поджатым хвостом и на согнутых ногах, при встрече с постоянными членами лаборатории (некоторые из которых постоянно ее ласкают) она непременно или стремительно бросается в стороны, или пятится назад и приседает к полу. Она так же реагирует на малейшее ненарочитое более живое движение и чуть более громко сказанное слово своего хозяина. Она держится в отношении всех нас так, как если бы мы были ее опаснейшими врагами, от которых ей приходится постоянно и жестоко страдать. Однако, несмотря на это, когда она в опытной комнате, наконец, освоилась с обстановкой, у нее было образовано много точных как положительных, так и отрицательных рефлексов. Это было так для нас неожиданно, что она получила от нас лестное название «умницы». К этому животному я вернусь еще позднее. Не было бы преувеличенным отнести таких животных к типу меланхоликов. Как не считать их жизнь омраченной, если они постоянно и без надобности тормозят главное проявление жизни - движение! Оба описанные типа, очевидно, крайние типы: у одного чрезмерно преобладает раздражительный процесс, у другого - тормозной. Поэтому они и ограниченные типы, так сказать, с суженными пределами жизни. Одному нужна постоянная смена раздражений, их новизна, которой часто может не быть в окружающей действительности. Другому, наоборот, требуется очень однообразная обстановка жизни, в которой, однако же, может не быть тех или других колебаний и изменений. По поводу описанных типов считаю нужным коротенько остановиться на следующем. Может быть, кто-нибудь, ввиду сообщенного об этих типах, усмотрит некоторое возражение против положения о тождестве сна и внутреннего торможения в том, что тип с преобладанием раздражительного процесса очень наклонен ко сну в обстановке наших опытов, а тип, так легко тормозимый, наоборот, остается бодрым при том же условии. Но это, как уже указано, крайние типы, c особеачествами нервной системы и отсюда -- с особенными условиями своего существования. Если принимать - и с правом, -- что функциональная разрешаемость клеток дает толчок к возникновению в них тормозного процесса, то становится понятным, что чрезвычайно раздражимые корковые клетки, т. клетки с стремительным функциональным разрушением, и будут особенно наклонны к развитию в них тормозного процесса и к его широкому распространению, коль скоро они подвергаются продолжительным однообразным раздражениям. Лишь быстрая смена новых раздражений, адресующихся к другим клеткам, может нейтрализовать естественный, но, может быть, биологически невыгодный результат данного свойства нервных клеток. Точно так же второй тип, если бы его так легко наступающее торможение двигательной области коры, этот пассивно-оборонительный рефлекс, сопровождалось распространением торможения на всю массу полушарий и далее вниз по мозгу, был бы биологически невозможным, без всяких шансов прочного существования. Должна была оставаться возможность восстановления активной деятельности организма, коль скоро миновала причина, вызвавшая первоначальное торможение, а это и дают клетки полушарий, остающиеся незаторможенными. Следовательно, ограничение распространения торможения в данном случае есть специальное, биологически выработанное свойство в общем дефектной нервной системы, специальное приспособление, точно так, как человек может приучить себя спать даже на ходу, т. е. ограничивать торможение только полушариями, не позволяя ему спускаться ниже. Обращаюсь к другим вариациям нервной системы. В середине между описанными крайними типами стоят многочисленные средние типы, в которых существует уже большее или меньшее равновесие между процессами раздражения и торможения. Но, однако, одни из них несколько приближаются к одному крайнему типу, другие к другому, оставаясь в целом более широко приспособленными к жизни, являясь, следовательно, жизненно сильными. Одни из этих животных подвижны, активны и большей частью агрессивны. Другие более покойны, солидны, сдержанны. Я помню из последней группы одну собаку прямо поразительного поведения. Я никогда не видал, чтобы она по приводе ее из собачника в лаборатории в ожидании опыта ложилась на пол, но при этом как будто безучастно относилась к тому, что около нее происходило, и не вступала ни с кем из нас, не исключая и своего экспериментатора, ни в дружеские, ни во враждебные отношения. На станке никакой сонливости и всегда точные как положительные, так и отрицательные условные рефлексы и особенно последние. Ей надо было во всяком случае приписать сильный тормозной процесс. Но, однако, она была способна и к сильному возбуждению. Раз мне удалось нарушить ее обычное спокойствие, произведя перед ней при помощи одной игрушечной трубы чрезвычайные звуки, да еще с звериной маской на лице. Теперь только она потеряла свою всегдашнюю сдержанность, начала громко лаять и рваться по направлению ко мне. Истинно флегматическая, но сильная натура. Другая группа, очевидно, принадлежит более к возбудимому типу, соответствуя, может быть, древней классификации холерическому темпераменту. У этих животных отрицательные условные рефлексы нередко нарушаются. Конечно, есть немало животных менее определенного типа. Но в целом вся масса наших животных перед нашими глазами распадается на две категории: с чрезвычайным или умеренным преобладанием раздражительного процесса и чрезвычайным или умеренным преобладанием тормозного процесса. Теперь я могу перейти к патологическим состояниям полушарий, которые мы или наблюдали случайно, или производили сами нарочито. Первый случай этого рода был наблюдаем при следующих обстоятельствах. Как было указано еще в третьей лекции, мы могли выработать условный пищевой рефлекс на сильнейший электрический ток, приложенный к коже собаки. Вместо прирожденной оборонительной реакции животное отвечало на него пищевой реакцией: поворачивалось сторону, откуда подавалась еда, облизывалась и давало обильное количество слюны (опыты Ерофеевой). Выработка рефлекса начиналась с очень слабого тока, который потом постепенно усиливался и доводился до чрезвычайной силы. Такой рефлекс существовал много месяцев, причем электрический ток иногда заменялся прижиганием или механическим разрушением кожи с тем же результатом. Собака все время оставалась совершенно нормальной, и через несколько месяцев этот своеобразный условный раздражитель, как и все другие условные раздражители, начал медленно переходить в тормозной, именно начало слюноотделительного эффекта стало отодвигаться от момента раздражения. Мы и раньше кроме одного места, на котором первоначально вырабатывался этот рефлекс, изредка пробовали другие места, а затем решили систематически переходить с электродами на новые места. Некоторое время при этом пищевой рефлекс сохранялся попрежнему без малейшего вмешательства оборонительнойии. Но при известном дальнейшем месте все сразу и резко изменилось. От пищевой реакции не осталось и следа, была налицо только сильнейшая оборонительная реакция. Даже при чрезвычайно ослабленном токе, который до выработки условного рефлекса был недействительным, теперь и на самом первое месте кожи выступала все та же сильнейшая оборонительная реакция. То же было повторено еще на двух собаках. У одной из них этот взрыв возбуждения произошел на девятом месте, у другой на тринадцатом месте его еще не было. Но когда электрическое раздражение в одном и том же опыте было повторено на многих этих местах, а не на одном, как делалось раньше, случилось то же самое. У всех этих собак сейчас же после этого ничего нельзя было сделать для возврата пищевого рефлекса на ток. Животные сделались в высшей степени возбудимыми и беспокойными, какими они до этого никогда не бывали. У одной собаки после трехмесячного перерыва опытов можно было снова начать вырабатывать тот же рефлекс, вести выработку осторожнее, чем в первый раз, и, наконец, вернуть его. У других не помог и перерыв. Очевидно, указанной процедурой нервная система была приведена в хроническое патологическое состояние. К сожалению, от того времени не осталось указании относительно типов нервной системы этих собак. Вероятно потому, что описанный факт произошел при исключительных условиях, он не возбудил в к себе особенно деятельного интереса. Но несколько времени спустя мы наблюдали то же самое уже при более обычных условиях экспериментирования. Вот это наблюдение. Изучалась анализаторная деятельность в отношении предела различения форм предметов глазом (опыт Шенгер-Крестовниковой). Перед собакой отбрасывалась на экране светлая форма круга, и это сопровождалось кормлением животного. Когда рефлекс образовался, было приступлено к отдифференцированию от круга эллипсиса того же освещения и той же величины площади с отношением полуосей 2 : 1, тe. за появлением круга следовало подкармливание, при появлении эллипсиса еда не давалась. Дифференцирование полное и постоянное получилось довольно скоро. Затем этапами (отношения полуосей 3 : 2, 4 : 3 и т. д.) начали приближать эллипсис к кругу, продолжая выработку дифференцировок на эти последовательные эллипсисы. Выработка с колебаниями (сначала все быстрее, а затем опять замедляясь) шла гладко до эллипсиса с отношением полуосей 9: 8. Теперь хотя различение и наступило довольно значительное, но не полное. Через 3 недели применения этой дифференцировки воложение дела не улучшилось, а резко ухудшилось: различение круга от этого эллипсиса совершенно исчезло. Вместе c этим резко изменилось поведение собаки. Прежде спокойная, она теперь в станке визжала, вертелась, срывала приборчики, к ней прикрепленные, или перегрызала резиновые трубки, идущие от них к экспериментатору, чего никогда не случалось до этого. При введении в экспериментальную комнату лаяла, тоже против ее обычая. При пробе более легких дифференцировок оказалось, что все они пострадали, даже и самая первая с отношением полуосей 2 : 1. Эту последнюю до прежней точности пришлось доводить гораздо медленнее (в два раза с лишним), чем это было при первой ее выработке. Во время второй выработки легкой дифференцировки животное постепенно делалось покойнее и, наконец, совершенно возвратилось к нормальному состоянию. Переход к более тонким дифференцировкам происходил даже скорее, чем в первый раз. При первом применении эллипсиса с отношением полуосей 9:8 получилось полное различение от круга, но со второго же раза не осталось и следа от этого различения, и собака опять пришла в резко возбужденное состояние со всеми прежними последствиями. На этом опыты на ней были окончены. Вот несколько подлинных опытов из приведенной работы.
 В первом опыте эллипсис с отноолуосей 4 : 3 дал нуль эффекта. Во втором опыте эллипсис с отношением 9:8 в первом периоде выработки на него дифференцировки имел своим эффектом только 1 каплю, а после применения его в течение 2 недель в третьем опыте сравнялся в положительном эффекте с кругом. После этого даже эллипсис с отношением 2 : 1 в четвертом опыте уже не был полностью отдифференцирован и только в пятом опыте при постоянном применении в течение полутора месяцев снова стал нулевым.
После этой работы приведенный факт сосредоточил на себе наше внимание, и было приступлено его нарочитой разработке. Было ясно, что встреча при известных условиях процесса раздражения и процесса торможения ведет к нарушению обычного баланса между ними и создает в большей или меньшей мере на больший или меньший срок ненормальное состояние нервной системы. B первом приведенном случае при образовании из сильного электрического тока пищевого раздражителя должен был быть заторможенным прирожденный оборонительный рефлекс, и во втором случае при дифференцировании, как мы уже знаем из седьмой лекции, тоже должно было действовать торможение. До определенной степени правильное уравновешивание при этом противоположных процессов осуществлялось, но при известных условиях их относительной интенсивности или пространственного разграничения нормальный баланс между ними делался невозможным и все кончалось преобладанием одного из них (как увидим дальше), т. е. патологическим состоянием.
Для дальнейших опытов мы умышленно выбирали собак с разными типами нервной системы, чтобы видеть, как на каждой из них отражается патологическое состояние их нервной деятельности под влиянием наших воздействий функционального (не оперативного) характера. Первые такие опыты (Петровой) были исполнены на двух собаках с противоположными свойствами нервной системы, судя по их общему поведению. Это те же собаки, которые уже упоминались в лекции о сне и у которых сначала вызванное сонное состояние затем было устранено применением очень быстро одного за другим многих (6) условных рефлексов и притом при пятисекундном промежутке между началом условного раздражителя и присоединением к нему безусловного. Во время этих опытов, помимо устранения сна, получилось яркое и точное доказательство большой разницы в характерах нервной системы обеих собак, вполне подтвердившее первоначальный диагноз этих характеров на основании обыденного наблюдения. Это доказательство доставил нам процесс превращения у собак их почти совпадающих условных рефлексов в сильно запаздывающие, когда безусловный раздражитель присоединялся к условному лишь 3 минуты спустя после начала последнего. Превращение происходило постепенно: каждый день безусловный раздражитель отодвигался от начала условного на 5 секунд. Конечно, соответственно этому и так называемый нами латентный период, т. е. промежуток времени, через который от начала условного раздражителя наступает слюноотделение, постепенно удлинялся. Развитие этого запаздывания производилось сразу на всех раздражителях. В то время как одна собака, с преобладанием тормозного процесса, по нашему предварительному диагнозу, справилась с задачей запаздывания сразу и более или менее спокойно, другая, с преобладанием процесса раздражения, отнеслась к той же задаче совершенно иначе. Когда отставление достигло 2 минут, собака начала приходить в возбуждение, а при дальнейшем отставлении до предельной величины, 3 минут, прямо впала в неистовство: беспрерывно двигалась всеми частями тела, нестерпимо визжала и лаяла, слюноотделение сделалось сплошным, чрезвычайно увеличиваясь во время действия условных раздражителей; от запаздывания не осталось и следа. Очевидно, требуемое условиями опыта предварительное торможение многих условных раздражителей было не под силу, была слишком большой задачей для раздражимой нервной система этой собаки и выразилось в естественной борьбе против этого истязания. Надо прибавить, что эта трудность уравновешивания двух противоположных процессов наблюдалась нами у многих собак в форме возбуждения, но никогда еще это возбуждение не доходило до такой степени. Но, правда, и задача на этот раз была гораздо серьезнее, так как требовалось сбалансировать один процесс с другим сразу на многих пунктах полушарий. Нам ничего не оставалось, как прекратить опыт в таком виде. Очень интересно, что задача, оказавшаяся сперва невозможной, все же была затем удовлетворительно решена данной нервной системой. Для этого пришлось сперва ограничиться действием только одного условного раздражителя. Собака успокоилась и даже стала спать во время опыта не только в станке, но и на полу на свободе, во время опытов. Теперь снова были применены все условные раздражители, но только с пятисекундным действием их до присоединения безусловного раздражителя. Затем это изолированное действие опять постепенно было удлинено до 3 минут. На этот раз спокойно образовался хороший запаздывающий рефлекс в форме, о которой я упоминал в лекции о сне. 1.5-2 минуты с начала действия условного раздражителя собака была сонной, но в конце второй минуты или в самом начале третьей она быстро выходила из сонно-пассивного положения, и наступала резкая пищевая как двигательная, так и секреторная реакция. Таким образом, благодаря отдыху, постепенности и повторению, было достигнуто то уравновешивание обоих процессов, которое не могло осуществиться в первый раз.
Так как разница в нервной системе собак точно определилась, то мы обратились к главной цели опытов, но решили идти к ней несколько иначе, чем это было в вышеописанных случайных наблюдениях. На сильно запаздывающих рефлексах, т. е. с длинным предварительным тормозным процессом, мы применили другие виды торможения: дифференцировочное, условное и угасательное. Расчет был такой: может быть, при такой сложной системе торможений произойдет то же нарушение нормального равновесия между обоими нервными процессами, которое было в тех наблюдениях. Наш расчет не оправдался, нарушения не наступило. Но при всех этих прибавочных случаях торможения постоянно подчеркивалась разница между обеими нашими собаками. Раздражимая собака выработку каждого нового торможения сопровождала временно значительным возбуждением, другая же при этом не обнаруживала почти никаких знаков затруднения. При таком положении дела мы воспользовались испытанным средством. Было приступлено к выработке условного пищевого рефлекса на электрический ток, приложенный к коже. Рефлекс выработался и практиковался с некоторыми перерывами в течение значительного времени. Теперь и без переноса электродов все на новые места, как это было в опытах Ерофеевой, наступило хроническое изменение нервной системы у их собак. Надо думать, что здесь имело влияние указанное предварительное усложнение тормозной деятельности. Но что было важно и ново, это то, что нарушение нормальной работы нервной системы y обеих собак выразилось прямо противоположно: у одной хронически пострадали отрицательные, тормозные рефлексы, у другой положительные и, лишь позже, последовательно тормозные.
Вот подробный ход опытов.
У возбудимой собаки имелись условные раздражители на удары метронома, звонок, бульканье, механическое раздражение кожи на бедре и отрицательные - на шипение вместе с ударами метронома (шипение на 5 секунд предшествовало ударам метронома) и механическое раздражение кожи на плече. Все условные раздражители продолжались до подкармливания 3 минуты.
В первом опыте эллипсис с отноолуосей 4 : 3 дал нуль эффекта. Во втором опыте эллипсис с отношением 9:8 в первом периоде выработки на него дифференцировки имел своим эффектом только 1 каплю, а после применения его в течение 2 недель в третьем опыте сравнялся в положительном эффекте с кругом. После этого даже эллипсис с отношением 2 : 1 в четвертом опыте уже не был полностью отдифференцирован и только в пятом опыте при постоянном применении в течение полутора месяцев снова стал нулевым.
После этой работы приведенный факт сосредоточил на себе наше внимание, и было приступлено его нарочитой разработке. Было ясно, что встреча при известных условиях процесса раздражения и процесса торможения ведет к нарушению обычного баланса между ними и создает в большей или меньшей мере на больший или меньший срок ненормальное состояние нервной системы. B первом приведенном случае при образовании из сильного электрического тока пищевого раздражителя должен был быть заторможенным прирожденный оборонительный рефлекс, и во втором случае при дифференцировании, как мы уже знаем из седьмой лекции, тоже должно было действовать торможение. До определенной степени правильное уравновешивание при этом противоположных процессов осуществлялось, но при известных условиях их относительной интенсивности или пространственного разграничения нормальный баланс между ними делался невозможным и все кончалось преобладанием одного из них (как увидим дальше), т. е. патологическим состоянием.
Для дальнейших опытов мы умышленно выбирали собак с разными типами нервной системы, чтобы видеть, как на каждой из них отражается патологическое состояние их нервной деятельности под влиянием наших воздействий функционального (не оперативного) характера. Первые такие опыты (Петровой) были исполнены на двух собаках с противоположными свойствами нервной системы, судя по их общему поведению. Это те же собаки, которые уже упоминались в лекции о сне и у которых сначала вызванное сонное состояние затем было устранено применением очень быстро одного за другим многих (6) условных рефлексов и притом при пятисекундном промежутке между началом условного раздражителя и присоединением к нему безусловного. Во время этих опытов, помимо устранения сна, получилось яркое и точное доказательство большой разницы в характерах нервной системы обеих собак, вполне подтвердившее первоначальный диагноз этих характеров на основании обыденного наблюдения. Это доказательство доставил нам процесс превращения у собак их почти совпадающих условных рефлексов в сильно запаздывающие, когда безусловный раздражитель присоединялся к условному лишь 3 минуты спустя после начала последнего. Превращение происходило постепенно: каждый день безусловный раздражитель отодвигался от начала условного на 5 секунд. Конечно, соответственно этому и так называемый нами латентный период, т. е. промежуток времени, через который от начала условного раздражителя наступает слюноотделение, постепенно удлинялся. Развитие этого запаздывания производилось сразу на всех раздражителях. В то время как одна собака, с преобладанием тормозного процесса, по нашему предварительному диагнозу, справилась с задачей запаздывания сразу и более или менее спокойно, другая, с преобладанием процесса раздражения, отнеслась к той же задаче совершенно иначе. Когда отставление достигло 2 минут, собака начала приходить в возбуждение, а при дальнейшем отставлении до предельной величины, 3 минут, прямо впала в неистовство: беспрерывно двигалась всеми частями тела, нестерпимо визжала и лаяла, слюноотделение сделалось сплошным, чрезвычайно увеличиваясь во время действия условных раздражителей; от запаздывания не осталось и следа. Очевидно, требуемое условиями опыта предварительное торможение многих условных раздражителей было не под силу, была слишком большой задачей для раздражимой нервной система этой собаки и выразилось в естественной борьбе против этого истязания. Надо прибавить, что эта трудность уравновешивания двух противоположных процессов наблюдалась нами у многих собак в форме возбуждения, но никогда еще это возбуждение не доходило до такой степени. Но, правда, и задача на этот раз была гораздо серьезнее, так как требовалось сбалансировать один процесс с другим сразу на многих пунктах полушарий. Нам ничего не оставалось, как прекратить опыт в таком виде. Очень интересно, что задача, оказавшаяся сперва невозможной, все же была затем удовлетворительно решена данной нервной системой. Для этого пришлось сперва ограничиться действием только одного условного раздражителя. Собака успокоилась и даже стала спать во время опыта не только в станке, но и на полу на свободе, во время опытов. Теперь снова были применены все условные раздражители, но только с пятисекундным действием их до присоединения безусловного раздражителя. Затем это изолированное действие опять постепенно было удлинено до 3 минут. На этот раз спокойно образовался хороший запаздывающий рефлекс в форме, о которой я упоминал в лекции о сне. 1.5-2 минуты с начала действия условного раздражителя собака была сонной, но в конце второй минуты или в самом начале третьей она быстро выходила из сонно-пассивного положения, и наступала резкая пищевая как двигательная, так и секреторная реакция. Таким образом, благодаря отдыху, постепенности и повторению, было достигнуто то уравновешивание обоих процессов, которое не могло осуществиться в первый раз.
Так как разница в нервной системе собак точно определилась, то мы обратились к главной цели опытов, но решили идти к ней несколько иначе, чем это было в вышеописанных случайных наблюдениях. На сильно запаздывающих рефлексах, т. е. с длинным предварительным тормозным процессом, мы применили другие виды торможения: дифференцировочное, условное и угасательное. Расчет был такой: может быть, при такой сложной системе торможений произойдет то же нарушение нормального равновесия между обоими нервными процессами, которое было в тех наблюдениях. Наш расчет не оправдался, нарушения не наступило. Но при всех этих прибавочных случаях торможения постоянно подчеркивалась разница между обеими нашими собаками. Раздражимая собака выработку каждого нового торможения сопровождала временно значительным возбуждением, другая же при этом не обнаруживала почти никаких знаков затруднения. При таком положении дела мы воспользовались испытанным средством. Было приступлено к выработке условного пищевого рефлекса на электрический ток, приложенный к коже. Рефлекс выработался и практиковался с некоторыми перерывами в течение значительного времени. Теперь и без переноса электродов все на новые места, как это было в опытах Ерофеевой, наступило хроническое изменение нервной системы у их собак. Надо думать, что здесь имело влияние указанное предварительное усложнение тормозной деятельности. Но что было важно и ново, это то, что нарушение нормальной работы нервной системы y обеих собак выразилось прямо противоположно: у одной хронически пострадали отрицательные, тормозные рефлексы, у другой положительные и, лишь позже, последовательно тормозные.
Вот подробный ход опытов.
У возбудимой собаки имелись условные раздражители на удары метронома, звонок, бульканье, механическое раздражение кожи на бедре и отрицательные - на шипение вместе с ударами метронома (шипение на 5 секунд предшествовало ударам метронома) и механическое раздражение кожи на плече. Все условные раздражители продолжались до подкармливания 3 минуты.
 С конца марта началось образование условного рефлекса на электрический ток. В апреле он образовался. Пока применяемый ток был не очень большой силы, все виды торможения более или менее сохранялись. В августе ток был очень усилен. Теперь впервые нарушилось запаздывание и условное торможение перестало быть полным. Для облегчения положения все условные раздражители, кроме звонка, продолжались изолированно до подкармливания только 30 секунд вместо 3 минут. Тем не менее ослабление тормозного процесса, несмотря на прекращение применения тока, шло вперед. Запаздывание совершенно исчезло. Шипение, предшествовавшее на 5 секунд ударам метронома в тормозной комбинации, само получило постоянное положительное действие, т. е. сделалось условным раздражителем второго порядка, и даже дифференцировка на механическое раздражение в значительной степени растормозилась.
С конца марта началось образование условного рефлекса на электрический ток. В апреле он образовался. Пока применяемый ток был не очень большой силы, все виды торможения более или менее сохранялись. В августе ток был очень усилен. Теперь впервые нарушилось запаздывание и условное торможение перестало быть полным. Для облегчения положения все условные раздражители, кроме звонка, продолжались изолированно до подкармливания только 30 секунд вместо 3 минут. Тем не менее ослабление тормозного процесса, несмотря на прекращение применения тока, шло вперед. Запаздывание совершенно исчезло. Шипение, предшествовавшее на 5 секунд ударам метронома в тормозной комбинации, само получило постоянное положительное действие, т. е. сделалось условным раздражителем второго порядка, и даже дифференцировка на механическое раздражение в значительной степени растормозилась.

 У спокойной собаки имелись те же условные раздражители, что и у возбудимой.
У спокойной собаки имелись те же условные раздражители, что и у возбудимой.
 C конца марта приступлено к образованию условного рефлекса на электрический ток. Этот рефлекс легко и скоро достиг величины 7 капель в течение 30 секунд. При дальнейшем усилении тока оборонительная реакция возвращалась, но затем исчезала, уступая место вполне лищевой. Однако скоро, при повторении выработанного рефлекса на ток, слюноотделительный эффект рефлекса стал уменьшаться и испытанные в это время рефлексы на другие условные раздражители почти исчезли, появляясь только в начале опыта и в слабой степени. Вот опыт 30 V 1923, демонстрирующий это положение дела.
C конца марта приступлено к образованию условного рефлекса на электрический ток. Этот рефлекс легко и скоро достиг величины 7 капель в течение 30 секунд. При дальнейшем усилении тока оборонительная реакция возвращалась, но затем исчезала, уступая место вполне лищевой. Однако скоро, при повторении выработанного рефлекса на ток, слюноотделительный эффект рефлекса стал уменьшаться и испытанные в это время рефлексы на другие условные раздражители почти исчезли, появляясь только в начале опыта и в слабой степени. Вот опыт 30 V 1923, демонстрирующий это положение дела.
 Так как в это время собака стала худеть и сделалась вялой, то все опыты были прерваны на значительный срок и собаку стали усиленно кормить, между прочим давали рыбий жир. Собака вернула прежний вес и бодрость. После перерыва все обычные условные рефлексы, кроме рефлекса на звонок, переделали из отставленных на 3 минуты в отставленные на 30 секунд, но это существенно не изменило результата: рефлексы восстановились только в незначительной степени. Примененный в это время электрический ток дал значительный слюноотделительный эффект, но при усилении тока это его действие опять ослабело и, наконец, и совсем исчезло, как и действие всех остальных рефлексов. Больше того. На этот раз начали исчезать постепенно и все виды внутреннего торможения, т. е. отрицательные условные раздражители, прежде нулевые, стали сопровождаться иногда более или менее значительным слюноотделением.
Вот опыт 6 XII 1923, показывающий положительное действие условного раздражителя только в тормозную фазу запахдывающего рефлекса.
Так как в это время собака стала худеть и сделалась вялой, то все опыты были прерваны на значительный срок и собаку стали усиленно кормить, между прочим давали рыбий жир. Собака вернула прежний вес и бодрость. После перерыва все обычные условные рефлексы, кроме рефлекса на звонок, переделали из отставленных на 3 минуты в отставленные на 30 секунд, но это существенно не изменило результата: рефлексы восстановились только в незначительной степени. Примененный в это время электрический ток дал значительный слюноотделительный эффект, но при усилении тока это его действие опять ослабело и, наконец, и совсем исчезло, как и действие всех остальных рефлексов. Больше того. На этот раз начали исчезать постепенно и все виды внутреннего торможения, т. е. отрицательные условные раздражители, прежде нулевые, стали сопровождаться иногда более или менее значительным слюноотделением.
Вот опыт 6 XII 1923, показывающий положительное действие условного раздражителя только в тормозную фазу запахдывающего рефлекса.

 С основанием надо принимать, что положительное действие условных тормозных раздражителей не обозначало ослабление тормозного процесса, а свидетельствовало о дальнейшем поражении процесса раздражения, являясь ультрапарадоксальной фазой состояния прежде нормально раздражимых клеток.
В это время общее состояние собаки было вполне удовлетворительное.
Итак, у двух собак разного типа нервной системы под действием совершенно одних и тех же вредных условий наступало хроническое отклонение от нормы нервной деятельности, но в разных направлениях. У одной, раздражимой, чрезвычайно ослаб и почти исчез процесс торможения в корковых клетках полушарий, у другой, спокойной, обычно хорошо тормозимой, чрезвычайно ослаб и почти исчез процесс раздражения в тех же клетках. Иначе говоря, мы имели перед собой два различных невроза.
Произведенные нами неврозы оказались очень упорными и длительными. При перерыве опытов они не обнаружили наклонности к поправлению. Тогда у раздражимой собаки для изучения мы решили применить испытанное терапевтическое средство бром, тем более, что в наших ранних опытах (опыты П. М. Никиворовского и В. С. Дерябина) иногда мы видела благоприятное действие брома для усиления торможения в случае его недостаточности. После существования невроза течение нескольких месяцев собаке стали вводить в rectum ежедневно по 100 куб. см 2 % раствора Все виды внутреннего торможения начали 316 Лекции о работе больших полушарий головного мозга быстро восстанавливаться в определенном порядке: прежде всего сделалась полной и прочной дифференцировка, затем условное торможение и, наконец, запаздыесятому дню все рефлексы были вполне нормальны.
С основанием надо принимать, что положительное действие условных тормозных раздражителей не обозначало ослабление тормозного процесса, а свидетельствовало о дальнейшем поражении процесса раздражения, являясь ультрапарадоксальной фазой состояния прежде нормально раздражимых клеток.
В это время общее состояние собаки было вполне удовлетворительное.
Итак, у двух собак разного типа нервной системы под действием совершенно одних и тех же вредных условий наступало хроническое отклонение от нормы нервной деятельности, но в разных направлениях. У одной, раздражимой, чрезвычайно ослаб и почти исчез процесс торможения в корковых клетках полушарий, у другой, спокойной, обычно хорошо тормозимой, чрезвычайно ослаб и почти исчез процесс раздражения в тех же клетках. Иначе говоря, мы имели перед собой два различных невроза.
Произведенные нами неврозы оказались очень упорными и длительными. При перерыве опытов они не обнаружили наклонности к поправлению. Тогда у раздражимой собаки для изучения мы решили применить испытанное терапевтическое средство бром, тем более, что в наших ранних опытах (опыты П. М. Никиворовского и В. С. Дерябина) иногда мы видела благоприятное действие брома для усиления торможения в случае его недостаточности. После существования невроза течение нескольких месяцев собаке стали вводить в rectum ежедневно по 100 куб. см 2 % раствора Все виды внутреннего торможения начали 316 Лекции о работе больших полушарий головного мозга быстро восстанавливаться в определенном порядке: прежде всего сделалась полной и прочной дифференцировка, затем условное торможение и, наконец, запаздыесятому дню все рефлексы были вполне нормальны.
 Надо заметить, что при этом не наблюдалось никакого уменьшения положительных рефлексов. Главное, что теперь обращало на себя внимание, - это большое постоянство этих рефлексов. Таким образом, по нашим опытам, бром является средством, не понижающим нервную возбудимость, а ее регулирующим.
Бром вводился только 11 дней. Но излечение им невроза на этот раз было радикальным, так какефлексы и 21/2 месяца спустя оставались вполне нормальными.
Невроз спокойной собаки не поддался ни влиянию брома, ни другим мерам, которые мы предпринимали. Она была оставлена без опытов на очень большой срок, в течение которого мы за ней не следили. После этого перерыва, неожиданно для нас, она оказалась вполне нормальной. ней мы встретимся еще раз. в следующей лекции.
Надо заметить, что при этом не наблюдалось никакого уменьшения положительных рефлексов. Главное, что теперь обращало на себя внимание, - это большое постоянство этих рефлексов. Таким образом, по нашим опытам, бром является средством, не понижающим нервную возбудимость, а ее регулирующим.
Бром вводился только 11 дней. Но излечение им невроза на этот раз было радикальным, так какефлексы и 21/2 месяца спустя оставались вполне нормальными.
Невроз спокойной собаки не поддался ни влиянию брома, ни другим мерам, которые мы предпринимали. Она была оставлена без опытов на очень большой срок, в течение которого мы за ней не следили. После этого перерыва, неожиданно для нас, она оказалась вполне нормальной. ней мы встретимся еще раз. в следующей лекции.
Лекция восемнадцатая. Патологические состояния больших пполушарии как результат функциональных воздействий на них
Мм. гг.! В настоящей лекции я продолжаю описание нашел опытов и наблюдений над патологическими состояниями коры больших полушарий. Эта тема под нашими руками и перед нашими глазами все ширится и углубляется не только вследствие сосредоточения на ней теперь нашего нарочитого внимания, но и благодаря некоторым случайностям. При этом мы постоянно видим, как нормальное под вредными разрушительными влияниями незаметными переходами превращается в патологическое и как патологическое часто открывает нам, разлагая и упрощая, то, что заслонено от нас, слитое и усложненное, в физиологической норме. В лекции о нормальных гипнотических состояниях было сообщено, что особенно интересные из них (с точки зрения приложимости их к человеку, как это будет показано в последней лекции) сделались предметом нашего исследования лишь после того, как они ярко выступили перед нами в патологическом случае. Вы помните, что там у собаки, имевшей несколько положительных и отрицательных условных рефлексов, между ними были: положительный кожно-механический на 24 ритмических прикосновения к коже в течение 30 секунд и отрицательный на 12 прикосновений в тот же срок. Положительные рефлексы в норме представляли отчетливую градацию в величине слюноотделительного эффекта соответственно силе условных раздражителей. Когда в одном опыте после отрицательного кожно-механического раздражителя непосредственно, без малейшего промежутка времени, был применен положительный кожно-механический раздражитель, т. е. одна частота прикосновений была сменена на другую, то произошло патологическое состояние коры. Оно выразилось в ближайшие дни в исчезании всех положительных условных рефлексов, а затем в течение длинного ряда дней в различно измененных против нормы, но определенно сменявшихся отношениях появившегося слюноотделительного эффекта к силе условных раздражителей. Все это патологическое состояние протекло в 5/2 недель. Очевидно, что этот случай должен быть поставлен в ряд с теми, которые были описаны в предыдущей лекции. То, что там у последней собаки проявилось в виде сильнейшего симптома, исчезания всех положительных условных рефлексов, державшегося многие месяцы, здесь, переходя через приближающиеся к норме фазы, закончилось к тридцать шестому дню. Вместе с тем ясно, что основной механизм происхождения патологического состояния во всех доселе приведенных случаях - один и тот же. Это - трудная встреча, столкновение процесса раздражения и торможения. Помимо уже описанных случаев патологического состояния коры полушарий, мы имеем еще и дальнейшие, представляющие не меньший, если не больший интерес как в отношении некоторых особенностей патологического состояния, так и в отношении несколько иного механизма его происхождения. Я остановлюсь прежде всего на случае, особенно тщательно изо дня в день прослеженном в течение нескольких месяцев (опыт Рикмана). Ввиду исключительного интереса, им представляемого, я воспроизведу его особенно подробно и документально. Дело идет о собаке, очень тормозимой и служившей ранее для многих опытов и наблюдений. У нее имелся ряд положительных условных пищевых рефлексов и один отрицательный на 60 ударов метронома в минуту, когда 120 ударов в минуту было положительным раздражителем. Положительные условные рефлексы отчетливо располагались по величине слюноотделительного эффекта, соответственно силе примененных для условного действия внешних агентов. Отрицательный условный раздражитель, так как к началу настоящего опыта насчитывалось 266 его повторений, был вполне точным, постоянным и достаточно сконцентрированным, т. е. его последовательное тормозящее действие на положительные рефлексы ограничивалось коротким сроком времени. Вот пример нормального опыта 1 XII 1925. Ввиду того, что собаки этого типа являются как бы специалистами торможения и все виды внутреннего торможения у них легко получаются и хорошо держатся, было решено на данной собаке специально испытать стойкость тормозного процесса. Для этого ее отрицательный раздражитель должен был быть переделан в положительный. Превращение производилось по способу, обычно скоро приводящему к цели, т. е. при сплошном, не прерываемом применением положительного раздражителя, комбинировании отрицательного раздражителя с безусловным. Прошу припомнить то, что по этому поводу было сообщено в лекнии об отрицательной индукции. Несмотря на это, устранение тормозного процесса происходило медленно. Хотя 3 дня старый отрицательный раздражитель сопровождался кормлением каждый раз сплошь от 4 до 7 раз, первый намек на устранение тормозного процесса обнаружился только на семнадцатом повторении этой процедуры и лишь в виде небольшой секреции, но без пищевой двигательной реакции. К двадцать седьмому повторению слюноотделительная реакция метронома 60 достигла значительной величины. В это время никакого резкого нарушения остальных положительных рефлексов заметить было нельзя, может быть, имелось только некоторое сближение по слюноотделительному эффекту сильных и слабых условных раздражителей.
Вот опыт, относящийся к этому времени, -- 14 XII 1925.
Ввиду того, что собаки этого типа являются как бы специалистами торможения и все виды внутреннего торможения у них легко получаются и хорошо держатся, было решено на данной собаке специально испытать стойкость тормозного процесса. Для этого ее отрицательный раздражитель должен был быть переделан в положительный. Превращение производилось по способу, обычно скоро приводящему к цели, т. е. при сплошном, не прерываемом применением положительного раздражителя, комбинировании отрицательного раздражителя с безусловным. Прошу припомнить то, что по этому поводу было сообщено в лекнии об отрицательной индукции. Несмотря на это, устранение тормозного процесса происходило медленно. Хотя 3 дня старый отрицательный раздражитель сопровождался кормлением каждый раз сплошь от 4 до 7 раз, первый намек на устранение тормозного процесса обнаружился только на семнадцатом повторении этой процедуры и лишь в виде небольшой секреции, но без пищевой двигательной реакции. К двадцать седьмому повторению слюноотделительная реакция метронома 60 достигла значительной величины. В это время никакого резкого нарушения остальных положительных рефлексов заметить было нельзя, может быть, имелось только некоторое сближение по слюноотделительному эффекту сильных и слабых условных раздражителей.
Вот опыт, относящийся к этому времени, -- 14 XII 1925.
 Но только что достигнутая величина слюноотделительного эффекта метронома 60 не осталась постоянной. Несмотря на сопровождение подкармливанием, она сейчас же начала падать и к тридцатому повторению была нулем. Вместе с тем тотчас после применения метронома 60 стали также нулевыми почти все остальные рефлексы.
Вот иллюстрирующий это опыт 18 XII 1925.
Но только что достигнутая величина слюноотделительного эффекта метронома 60 не осталась постоянной. Несмотря на сопровождение подкармливанием, она сейчас же начала падать и к тридцатому повторению была нулем. Вместе с тем тотчас после применения метронома 60 стали также нулевыми почти все остальные рефлексы.
Вот иллюстрирующий это опыт 18 XII 1925.
 Собака по внешнему виду совершенно здорова и вне станка ест ту же пищу с жадностью, как и в станке в этом опыте при первом условном раздражителе до применения метронома 60.
Дальше, хотя положительное действие метронома 60 несколько восстанавливается, его сильное задерживающее действие на остальные условные рефлексы продолжается. Однако эти рефлексы, как только опыт ставится без участия метронома, оказываются вполне нормальными, может быть только слабые раздражители к концу опыта снижаются немного больше, чем обыкновенно.
Вот опыт, доказывающий это, --- 24 XII 1925.
Собака по внешнему виду совершенно здорова и вне станка ест ту же пищу с жадностью, как и в станке в этом опыте при первом условном раздражителе до применения метронома 60.
Дальше, хотя положительное действие метронома 60 несколько восстанавливается, его сильное задерживающее действие на остальные условные рефлексы продолжается. Однако эти рефлексы, как только опыт ставится без участия метронома, оказываются вполне нормальными, может быть только слабые раздражители к концу опыта снижаются немного больше, чем обыкновенно.
Вот опыт, доказывающий это, --- 24 XII 1925.
 Я умышленно привожу несколько опытов с нормальными рефлексами, чтобы показать, как долго и упорно сохраняется норма, несмотря на повторяющееся нарушающее действие метрономных рефлексов. И дальше остается то же положение. Оба ритма метронома при постоянных колебаниях их положительного эффекта от 0.5 до 7.5 неизменно влекут за собой в течение опыта, когда они применяются, нарушение всех следующих за ними условных раздражителей в виде полного торможения их и разных переходных фаз к нему. Интересно, что часто при этом метроном 120 производит более глубокое нарушение, чем метроном 60.
Представляю несколько примеров из этого периода.
Я умышленно привожу несколько опытов с нормальными рефлексами, чтобы показать, как долго и упорно сохраняется норма, несмотря на повторяющееся нарушающее действие метрономных рефлексов. И дальше остается то же положение. Оба ритма метронома при постоянных колебаниях их положительного эффекта от 0.5 до 7.5 неизменно влекут за собой в течение опыта, когда они применяются, нарушение всех следующих за ними условных раздражителей в виде полного торможения их и разных переходных фаз к нему. Интересно, что часто при этом метроном 120 производит более глубокое нарушение, чем метроном 60.
Представляю несколько примеров из этого периода.





 Так как последний опыт показал, что рефлексы в опытах без участия раздражений метрономом стали к концу опыта падать, хотя условные раздражители и сохраняли между собой нормальные отношения, то в течение нескольких дней они применялись совпадающими, а метрономные совсем не употреблялись. После того во всех рефлексах изолированное условное раздражение (т. е. до присоединения безусловного раздражителя) продолжалось только 15 секунд, а не 20 секунд, как раньше. Кроме того, за этот период прибавлен новый условный раздражитель бульканье (звук при прохождении пузырьков воздуха через воду), принадлежащий к группе сильных раздражителей. Рефлексы увеличились и не падали к концу опыта. Теперь снова через промежуток в 11 дней было введено в опыт раздражение метрономом и именно при 120 ударах в минуту.
Вот этот опыт 2 III 1926.
Так как последний опыт показал, что рефлексы в опытах без участия раздражений метрономом стали к концу опыта падать, хотя условные раздражители и сохраняли между собой нормальные отношения, то в течение нескольких дней они применялись совпадающими, а метрономные совсем не употреблялись. После того во всех рефлексах изолированное условное раздражение (т. е. до присоединения безусловного раздражителя) продолжалось только 15 секунд, а не 20 секунд, как раньше. Кроме того, за этот период прибавлен новый условный раздражитель бульканье (звук при прохождении пузырьков воздуха через воду), принадлежащий к группе сильных раздражителей. Рефлексы увеличились и не падали к концу опыта. Теперь снова через промежуток в 11 дней было введено в опыт раздражение метрономом и именно при 120 ударах в минуту.
Вот этот опыт 2 III 1926.
 Мы видим, что введение в опыт метронома 120 (прошу помнить, давнего положительного раздражителя) сейчас же обусловило опять нарушение всех остальных условных рефлексов. Сразу наступила уравнительная фаза, переходящая в парадоксальную. Но этого мало. На следующий день, и на долго непрекращающийся срок, корковые клетки пришли в такое состояние, что они не могли выносить сильных раздражений без того, чтобы не переходить в полное тормозное состояние. Факт, что более глубокое нарушение нервной деятельности наступает не сразу, а день и больше спустя после вредного влияния, мы наблюдали очень часто в опыте c нервными патологическими состояниями.
Вот опыт на следующий день день - - 3 III 1926.
Мы видим, что введение в опыт метронома 120 (прошу помнить, давнего положительного раздражителя) сейчас же обусловило опять нарушение всех остальных условных рефлексов. Сразу наступила уравнительная фаза, переходящая в парадоксальную. Но этого мало. На следующий день, и на долго непрекращающийся срок, корковые клетки пришли в такое состояние, что они не могли выносить сильных раздражений без того, чтобы не переходить в полное тормозное состояние. Факт, что более глубокое нарушение нервной деятельности наступает не сразу, а день и больше спустя после вредного влияния, мы наблюдали очень часто в опыте c нервными патологическими состояниями.
Вот опыт на следующий день день - - 3 III 1926.
 Собака по общему виду совершенно здорова.
Такое положение дела остается 11 дней. Тогда мы применяем следующую меру. Сильный тон совсем отменяем, а звонок и бульканье значительно заглушаем.
Вот такой опыт 15 III 1926.
Собака по общему виду совершенно здорова.
Такое положение дела остается 11 дней. Тогда мы применяем следующую меру. Сильный тон совсем отменяем, а звонок и бульканье значительно заглушаем.
Вот такой опыт 15 III 1926.
 Совершенно в таком роде опыты продолжаются 9 дней. Теперь снова применяются сильные раздражители, и вот результат.
Совершенно в таком роде опыты продолжаются 9 дней. Теперь снова применяются сильные раздражители, и вот результат.
 На свободе держится совершенно нормально и пищу ест с жадностью. Через день снова применяются только слабые раздражители, и все рефлексы налицо.
На свободе держится совершенно нормально и пищу ест с жадностью. Через день снова применяются только слабые раздражители, и все рефлексы налицо.
 Выдающиеся содержательность и важность этих опытов должны оправдать, быть может, чрезмерную передачу протокольного материала.
Итак, коротко говоря, что мы имеем в данном материале? Превращение в звуковом анализаторе одного тормозного пункта в раздражимый произошло медленно и несовершенно, а главное - сделало этот пункт ненормальным, действие на который соответствующим раздражителем сейчас же приводит всю кору в патологическое состояние, выражающееся в том, что она не может раздражаться сильными раздражите.ями без того, чтобы не переходить в разные фазы тормозного состояния, до полного торможения включительно. Сначала это состояние коры довольно скоро при отмене раздражения ненормального пункта возвращается в норме, а, наконец, при дальнейшем повторении его делается стационарным. Так как другие звуковые раздражители сами по себе действуют вполне нормально, то надо принять, что в данном случае имеется частичное, узко локализированное нарушение звукового анализатора, так сказать, хроническая функциональная язва его, прикосновение к которой адекватным раздражителем вредно отзывается на всей коре, вызывая в ней, наконец, длительное патологическое состояние.
В этом факте нельзя не видеть яркого, осязательного доказательства мозаичного строения коры, о котором шла речь раньше.
Описанное окончательное нарушение деятельности всей коры можно представлять себе происшедшим двумя механизмами. Или раздражитель, вызывая в ненормальном пункте процесс раздражения, тем самым углубляет в нем процесс торможения и делает его здесь на некоторый срок стационарным, а торможение, иррадиируя, вовлекает в то же состояние и остальные клетки коры. Или же раздражитель на этот пункт действует как разрушительный агент, и тогда этот пункт, как и всякий разрушаемый пункт тела, в силу механизма внешнего торможения, влечет за собой торможение остальных частей полушарий. Образование ненормального пункта есть, очевидно, результат столкновения процессов раздражения и торможения.
К настоящему времени мы располагаем, кроме приведенных в текущей и прошлой лекциях, еще очень большим количеством случаев, в которых при трудной встрече противоположных нервных процессов наступало более или менее продолжительное, часто не поддающееся никаким нашиам, отклонение от нормы деятельности коры то в сторону преобладания раздражительного, то в сторону тормозного процесса. Происходило это или еще при выработке трудных дифференцировок, специально при дифференцировании последовательных комплексных раздражителей (опыты А. Г. Иванова-Смоленского, М. Н. Юрман и A. М. Зимкиной), или то при непосредственной, то при скорой смене в кожном анализаторе тормозного раздражителя на положительный, специально при смене одной частоты на другую механических раздражений кожи на одном и том же месте (когда они были сделаны возбудителями противоположных процессов). В последнем случае при очень возбудимом и даже агрессивном типе (опыт Л. Н. Федорова) возбуждение собаки доходило до невозможности с ней дальше работать. И лишь постоянное применение бромистого кальция и исключение из опыта даже положительного кожно-механического раздражителя вернули собаку к нормальному положению по отношению к остальным условным раздражителям. А у собаки более тормозного типа (опыт Петровой), можно было думать, при этом так же образовался узко локализированный, как в вышеприведенных опытах (Рикмана), ненормальный пункт в кожном анализаторе, положительное раздражение которого вызывало всякий раз в текущем опыте, и иногда даже и в опытах ближайших дней, разлитое торможение всей коры. К сожалению, дальнейший анализ этого случая был прерван серьезным заболеванием собаки (нефритом).
Из области описываемых патологических состояний коры мне остается описать еще несколько случаев с другим механизмом происхождения этих состоянии. этих случаях особенно незаметно патологическое сливается с нормальным, или, правильнее сказать, оно является в них постоянным свойством слабой от рождения нервной системы. Но прежде чем перейти к ним, нелишне ради ясности, хотя бы коротко, остановиться на вопросе о внешних раздражителях, действующих прямо тормозящим образом на клетки коры полушарий. Это три рода раздражителей: слабые, однообразно повторяющиеся, очень сильные и необычные, представляемые вообще или новыми явлениями, или новой связью, новой последовательностью старых явлений.
Наша жизнь и жизнь животных слишком переполнена случаями такого действия этих раздражителей, чтобы была надобность здесь приводить примеры их. Биологическое значение этого факта более или менее ясно. Если раздражители значительной силы и, главное, постоянно сменяющиеся обусловливают и должны обусловливать деятельное состояние коры, чтобы поддерживать тонкое уравновешивание организма с окружающей средой, то, естественно, слабые и однообразные, не требующие от организма никакой деятельности, должны располагать к торможению, к покою, чтобы дать корковым клеткам время для их восстановления после работы. Тормозящее действие сильных раздражителей есть, очевидно, специальный пассивно-оборонительный рефлекс, как, например, при так называемом гипнозе животных (собственно настоящем гипнозе), так как неподвижность животного, с одной стороны, делает его менее заметным для врага, а с другой - устраняет или умеряет агрессивную реакцию сильного соперника. Наконец вообще необычная обстановка должна ограничивать прежнюю энергию движений животного, так как при новом положении вещей прежний образ действия как, может быть, теперь несоответственный мог бы повести к какому-нибудь вреду для животного. Таким образом при новом, хотя бы и незначительном, колебании окружающей среды обыкновенно имеют место два рефлекса: положительный исследовательский и тормозной, так сказать, рефлекс сдержанности. осторожности. Остается интересным вопросом: самостоятельны ли оба эти рефлекса, или второй есть следствие первого в силу механизма внешнего торможения? На первый взгляд вероятнее второе. Физиологический механизм тормозящего действия всех трех родов раздражителей будет предметом обсуждения в одной из последних лекций.
Необыкновенное стихийное событие, именно - очень большое наводнение, случившееся в Ленинграде 23 сентября 1924 года, представило нам случай наблюдать и изучать хроническое патологическое состояние нервной системы наших собак, происшедшее под влиянием этого события как чрезвычайно сильного внешнего раздражителя. Помещение для животных, стоявшее на земле и находившееся в расстоянии четверти километра от здания лаборатории, стало заливать водой. При страшной буре, при сильном волнении прибывающей воды, при ударах больших волн о здания, при треске и шуме ломаемых и падающих деревьев пришлось спешно, партиями и вплавb, переводить наших животных во второй этаж лаборатории и оставлять их здесь в необычных компаниях. Все это, очевидно, чрезвычайно затормозиловсех без исключения животных, так как в это время между ними не замечалось обычных драк. Одни из собак, по миновании события и водворении их на прежнее место, остались такими, какими были; другие, и именно тормозного типа, оказались после этого нервно-больными и на очень большой срок, в чем мы убедились опытами над их условными рефлексами. К этим опытам я и перехожу теперь.
Первая собака (опыты Сперанского), уже упоминавшаяся раньше, сильное здоровое животное, но очень тормозимое. У нее все условные рефлексы были очень значите.ьны, особенно постоянны и точны, но при условии, что экспериментальная обстановка оставалась строго обычной. Напоминаю, что рефлексов (пищевых) было у нее 10, из них 6 положительных и 4 отрицательных (дифференцировки); из положительных 3 звуковых и 3 оптических; из звуковых самый сильный звонок - вызывал самое большое слюноотделение, оптические по этому эффекту были равны между собой, и, наконец, положительные звуковые рефлексы превосходили оптические по слюноотделению на треть и больше. Через неделю после наводнения поставленная в станок собака держалась в нем необычно беспокойно, условных рефлексов почти не было, и собака, вообще очень жадная, еду не брала, даже отворачивалась от нее. Так стояло дело 3 дня. Собаку оставили без еды 3 дальнейших дня, но это не изменило результата. Исключив другие причины и на основании некоторых наблюдений над собакой, мы пришли ключению, что это продолжающееся действие наводнения, и приняли такую меру. Экспериментатор, обыкновенно производивший опыт за дверью комнаты, где собака находилась одна, теперь остался с ней в комнате, а я из-за двери вел опыт. Рефлексы сейчас же появились, и собака жадно ела подаваемую ей ееу. Но стоило экспериментатору оставить собаку одну, повторилось то, что было. Чтобы восстановить рефлексы, пришлось систематически то оставаться в комнате, то выходить из нее на некоторое время. На одиннадцатый день опытов был впервые применен остававшийся до сих пор неупотребленным раздражитель - звонок, самый действительный по условному эффекту и вместе физически самый сильный. Все остальные рефлексы после этого сразу упали, и еду собака перестала брать; при этом стала очень возбужденной, беспокойно озиралась кругом и особенно упорно смотрела со станка на пол. Применение социального раздражителя, т. е. присутствие экспериментатора около собаки, постепенно восстановило рефлексы, но повторенный через 5 дней звонок вызвал то же самое. Тогда звонок стали применять только при нахождении экспериментатора в комнате с собакой. Медленно восстановлялись нормальные отношения. На рефлексах замечалась часто уравнительная фаза, при звонке еда иногда прерывалась, после звонка часто падали остальные рефлексы. Наконец на сорок седьмой день опытов, почти через 2 месяца после наводнения, получился вполне нормальный опыт. В течение его был испробован следующий прием. Из-под двери в комнату собаки была пущена тихая струя воды, которая образовала небольшую лужу около стола, на котором в станке стояла собака.
Вот этот опыт (17 XI 1924), веденный как в норме, без экспериментатора в комнате, где находилась собака.
Выдающиеся содержательность и важность этих опытов должны оправдать, быть может, чрезмерную передачу протокольного материала.
Итак, коротко говоря, что мы имеем в данном материале? Превращение в звуковом анализаторе одного тормозного пункта в раздражимый произошло медленно и несовершенно, а главное - сделало этот пункт ненормальным, действие на который соответствующим раздражителем сейчас же приводит всю кору в патологическое состояние, выражающееся в том, что она не может раздражаться сильными раздражите.ями без того, чтобы не переходить в разные фазы тормозного состояния, до полного торможения включительно. Сначала это состояние коры довольно скоро при отмене раздражения ненормального пункта возвращается в норме, а, наконец, при дальнейшем повторении его делается стационарным. Так как другие звуковые раздражители сами по себе действуют вполне нормально, то надо принять, что в данном случае имеется частичное, узко локализированное нарушение звукового анализатора, так сказать, хроническая функциональная язва его, прикосновение к которой адекватным раздражителем вредно отзывается на всей коре, вызывая в ней, наконец, длительное патологическое состояние.
В этом факте нельзя не видеть яркого, осязательного доказательства мозаичного строения коры, о котором шла речь раньше.
Описанное окончательное нарушение деятельности всей коры можно представлять себе происшедшим двумя механизмами. Или раздражитель, вызывая в ненормальном пункте процесс раздражения, тем самым углубляет в нем процесс торможения и делает его здесь на некоторый срок стационарным, а торможение, иррадиируя, вовлекает в то же состояние и остальные клетки коры. Или же раздражитель на этот пункт действует как разрушительный агент, и тогда этот пункт, как и всякий разрушаемый пункт тела, в силу механизма внешнего торможения, влечет за собой торможение остальных частей полушарий. Образование ненормального пункта есть, очевидно, результат столкновения процессов раздражения и торможения.
К настоящему времени мы располагаем, кроме приведенных в текущей и прошлой лекциях, еще очень большим количеством случаев, в которых при трудной встрече противоположных нервных процессов наступало более или менее продолжительное, часто не поддающееся никаким нашиам, отклонение от нормы деятельности коры то в сторону преобладания раздражительного, то в сторону тормозного процесса. Происходило это или еще при выработке трудных дифференцировок, специально при дифференцировании последовательных комплексных раздражителей (опыты А. Г. Иванова-Смоленского, М. Н. Юрман и A. М. Зимкиной), или то при непосредственной, то при скорой смене в кожном анализаторе тормозного раздражителя на положительный, специально при смене одной частоты на другую механических раздражений кожи на одном и том же месте (когда они были сделаны возбудителями противоположных процессов). В последнем случае при очень возбудимом и даже агрессивном типе (опыт Л. Н. Федорова) возбуждение собаки доходило до невозможности с ней дальше работать. И лишь постоянное применение бромистого кальция и исключение из опыта даже положительного кожно-механического раздражителя вернули собаку к нормальному положению по отношению к остальным условным раздражителям. А у собаки более тормозного типа (опыт Петровой), можно было думать, при этом так же образовался узко локализированный, как в вышеприведенных опытах (Рикмана), ненормальный пункт в кожном анализаторе, положительное раздражение которого вызывало всякий раз в текущем опыте, и иногда даже и в опытах ближайших дней, разлитое торможение всей коры. К сожалению, дальнейший анализ этого случая был прерван серьезным заболеванием собаки (нефритом).
Из области описываемых патологических состояний коры мне остается описать еще несколько случаев с другим механизмом происхождения этих состоянии. этих случаях особенно незаметно патологическое сливается с нормальным, или, правильнее сказать, оно является в них постоянным свойством слабой от рождения нервной системы. Но прежде чем перейти к ним, нелишне ради ясности, хотя бы коротко, остановиться на вопросе о внешних раздражителях, действующих прямо тормозящим образом на клетки коры полушарий. Это три рода раздражителей: слабые, однообразно повторяющиеся, очень сильные и необычные, представляемые вообще или новыми явлениями, или новой связью, новой последовательностью старых явлений.
Наша жизнь и жизнь животных слишком переполнена случаями такого действия этих раздражителей, чтобы была надобность здесь приводить примеры их. Биологическое значение этого факта более или менее ясно. Если раздражители значительной силы и, главное, постоянно сменяющиеся обусловливают и должны обусловливать деятельное состояние коры, чтобы поддерживать тонкое уравновешивание организма с окружающей средой, то, естественно, слабые и однообразные, не требующие от организма никакой деятельности, должны располагать к торможению, к покою, чтобы дать корковым клеткам время для их восстановления после работы. Тормозящее действие сильных раздражителей есть, очевидно, специальный пассивно-оборонительный рефлекс, как, например, при так называемом гипнозе животных (собственно настоящем гипнозе), так как неподвижность животного, с одной стороны, делает его менее заметным для врага, а с другой - устраняет или умеряет агрессивную реакцию сильного соперника. Наконец вообще необычная обстановка должна ограничивать прежнюю энергию движений животного, так как при новом положении вещей прежний образ действия как, может быть, теперь несоответственный мог бы повести к какому-нибудь вреду для животного. Таким образом при новом, хотя бы и незначительном, колебании окружающей среды обыкновенно имеют место два рефлекса: положительный исследовательский и тормозной, так сказать, рефлекс сдержанности. осторожности. Остается интересным вопросом: самостоятельны ли оба эти рефлекса, или второй есть следствие первого в силу механизма внешнего торможения? На первый взгляд вероятнее второе. Физиологический механизм тормозящего действия всех трех родов раздражителей будет предметом обсуждения в одной из последних лекций.
Необыкновенное стихийное событие, именно - очень большое наводнение, случившееся в Ленинграде 23 сентября 1924 года, представило нам случай наблюдать и изучать хроническое патологическое состояние нервной системы наших собак, происшедшее под влиянием этого события как чрезвычайно сильного внешнего раздражителя. Помещение для животных, стоявшее на земле и находившееся в расстоянии четверти километра от здания лаборатории, стало заливать водой. При страшной буре, при сильном волнении прибывающей воды, при ударах больших волн о здания, при треске и шуме ломаемых и падающих деревьев пришлось спешно, партиями и вплавb, переводить наших животных во второй этаж лаборатории и оставлять их здесь в необычных компаниях. Все это, очевидно, чрезвычайно затормозиловсех без исключения животных, так как в это время между ними не замечалось обычных драк. Одни из собак, по миновании события и водворении их на прежнее место, остались такими, какими были; другие, и именно тормозного типа, оказались после этого нервно-больными и на очень большой срок, в чем мы убедились опытами над их условными рефлексами. К этим опытам я и перехожу теперь.
Первая собака (опыты Сперанского), уже упоминавшаяся раньше, сильное здоровое животное, но очень тормозимое. У нее все условные рефлексы были очень значите.ьны, особенно постоянны и точны, но при условии, что экспериментальная обстановка оставалась строго обычной. Напоминаю, что рефлексов (пищевых) было у нее 10, из них 6 положительных и 4 отрицательных (дифференцировки); из положительных 3 звуковых и 3 оптических; из звуковых самый сильный звонок - вызывал самое большое слюноотделение, оптические по этому эффекту были равны между собой, и, наконец, положительные звуковые рефлексы превосходили оптические по слюноотделению на треть и больше. Через неделю после наводнения поставленная в станок собака держалась в нем необычно беспокойно, условных рефлексов почти не было, и собака, вообще очень жадная, еду не брала, даже отворачивалась от нее. Так стояло дело 3 дня. Собаку оставили без еды 3 дальнейших дня, но это не изменило результата. Исключив другие причины и на основании некоторых наблюдений над собакой, мы пришли ключению, что это продолжающееся действие наводнения, и приняли такую меру. Экспериментатор, обыкновенно производивший опыт за дверью комнаты, где собака находилась одна, теперь остался с ней в комнате, а я из-за двери вел опыт. Рефлексы сейчас же появились, и собака жадно ела подаваемую ей ееу. Но стоило экспериментатору оставить собаку одну, повторилось то, что было. Чтобы восстановить рефлексы, пришлось систематически то оставаться в комнате, то выходить из нее на некоторое время. На одиннадцатый день опытов был впервые применен остававшийся до сих пор неупотребленным раздражитель - звонок, самый действительный по условному эффекту и вместе физически самый сильный. Все остальные рефлексы после этого сразу упали, и еду собака перестала брать; при этом стала очень возбужденной, беспокойно озиралась кругом и особенно упорно смотрела со станка на пол. Применение социального раздражителя, т. е. присутствие экспериментатора около собаки, постепенно восстановило рефлексы, но повторенный через 5 дней звонок вызвал то же самое. Тогда звонок стали применять только при нахождении экспериментатора в комнате с собакой. Медленно восстановлялись нормальные отношения. На рефлексах замечалась часто уравнительная фаза, при звонке еда иногда прерывалась, после звонка часто падали остальные рефлексы. Наконец на сорок седьмой день опытов, почти через 2 месяца после наводнения, получился вполне нормальный опыт. В течение его был испробован следующий прием. Из-под двери в комнату собаки была пущена тихая струя воды, которая образовала небольшую лужу около стола, на котором в станке стояла собака.
Вот этот опыт (17 XI 1924), веденный как в норме, без экспериментатора в комнате, где находилась собака.
 Теперь в 11 часов 59 минут была пущена вода в комнату собаки.
Теперь в 11 часов 59 минут была пущена вода в комнату собаки.
 Много месяцев спустя, когда рефлексы были налицо, а звонок умышленно долго не употреблялся, применение его сначала дало обычный, превосходивший другие рефлексы, эффект. Но при повторении в небольшом ряде дней и только по разу в день звонок постепенно терял свое действие, стал нулевым и сильно понизил все остальные рефлексы. Интересно, что в это время восстановляющим образом на рефлексы действовал не только сам экспериментатор, но и его одежда, положенная около собаки и ею не видимая, следовательно ее запах.
Таким образом под влиянием чрезвычайного раздражителя клетки коры, и ранее очень склонные переходить в тормозное состояние, хронически сделались еще более тормозимыми. Раздражители, давно ставшие индифферентными (обстановка опыта), а также сильные агенты, ранее уже бывшие сильными, положительно действующими условными раздражителями (звонок), теперь резко тормозили эти ослабленные клетки. Ничтожные энергетически компоненты того чрезвычайного раздражителя (наводнения) были достаточны, чтобы вызвать первоначальную реакцию.
Другая собака (опыты Рикмана) была та самая, опыты над которой были подробно приведены в начале этой лекции. Описываемое теперь состояние собаки относится к более раннему времени. Влияние на нее наводнения выразилось в несколько иной патологической форме, но с тем же основным механизмом, т. е. в виде чрезвычайной тормозимости.
Вот опыт накануне наводнения - 22 IX 1924.
Много месяцев спустя, когда рефлексы были налицо, а звонок умышленно долго не употреблялся, применение его сначала дало обычный, превосходивший другие рефлексы, эффект. Но при повторении в небольшом ряде дней и только по разу в день звонок постепенно терял свое действие, стал нулевым и сильно понизил все остальные рефлексы. Интересно, что в это время восстановляющим образом на рефлексы действовал не только сам экспериментатор, но и его одежда, положенная около собаки и ею не видимая, следовательно ее запах.
Таким образом под влиянием чрезвычайного раздражителя клетки коры, и ранее очень склонные переходить в тормозное состояние, хронически сделались еще более тормозимыми. Раздражители, давно ставшие индифферентными (обстановка опыта), а также сильные агенты, ранее уже бывшие сильными, положительно действующими условными раздражителями (звонок), теперь резко тормозили эти ослабленные клетки. Ничтожные энергетически компоненты того чрезвычайного раздражителя (наводнения) были достаточны, чтобы вызвать первоначальную реакцию.
Другая собака (опыты Рикмана) была та самая, опыты над которой были подробно приведены в начале этой лекции. Описываемое теперь состояние собаки относится к более раннему времени. Влияние на нее наводнения выразилось в несколько иной патологической форме, но с тем же основным механизмом, т. е. в виде чрезвычайной тормозимости.
Вот опыт накануне наводнения - 22 IX 1924.
 На третий день после наводнения, 26 IX 1924, опыт шел следующим образом:
На третий день после наводнения, 26 IX 1924, опыт шел следующим образом:
 Собака еду брала, но положительные рефлексы были очень ослаблены, и больше всего положительно действовал отрицательный раздражитель (ультрапарадоксальная фаза).
Затем надолго установилось следующее положение. Пока не применялся отрицательный раздражитель (метроном 60), положительные раздражители были удовлетворительны и достигали часто нормальной величины. Но даже однократное применение его на все остальное время опыта и на многие следующие дни делало все условные рефлексы нулевыми или очень уменьшенными. Вот два примера.
Собака еду брала, но положительные рефлексы были очень ослаблены, и больше всего положительно действовал отрицательный раздражитель (ультрапарадоксальная фаза).
Затем надолго установилось следующее положение. Пока не применялся отрицательный раздражитель (метроном 60), положительные раздражители были удовлетворительны и достигали часто нормальной величины. Но даже однократное применение его на все остальное время опыта и на многие следующие дни делало все условные рефлексы нулевыми или очень уменьшенными. Вот два примера.
 В периоде восстановления рефлексов, при исключении отрицательного раздражителя, наблюдались всереходные фазы между полным торможением и нормой. Сначала восстановлению помогали наши наиболее действительные приемы: перерыв на несколько дней опытов и укорочение изолированного действия условных раздражителей, но, наконец, они оказывались недостаточными. Только один-два рефлекса в начале опыта обладали небольшим эффектом, а все остальные были нулевыми. Собака становилась неподвижной и упорно отказывалась от пищи. Пришлось применить последнюю меру. Опыт производился не в станке, а на полу, на свободе, чем для таких собак частью устранялось некоторое тормозящее действие станка, частью прибавлялось для полушарий несколько раздражающих импульсов, идущих из двигательного аппарата. Этомогло. Рефлексы стали возвращаться и увеличиваться. Собака теперь брала еду. Наконец была достигнута норма. Снова примененный теперь отрицательный раздражитель только в первые 7 дней повлек за собой исчезание всех рефлексов до конца опыта, но не распространил своего тормозящего действия на следующие дни. Затем в течение 2 недель постепенно устранилось и это влияние. После этого начали осторожно практиковать торможение. Дифференцировочное торможение повторялось в одном и том же опыте, концентрировалось применением вскоре после него положительного раздражителя и, наконец, утончалось. Только после 2 месяцер опытов на полу и спустя 8 месяцев после наводнения можно было вернуться к опытам в обыкновенных условиях на станке.
Таким образом наводнение обусловило такую тормозимость корковых клеток, что незначительно с нашей стороны в виде отрицательного раздражителя делало долгое время невозможным существование положительных условных рефлексов в нашей обычной обстановке опытов.
Итак, по нашим опытам и наблюдениям, хроническое патологическое состояние больших полушарий могут производить два фактора: трудная встреча, столкновение процессов раздражения и торможения и сильные чрезвычайные раздражители.
Но я должен сделать еще сообщение о собаке (опыты А. С. Вишневского), относительно которой я, к сожалению, определенно сказать не могу: есть ли ее настоящее состояние прирожденное состояние, изменившиеся под влиянием общих жизненных условий, возраста, родов и пр., или тоже, как в двух предшествующих случаях, специальный результат влияния наводнения. Это - собака, описанная в прошлой лекции как крайний представитель крайнего тормозного типа. Перед наводнением она долго оставалась без наблюдений, как и после наводнения стала предметом изучения только спустя 3-4 месяца. Задолго до наводнения над ней, как уже сказано раньше, было произведено. немало даже очень ценных опытов условными рефлексами. В настоящее время, несмотря на все меры, для опытов на наши обычные темы она негодна. Можно анализировать только ее состояние. Какая этому причина: наводнение или что другое, остается для нас нерешенным вопросом. Полоса ее нормальной, обычной для других собак, жизни, по крайней мере в условиях лаборатории, в высшей степени сужена. В лаборатории почти исключительно она или сплошь обнаруживает пассивно-оборонительный рефлекс, т. е. отвечает ориентировочной реакцией на все самые незначительные колебания среды и тотчас за этим тормозит свои движения до отказа от еды, или, как исключение из своего типа, спит. Только два приема возвращают ее к обычному для других собак состоянию. Это очень быстрый переход через 1-2 секунды от начала условного раздражителя к подаче еды и ведение опыта на полу, при условии хождения экспериментатора по комнате, причем собака неотступно ходит за ним, но и при этом за условным раздражителем скоро должно следовать подкармливание. То и другое изменяет состояние собаки в том отношении, что собака перестает так реагировать на малые колебания среды, как раньше, и может есть подаваемую ей пищу, прерывая еду только при значительных внешних раздражениях. Теперь начинают появляться и условные рефлексы. Но стоит отодвинуть даже только на 5-10 секунд подачу еды от начала условного раздражителя, как собака очень скоро становится сонлевой и может даже заснуть над кормушкой во время еды. Такое совершенно исключительное состояние нервной системы, очевидно именно клеток коры полушарий, мы представляем себе как состояние крайнего истощения этих клеток, как высшее выражение так называемой раздражительной слабости их. Прибавляю слово «очевидно» потому, что такая тонкая реактивность нервной системы, и именно всех ее анализаторов, доступна только большим полушариям. Подробным изучением этого случая мы занять; в настоящее время.
В периоде восстановления рефлексов, при исключении отрицательного раздражителя, наблюдались всереходные фазы между полным торможением и нормой. Сначала восстановлению помогали наши наиболее действительные приемы: перерыв на несколько дней опытов и укорочение изолированного действия условных раздражителей, но, наконец, они оказывались недостаточными. Только один-два рефлекса в начале опыта обладали небольшим эффектом, а все остальные были нулевыми. Собака становилась неподвижной и упорно отказывалась от пищи. Пришлось применить последнюю меру. Опыт производился не в станке, а на полу, на свободе, чем для таких собак частью устранялось некоторое тормозящее действие станка, частью прибавлялось для полушарий несколько раздражающих импульсов, идущих из двигательного аппарата. Этомогло. Рефлексы стали возвращаться и увеличиваться. Собака теперь брала еду. Наконец была достигнута норма. Снова примененный теперь отрицательный раздражитель только в первые 7 дней повлек за собой исчезание всех рефлексов до конца опыта, но не распространил своего тормозящего действия на следующие дни. Затем в течение 2 недель постепенно устранилось и это влияние. После этого начали осторожно практиковать торможение. Дифференцировочное торможение повторялось в одном и том же опыте, концентрировалось применением вскоре после него положительного раздражителя и, наконец, утончалось. Только после 2 месяцер опытов на полу и спустя 8 месяцев после наводнения можно было вернуться к опытам в обыкновенных условиях на станке.
Таким образом наводнение обусловило такую тормозимость корковых клеток, что незначительно с нашей стороны в виде отрицательного раздражителя делало долгое время невозможным существование положительных условных рефлексов в нашей обычной обстановке опытов.
Итак, по нашим опытам и наблюдениям, хроническое патологическое состояние больших полушарий могут производить два фактора: трудная встреча, столкновение процессов раздражения и торможения и сильные чрезвычайные раздражители.
Но я должен сделать еще сообщение о собаке (опыты А. С. Вишневского), относительно которой я, к сожалению, определенно сказать не могу: есть ли ее настоящее состояние прирожденное состояние, изменившиеся под влиянием общих жизненных условий, возраста, родов и пр., или тоже, как в двух предшествующих случаях, специальный результат влияния наводнения. Это - собака, описанная в прошлой лекции как крайний представитель крайнего тормозного типа. Перед наводнением она долго оставалась без наблюдений, как и после наводнения стала предметом изучения только спустя 3-4 месяца. Задолго до наводнения над ней, как уже сказано раньше, было произведено. немало даже очень ценных опытов условными рефлексами. В настоящее время, несмотря на все меры, для опытов на наши обычные темы она негодна. Можно анализировать только ее состояние. Какая этому причина: наводнение или что другое, остается для нас нерешенным вопросом. Полоса ее нормальной, обычной для других собак, жизни, по крайней мере в условиях лаборатории, в высшей степени сужена. В лаборатории почти исключительно она или сплошь обнаруживает пассивно-оборонительный рефлекс, т. е. отвечает ориентировочной реакцией на все самые незначительные колебания среды и тотчас за этим тормозит свои движения до отказа от еды, или, как исключение из своего типа, спит. Только два приема возвращают ее к обычному для других собак состоянию. Это очень быстрый переход через 1-2 секунды от начала условного раздражителя к подаче еды и ведение опыта на полу, при условии хождения экспериментатора по комнате, причем собака неотступно ходит за ним, но и при этом за условным раздражителем скоро должно следовать подкармливание. То и другое изменяет состояние собаки в том отношении, что собака перестает так реагировать на малые колебания среды, как раньше, и может есть подаваемую ей пищу, прерывая еду только при значительных внешних раздражениях. Теперь начинают появляться и условные рефлексы. Но стоит отодвинуть даже только на 5-10 секунд подачу еды от начала условного раздражителя, как собака очень скоро становится сонлевой и может даже заснуть над кормушкой во время еды. Такое совершенно исключительное состояние нервной системы, очевидно именно клеток коры полушарий, мы представляем себе как состояние крайнего истощения этих клеток, как высшее выражение так называемой раздражительной слабости их. Прибавляю слово «очевидно» потому, что такая тонкая реактивность нервной системы, и именно всех ее анализаторов, доступна только большим полушариям. Подробным изучением этого случая мы занять; в настоящее время.
Лекция девятнадцатая. Патологические состояния больших полушарий как результат оперативных воздействий на них: а) общие изменения деятельности коры полушарий; 6) нарушения деятельности звукового анализатора
Мм. гг.! Получив достаточную, вполне объективную характеристику деятельности больших полушарий, мы естественно должны были перейти к подробному изучению конституции больших полушарий, к изучению функций отдельных частей их, как и было проектировано в самом начале нашей работы (см. мой доклад на Международном медицинском конгрессе в Мадриде 1903 г. в моем «Двадцатилетнем опыте объективного изучения высшей нервной деятельности животных»). Перед нами для этой цели стоял пока единственный метод, метод частичного разрушения, удаления отдельных частей полушарий, способ, явно отягченный важными недостатками. И как могло быть иначе? Большие полушария представляют собой сложнейшую и тончайшую конструкцию, произведенную творческой силой земной природы, а мы для ознакомления с ней обращаемся к ней с грубой формой разрушения, грубым валовым отторжением тех или других кусков ее. Представьте себе, что нам предстоит изучить конструкцию и работу бесконечно более простого инструмента поделки человеческих рук. И мы, невежественно не различая частей его, не разбирая его осторожно, удаляли бы, например, при помощи пилы или другого какого-либо разрушительного орудия, то восьмую, то четвертую и т. д. часть его и таким образом собирали бы материал для суждения об устройстве этого инструмента и об его работе. А таково в сущности наше обыкновенное поведение в отношении больших полушарий, как и других частей мозга. Молотком с долотом или пилой мы взламываем плотное вместилище их, вскрываем их несколько покровов, разрываем их кровеносные сосуды и производим, наконец, разнообразное механическое воздействие на них (сотрясением, давлением, растяжением), отторгая от них куски той или другой величины. Но такова специальная особенность, таково своеобразное могущество живого вещества. Несмотря на все это, даже через часы, а через сутки наверное, без специального и точного исследования иногда нельзя заметить на животном никакого уклонения от нормы, а с другой стороны, все же и с этими приемами можно нечто выяснить относительно функции больших полушарий. Однако это не должно удовлетворять, успокаивать физиолога; на нем лежит постоянная обязанность: опираясь на теперешние успехи естествознания и чрезвычайное увеличение современных технических средств, стараться изыскивать для той же цели другие приемы, не так удаленные от недосягаемого совершенства исследуемого им прибора. Естественно, что теперешний прием исследования полушарий посредством удаления частей их ведет, собственно, к сложному патологическому состоянию, и выводы из получаемых при этом результатов относительно конституции полушарий, даже очень осторожные и с большими ограничениями, все же не могут быть вполне обеспечены против вероятности быть ошибочными. Ведь полушария - специальный прибор сношений, связей, к тому же обладающий высшей реактивностью, и, следовательно, всякое нарушение его в одном месте должно давать себя знать, отражаясь на всем приборе или по крайней мере на многих отдаленных его пунктах или частях. Помимо непосредственного влияния операции, относительно которого можно было бы, и с основанием, надеяться, что оно течением времени устранится пластической, организующей работой живого вещества, остается еще серьезное зло оперативного приема, его более отдаленное следствие - это образование рубцовой ткани на месте мозгового дефекта как новой причины раздражения и дальнейшего разрушения. С одной стороны, рубец как механический раздражитель прилежащих нормальных частей мозга вызывает в нем периодические взрывы возбуждения, с другой, давя, растягивая и разрывая, он продолжает разрушать дальше и дальше мозговую массу. Я был несчастлив, рядом с некоторыми улучшениями оперативного приема, сделать при этом, как я думаю теперь, и важную ошибку. Избегая кровотечения при главной операции, -я у моих собак задолго предварительно начисто удалял темпоральный мускул, покрывающий череп. Моя цель была достигнута: атрофированный череп можно было вскрывать часто без капли крови. Но вместе с тем атрофировалась и dura mater, делаясь сухой и ломкой, так что большей частью потом не удавалось зашивать ее наглухо. Вследствие этого мозговая рана вступала в связь с раневой поверхностью посторонних тканей, с их грубым рубцеванием, и отсюда рубец легко внедрялся и разрастался все больше и больше в мозговой ткани. Почти все мной оперированные собаки кончали судорожными припадками, которые иногда наступали уже через 5-6 недель после операции. Редко смерть наступала при первых судорогах. Чаще эти судороги, сначала редкие и слабые, учащаясь и усиливаясь в течение месяцев, все же приводили животное или к смерти, илн к новому и более значительному расстройству нервной деятельности. Применяемые мной меры - то повторная наркотизация, то вырезывание рубцов - иногда помогами, но не могли сделаться, конечно, верным средством. Помимо затруднений, возникающих со стороны теперешнего способа исследования функций отдельных участков полушарий, экспериментатору приходится серьезно считаться с чертой конструкции полушарий, особенно резко выдвинутой именно в них. Если во всем организме мы постоянно встречаемся с запасными средствами против частичных нарушений его, то в нервной системе, как устанавливающей все связи и отношения организма, этот принцип должен обнаруживаться в высшей степени. И это мы видим как в спинном мозгу на сложном и очень разнообразном ходе его волокон, так и в периферической нервной системе на ее широко практикуемый так называемой возвратной чувствительности, что явно значительно нейтрализует вред механических нарушений этих отделов. Конечно, то же в высшей мере должно быть осуществлено в верховной инстанции аппарата, регулируюшего всю внутреннюю и внешнюю деятельность организма. Ввиду сказанного, располагая нашими условными рефлексами, относящимися к разным анализаторам, мы старались полностью констатировать и изучать патологические состояния полушарий после экстирпации тех или других участиов их, утилизируя это изучение по мере возможности и для вопроса об общей конституции полушарий и о значении отдельных частей их. Первое явление, которое наблюдается после частичной экстирпации полушарий, - это иссловных рефлексов. но большей частью не всех, а только называемых нами искусственными условными рефлексами, т. е. сделанных нами в лаборатории, значит, относительно новых, мало практикованных Затем постоянное правило, что натуральные условные рефлексы, например рефлекс на пищу, как двигательный, так и секреторный (слюнной), оказываются налицо раньше всех искусственных. Иногда даже можно не уловить времени их отсутствия после рассеяния наркоза, сопровождавшего операцию. Примеры относительной стойкости натуральных рефлексов сравнительно с искусственными встречаются в каждой нашей работе с частичНым разрушением полушарий, так что нет надобности приводить примеры этого особо. Обыкновенно все наши условные рефлексы после операции исчезают, хотя бы операция была произведена на одном полушарии и безразлично в том или другом пункте его. Исчезают условные рефлексы на разные сроки, от одного дня до многих недель и даже месяцев. Хотя обычно чем больше разрушение, тем длиннее период исчезания рефлексов, однако в этом отношении, беря все случаи, наблюдаем и немалые исключения. Даже при двух операциях, повидимому совершенно одинаковых как по месту и размеру, так и по исполнению, сроки исчезания рефлексов могут очень различаться. Вероятно, размер окончательного операционного раздражения и разрушения, помимо ловкости и внимательности оператора, зависит от анатомических функциональных особенностей данного оперируемого животного Возвращаются условные рефлексы не все сразу, а в известном порядке. Помимо уже упомянутой выше прочности рефлексов, срок возврата зависит от места оперирования: рефлексы анализатора, лежащего дальше от разрушаемого района, восстанавлизаются раньше. Но, кроме этого, имеют значение также, вероятно, как сила условного раздражителя, так и натура анализаторов. Так, при удалении g. pyriformis (опыты Завадского) условный рефлекс с полости рта (водяной, о котором речь позже) появился на одиннадцатый день, рефлекс на камфору - на восемнадцатый, на общее освещение комнаты - на двадцать пятый день, а кожномеханический не был восстановлен еще и на тридцать пятый день. Все эти факты свидетельствуют об известном влиянии, распространяющемся от места нарушения ио массе полушарий, а затем постепенном концентрировании его к исходному пункту. Это влияние, вообще говоря, есть операционное раздражение. Если сильные раздражители, а такое только столкновение нервных процессов производят, как мы это видели в ранних лекциях, тормозящее действие и притом продолжительное, то тем более того же можно ждать от оперативного приема, механического разрушения части мозговой конструкции. Когда условные рефлексы, наконец, восстановляются, они не только достигают нормального размера, но иногда переходят его, усиливаются и делаются устойчивее, а рядом с этим выступает ослабление процесса торможения. Этому имеются в нашем материале опять многочисленные примеры. После удаления части слуковой сферы Мунка (опыты Эльяссона) на двух собаках условные пищевые рефлексы резко увеличились и стали постоянными в течение всего опыта, когда до операции значительно падали к концу опыта. Еще резче это выступило у одной собаки после удаления задней половины полушарий (опыты А. Н. Кудрина), у которой пищевые рефлексы, дававшие за время изолированного действия условного раздражителя 1-2 капли, возросли до 13 капель за тот же срок. У многих собак наблюдается гораздо более продолжительное слюноотделение по прекращении действия безусловного раздражителя. Иногда отмечается более длительный период угасания, затрудняется образование дифференцировок, условного тормоза, и наступает промежуточное слюноотделение между применениями раздражителей, которого раньше не было. В последнем случае, вероятно, растормаживается обстановочный рефлекс, упомянутый в седьмой лекции. Остается вопросом: есть ли ослабление тормозного процесса результат усиления процесса раздражения или обратно? В тормозном процессе после операции наблюдается и еще изменение. Это - инертность, так сказать, неподвижность его. Как мы видели, тормозной процесс при его практике концентрируется и во времени и в пространстве. В послеоперационном периоде он концентрируется очень медленно и вяло. Это явление инертности торможения обнаруживается не только в том анализаторе, который подвергся нарушению, но и на других, непосредственно нетронутых (опыты Красногорского). Таким образом цель - удалив часть полушарий, видеть исчезновение функции удаленной части из общей нормальной деятельности полушарии - покрывается на первых порах отзвуком операционного удара по всей массе полушарий. Когда этот отзвук, часто только медленно и постепенно затихающий, наконец, перестает давать себя знать, скоро появляется, как уже выше сказано, новое, также общего дллушарий значения, обстоятельство - образование рубца. Влияние рубца в различных случаях очень разнообразно. Иногда при одной и той же операции работа рубца обнаруживается то очень скоро, сильно и часто, то наоборот. Последнее, к сожалению, у нас случалось реже, чем первое. Самым частым симптомом действия рубца являются судороги всего тела или местные. Эти взрывы возбуждения в полушариях дают себя знать и вне периода судорог, в изменении нормальной деятельности полушарий. Мы должны различать изменения, сопровождающие взрывы, менее значительные и отдельные, и изменения, являющиеся результатом сильнейших взрывов или и не очень сильных, но часто повторяющихся. Начнем с первых. Исследуя изо дня в день условные рефлексы, можно довольно точно заранее предсказывать наступление судорог. Если вдруг без всякой видимой причины условные рефлексы начинают падать и, наконец, исчезают, то это знак приближающихся судорог. Иногда можно уловить более раннюю стадию - исчезание дифференцировок, т. е. нарушение тормозного процесса. Когда судорожные приступы минуют, восстановление условных рефлексов происходит в очень разные сроки: то через часы, то через дни. Иногда восстановление принимает более сложный вид. Сейчас же после судорог рефлексы есть, а затем они снова исчезают на довольно продолжительный срок. Вероятно, сначала на некоторое время существует иррадиация взрыва возбуждения, а затем только начинается его концентрирование с отрицательной индукцией. Что какается очень сильных взрывов возбуждения (сильных судорог) или многократно и очень часто повторяющихся, то последовательное действие их очень разнообразно. У одной собаки, вероятно после них, сразу получилась абсолютная глухота. Другая собака, державшаяся ранее совершенно нормально относительно людей, других собак и пищи, после судорог сторонилась, бежала от всего этого, пока новый приступ судорог не убил ее. Третья после многократных и частых судорог представила ряд совершенно особенных симптомов. Я опишу ее подробно позже, здесь же остановлюсь только на одном ее симптоме. Все рефлексы у нее после судорог восстанавливались, но они должны были быть почти совпадающими. Сколько-нибудь значительное (более 5 секунд) удлинение и повторное применение удлиненного изолированного условного раздражения быстро вело к иссловного действия. к развитию сонливости и отказу от пищи. Ясно, что собака впала в хроническое состояние раздражительной слабости, которое описано на собаке, приведенной в конце прошлой лекции. После каждого повторения судорог это состояние усиливалось. Совершенно естественно понимать это явление как наступающее после сильных взрывов возбуждения истощение корковых клеток, которые под влиянием внешних раздражителей стремительно переходят в тормозное состояние. В данном случае, в связи с общим возбуждением полушарий, это состояние охватывает всю массу полушарий; с частичным же проявлением его на отдельных анализаторах, подвергающихся нарушению, мы встретимся не раз дальше. Связь этого состояния с сонливостью и сном уже упомянута в лекции о сне. Но иногда работа рубца обнаруживается и иначи, без раздракения двигательной области полушарий, ограничиваясь раздражением других анализаторов. У собаки были вырезаны лобные доли (опыты Бабкина). Собака после операции быстро оправилась. Но около 2 месяцев спустя на 10 дней у нее появилась чрезвычайная гиперестезия кожи. Она кричала при малейшем прикосновении, кричала при движении и при этом падала, вся съеживаясь. Очевидно, рубец в кожном анализаторе раздражал отдел рецепторных разрушительных (субъективно говоря, болевых) клаток, если таковые отдельно имеются. Еще интереснее и отчетливее случай другой собакой (опыты Ерофеевой), у которой был частично экстирпирован кожный анализатор. У нее через месяца после операции разразился сильнейший судорожный припадок. Во время припадка она была вновь оперирована. Рубец, давший большие выросты дальше места первоначального оперирования, был возможно тщательно удален. Судороги после того больше не повторялись. Но наступили другие припадки, повторявшиеся и каждый раз продолжавшиеся по нескольку дней. Когда ее хозяин-экспериментатор или еда попадали в поле зрения левого глаза (она была оперирована справа), она быстро отвертывалась, а если была свободна, убегала в видимом особенном возбуждении. С правой же стороны ее реакция на то и другое в то же время оставалась совершенно нормальной. Иногда она сама по себе на свободе, озираясь в левую сторону, быстро вскакивала и бежала куда ни попало. Все это легко понимается, если мы примем, что остатки рубца, действовавшие в области оптического анализатора и притом одной стороны, внутренним раздражением прибавляли нечто к внешним раздражениям глаза, придавая-таким образом внешним предметам необычайный, чрезвычайный вид, на что и наступала реакция животного как и на всякий реальный, но чрезвычайный предмет. Короче говоря - рубцом обусловливалась иллюзия. Очевидно, то же самое было и у собаки, упомянутой выше, которая после судорожного припадка также убегала от знакомых людей и подаваемой пищи, приходя в особенное возбуждение. Нужно думать, взрыв возбуждения, вызванный рубцом, закончившись в двигательной области коры, последовательно задержался на некоторое время в оптическом анализаторе. Можно с правом сказать, что в приведенных случаях мы имеем дело с так называемыми эпилептическими эквивалентами. Эти факты подали нам мысль исследовать раздражением все анализаторы. Вживляя надолго электроды на разных пунктах поверхности полушарий, мы рассчитываем на собаках с различными условными раздражителями посредством электрического тока видоизменять эффект то тех, то других условных раздражителей. Методическая часть выработана. Предстоят опыты. Пр Приходится жалеть, что в период (более ранний в нашей работе) большинства опытов с экстирпациями мы еще не располагали сведениями ни о разных типах нервной системы наших экспериментальных животных, ни о патологическом состоянии полушарий под влиянием функциональных воздействий на них. Вероятно, при этих сведениях утилизация материала с экстирпациями была бы и шире и глубже. Теперь, после беглого обзора общих последствий операций над большими полушариями, я перехожу к возможному использованию методом условных рефлексов опытов с экстирпациями для цели определения физиологического значения как всей массы полушарий, так и больших или меньших частей их. Мы, как и прежде всех Гольц, лишали наше экспериментальное животное совсем больших полушарий, но с тем специально, чтобы установить отношение их к условнорефлекторной деятельности животного, как мы ее изучили (опыта Зеленого). Так как общее поведение собак после удаления полушарий многократно и подробно описано самим Зеленым, то я коснусь здесь исключительно вопроса о связи наших условных рефлексов с большими полушариями. Обычные условные рефлексы, очевидно, совершенно отсутствовали у собак без полушарий, несмотря на наше настойчивое воспроизведение условий, при которых они должны были проявиться. Поэтому мы сосредоточили особенное внимание на одном из рефлексов, оказывавшемся по другим нашим опытам в высшей степени устойчивым, в отличие от всех остальных. Это так называемый нами водяной условный рефлекс, рефлекс с воспринимающей поверхности рта. Если собаке приборчиком, помещенным во рту, вливать кислоту, то после немногих таких вливаний и вливание одной воды, которая обыкновенно не влечет за собой сколько-нибудь значительного слюноотделения (самое большее --- 1--2 капли), теперь обильно гонит слюну. Следовательно, какое-то раздражение водой рецепторного аппарата полости рта, совпадая во времени с действием кислоты, приобретает условное кислотноее действие, чтобнаруживается в большом количестве слюны и в специфической двигательной реакции. Этот водяной рефлекс, как увидим позднее при другом случае, обладает всеми свойствами других условных рефлексов, так что не может быть сомнений в его условной натуре. У собаки без полушарий, которая у нас жила всего больше, водяной условный рефлекс был выработан перед окончательным удалением всей массы полушарий и достиг размера 8-10 капель на 5 куб. см воды. После полного удаления полушарий, начиная с шестого дня, собаке почти ежедневно вливалось пс 5 куб. см 0.25 раствора соляной кислоты по нескольку раз, всего свыше 500 раз. Только 7 месяцев спустя на воду начал появляться рефлекс, который, постепенно усиливаясь, поднялся до 13 капель на 5 куб. см воды. Но что это такое? Этот водяной рефлекс во многих отношениях резко отличался от условного водяного рефлекса. Существенное его отличие состояло в том, что он не угашался, как это отчетливо происходит с настоящим условным водяным рефлексом нормальных животных, когда вода, влитая несколько раз одна без кислоты, скоро теряет свое слюногонное действие. У собаки без полушарий она получила постоянное действие. Наблюдение собаки после вливания воды открыло истинный смысл дела. После вливания воды нее наступали характерные движения, которые обычно появлялись, когда она была голодна; собака начинала ходить туда и сюда наклоненной вниз головой и подергивающимися ноздрями, как бы чего-то ищущая. Очевидно, соприкосновение воды со слизистой оболочкой рта вызывало теперь у собаки сильный безусловный пищевой рефлекс, а это совпадает с тем, что безусловные рефлексы головного мозга, например слюнные, сначала после операции удаления больших полушарий сильно задержанные, затем постепенно восстановляются и, наконец, весьма значительно усиливаются против нормы. Таким образом, не претендуя на абсолютную точность положения, надо принимать, что большие полушария являются главнейшим органом условных рефлексов, что они обладают синтетической функцией в таком объеме и совершенстве, как никакая другая часть центральной нервной системы. Из отдельных анализаторов мы всего более занимались звуковым, с которого я и начну. На трех собаках после частичной операции над большими полушариями мы наблюдали абсолютную глухоту. У двух из них (опыты Кудрина) она наступила после удаления всей задней половины обоих полушарий по линии, начинающейся вверху прямо позади g. sygmoideus и идущей к вершине g. sylviaticus, а затем по fissura fossae Sylvii. Глухота наступила сейчас же после второй операции (удаление происходило в два приема, сперва на одной стороне, потом на другой). Одна собака жила после второй операмесяцев, другая 7. У третьей собаки (опыты Маковского) были удалены с обеих сторон gg. sylviatici poster. , ectosylv. poster. и suprasylv. poster. на одной стороне с захватом среднего и даже переднего отдела этих извилин. Через 1//2 месяца после операции у нее сразу наступила абсолютная глухота. Накануне было ослабление тормозного процесса. Нужно думать, что ночью были судороги. После этого собака, повидимому здоровая, жила еще месяц. За это время у нее были выработаны новые рефлексы на агенты, относящиеся к кожно-механическому, запаховому и световому анализаторам. Погибла при судорожном припадке. Многие другие собаки, оперированные совершенно так же, реагировали на звук, хотя жили после операции не меньше, а некоторые даже больше тех собак. Как понимать эту совершенную потерю звуковой реакции? Раз остается точным фактом, что после полного удаления больших полушарий собака все же резко реагирует на звук, необходимо признать в наших трех случаях или какое-либо повреждение подкорковых узлов, или задерживание, простирающееся до них. Последнее не должно быть исключено потому, во-первых, что при исследовании (к сожалению, только макроскопическом) не было замечено никакого повреждения подкорковых узлов, во-вторых, потому, что при операции, как у первых двух собак, световая реакция, сперва совершенно исчезавшая, через 2 месяца восстанавливалась, и даже световой условный рефлекс достиг дооперационной высоты, и, наконец, в-третьих, потому, что мы знаем, как импульсы, исходящие из больших полушарий, тормозят рефлексы низших инстанций. Если признать задерживающее действие у третьей собаки, то нельзя не обратить внимание на то, что это задерживание распространилось очень изолированно по системе звукового анализатора, совершенно не затронув других анализаторов. Как правило, после операции удаления височных долей или всей задней половины обоих полушарий общая звуковая функция (поднимание ушей и поворот головы) восстанавливается самое крайнее через немногие дни, обыкновенно через часы, иногда даже вообще не улавливается время исчезания. Это есть, очевидно, безусловный ориентировочный, исследовательский рефлекс, принадлежащий подкорковым узлам, так как он остается навсегда у собак без больших полушарий. Весь остальной объем звуковой функции должен быть отнесен насчет больших полушарий. Эта остальная деятельность звукового анализатора, свойственная только полушариям, сначала после указанных операций вся отсутствует, но потом постепенно, то более, то менее быстро возвращается, никогда, однако, не восстанавливаясь полностью. Когда по удалении височных долей с обеих сторон восстановился исследовательский рефлекс и уже появились условные рефлексы на агенты из других анализаторов, никаких звуковых условных рефлексов не оказывается. Такое положение дела может продолжаться от многих дней до многих недель, в связи с большей или меньшей полнотой данной операции. Кроме того, существенно производство операции или одновременное на обеих сторонах, или по крайней мере через короткий промежуток сперва на одной. потом на другой стороне. В противном случае, т. е. при длинном промежутке, этой фазы можно не заметить. Что значит это отсутствие условных звуковых рефлексов? Возможен ряд предположений: может быть, оставшиеся клетки звукового анализатора по той или другой причине (может быть, ослабленные операцией, или очень уменьшенные в числе, или какие-нибудь запасные, ранее неработавшие, клетки) способны теперь к процессу раздражения и под влиянием внешних раздражений прямо переходят в тормозное состояние. Это одно. Другое состоит в следующем. Может быть, после операции на корковом конце звукового анализатора так понизилась возможность анализаторной функции, что все падающие на животное звуки (а их в обыкновенных лабораториях так много) стали как бы одинаковыми, и потому наш безусловный раздражитель теперь больше чаще не совпадает, чем совпадает cбщенным звуком, и потому условное значение нашего звука должно угаснуть, исчезнуть. Наконец, может быть, под влиянием операции как-то ослабела и временно исчезла сама синтетическая деятельность в звуковом анализаторе. Мы испыервые два предположения специальными опытами (опыты И. И. Крыжановского). Прежде всего звук был введен в условную связь в виде задерживающего агента, в виде условного тормоза. У собаки на условном пищевом раздражителе из запаха камфоры были образованы два условных тормоза: кожно-механический и звуковой (ре духового камертона, 288 колебаний в секунду). Через 3 дня после удаления обеих височных долей положительный камфорный рефлекс был налицо. Условные тормоза несколько дней оставались почти без действия. Но на двенадцатый день они тормозили вполне хорошо. Так же хорошо тормозили и все другие разнообразные, нарочито испробованные звуки. Положительных звуковых условных рефлексов это время не было. Ни на плеск кислоты, вливаемой в рот, ни на хруст сухарей, даваемых собаке есть, рефлексов не образовалось, что в норме происходит чрезвычайно быстро. Что звук действовал в качестве условного тормоза, а не как агент внешнего торможения, было доказано разрушением условно-тормозного действия посредством сопровождения тормозной комбинации кормлением животного, а затем восстановление ее по отмене кормления при этой комбинации. Для контроля совершенно так же разрушался и восстанавливался и кожно-механический условный тормоз. Следовательно, звук был настоящим условным тормозным раздражителем. Еще через несколько дней после этих опытов начали появляться и положительные условные рефлексы. В настоящее время для проверки первого предположения мы применили бы и другие вариации опыта. Для испытания второго предположения, именно, имея в виду чрезвычайно обобщенный звук, мы воспользовались следовым условным рефлексом, образованным на поздних следах, при которых положительный условный раздражитель также имеет и даже еще более обобщенный характер. У собаки имелся следовой рефлекс на кожно-механическое раздражение кожи. Промежуток между концом условного раздражителя и началом безусловного составлял 2 минуты. Слюноотделение обычно начиналось во вторую минуту паузы. Только через 10 дней после полного удаления височных долей появился следовой рефлекс на механическое раздражение кожи. На двенадцатый день испробованное звуковое раздражение дало за 4 минуты всего 8 капель, отделение началось в третью минуту по прекращении раздражения. Но еще через 5 дней тот же звук дал уже 38 капель за 6 минут, причем, как обычно, во вторую минуту по прекращении раздражения. Лишь спустя еще 18 дней начали появляться положительные условные рефлексы на наличные звуки. После этих опытов испытание третьего предположения естьственно отпадало. Две описанные формы опытов, вероятно, относились к двум разным фазам состояния звукового анализатора после операции над ним, первая - к ранней, вторая - к более поздней, тем более, что на других собаках после той же операции мы наблюдали обобщенный характер звуковых раздражений, но с положительным эффектом их и на наличных условных рефлексах (опыты Бабкина). Вот один из этих опытов. У собаки был выработан условный пищевой рефлекс, между прочим, на нисходящий ряд соседних тонов, от которого был хорошо отдифференцирован восходящий ряд тех же тонов. На восьмой день после удаления височных долей опыт шел следующим образом. Мы видим, что, кроме выработанного условного звукового раздражителя, положительно действовало много и посторонних звуков, а некоторые даже одинаково с ним. Когда посторонние звуки как неподкрепляемые ослабевали в их действии, то после них сильно терял в своем действии и выработанный раздражитель. При подкреплении его восстановлялось действие и посторонних звуковых раздражителей, т. е. звук в качестве условного раздражителя стал очень обобщенализ звуков очень понизился, почти исчез. Когда анализ начинает возвращаться к норме, это очень часто происходит очень медленно. Сперва от тоновых раздражителей отпадают звуки другого рода, удары и шумы, но потом долго остается несовершенным само дифференцирование тонов. Например в тех же опытах (Бабкина) прежняя, до операции существовавшая, дифференцировка тонов через постепенные этапы вернулась только 2 месяца спустя после операции.
Описанный факт отсутствия или понижения анализаторной функции звукового нервного аппарата есть, очевидно, то же самое, что описано впервые Мунком под названием психической глухоты. Нельзя не видеть существенной разницы в понимании факта с психологической и чисто физиологической точки зрения. При формуле Мунка «животное слышит, но не понимает» исследование упираетсB термин «понимает», как в угол. Что же дальше? С физиологической же точки открывается обширное поле исследования этапов восстановления нарушенной функции звукового анализатора. В норме звуки дифференцируются, во-первых, по общим свойствам: силе, продолжительности, сплошному или прерывистому течению и по месту, и, во-вторых, по их чисто звуковым свойствам: удары, тоны и г. д. Нужно было ожидать ( и в этом отношении, как только что упомянуто, у нас уже имеются положительное данные), что при восстановлении нарушенного звукового анализатора должны проходить перед экспериментатором разные стадии деятельности его, з изучение их дает основание для более полного понимания механизма звукового анализа.
Но сказанным не исчерпывается нарушение деятельности звукового анализатора при удалении височных долей. Остается еще - и, очевидно, главнейший - дефект ее, выдвигается еще важный этап звукового анализа. Уже давно было замечено и подтверждено многими авторами, что после этой операции собаки перестают отзываться на кличку. То же видели и мы. Что значит это? Надо было думать, что здесь идет дело об исчезании специально анализа комплексных звуковых раздражений. Ради выяснения этого пункта нами были поставлены нарочитые опыты (опыты Бабкина). Условные раздражители образовывались из последовательных рядов одних и тех же тонов, но различным образом расположенных в группе: или с различной последовательностью тонов по высоте, илc разными паузами между тонами на разных местах группы. Из одного расположения тонов делался положительный условный раздражитель, из другого отрицательный (дифференцировка). Эти дифференцировки, как сообщено в ранних лекциях, давались звуковому анализатору труднее, чем дифференцировки отдельных тонов. Кроме дифференцировок комплексных раздражителей, данных собак имелись дифференцировки и отдельных тонов. Затем у собак удалялись полностью височные доли полушарий. У пятг: собак, служивших для этих опытов, получилсясовершенно однообразный результат. В то время как дифференцировка отдельных тонов в ранее, до операции, испытанных пределах (до одного тона) рано или поздно восстанавливалась, ни малейшего намека на возвращение дифференцировок комплексных раздражений увидеть не удалось, хотя большинство собак подвергались опытам до 2-3 месяцев после операции, а одна собака даже спустя почти 3 года после операции. У последней собаки, оставшейся после работы Кудрина, была удалена, как указано выше, вся задняя половина полушарий на линии прямо позади g. sygmoideus до вершины g. sylviaticus и потом до основания мозга по fissura fossae Sylvii. Эта операция была закончена 5 мая 1909 года. Данное исследование было предпринято к концу 1911 года. Я опишу этот опыт. У этой собаки теперь были образованы пищевые рефлексы на ряд восходящих тонов (духовые камертоны 290, 325, 370 и 413 колебаний в секунду) и отдельно на тон тонвариатора Макса Коля в 1200 колебаний в секунду. Рефлексы образовались довольно скоро. Затем была предпринята выработка дифференцировки на тон в 1066 колебаний в секунду (через этапы тонов в 600 и 900 колебаний в секунду). Она была достигнута. Дифференцировка на нисходящий ряд вышеприведенных тонов нимало не обозначилась, несмотря на повторение 400 раз восходящего ряда и 150 нисходящего; точно так же не существовала все 3 года и реакция на кличку.
Вот опыт 15 III 1912.
Мы видим, что, кроме выработанного условного звукового раздражителя, положительно действовало много и посторонних звуков, а некоторые даже одинаково с ним. Когда посторонние звуки как неподкрепляемые ослабевали в их действии, то после них сильно терял в своем действии и выработанный раздражитель. При подкреплении его восстановлялось действие и посторонних звуковых раздражителей, т. е. звук в качестве условного раздражителя стал очень обобщенализ звуков очень понизился, почти исчез. Когда анализ начинает возвращаться к норме, это очень часто происходит очень медленно. Сперва от тоновых раздражителей отпадают звуки другого рода, удары и шумы, но потом долго остается несовершенным само дифференцирование тонов. Например в тех же опытах (Бабкина) прежняя, до операции существовавшая, дифференцировка тонов через постепенные этапы вернулась только 2 месяца спустя после операции.
Описанный факт отсутствия или понижения анализаторной функции звукового нервного аппарата есть, очевидно, то же самое, что описано впервые Мунком под названием психической глухоты. Нельзя не видеть существенной разницы в понимании факта с психологической и чисто физиологической точки зрения. При формуле Мунка «животное слышит, но не понимает» исследование упираетсB термин «понимает», как в угол. Что же дальше? С физиологической же точки открывается обширное поле исследования этапов восстановления нарушенной функции звукового анализатора. В норме звуки дифференцируются, во-первых, по общим свойствам: силе, продолжительности, сплошному или прерывистому течению и по месту, и, во-вторых, по их чисто звуковым свойствам: удары, тоны и г. д. Нужно было ожидать ( и в этом отношении, как только что упомянуто, у нас уже имеются положительное данные), что при восстановлении нарушенного звукового анализатора должны проходить перед экспериментатором разные стадии деятельности его, з изучение их дает основание для более полного понимания механизма звукового анализа.
Но сказанным не исчерпывается нарушение деятельности звукового анализатора при удалении височных долей. Остается еще - и, очевидно, главнейший - дефект ее, выдвигается еще важный этап звукового анализа. Уже давно было замечено и подтверждено многими авторами, что после этой операции собаки перестают отзываться на кличку. То же видели и мы. Что значит это? Надо было думать, что здесь идет дело об исчезании специально анализа комплексных звуковых раздражений. Ради выяснения этого пункта нами были поставлены нарочитые опыты (опыты Бабкина). Условные раздражители образовывались из последовательных рядов одних и тех же тонов, но различным образом расположенных в группе: или с различной последовательностью тонов по высоте, илc разными паузами между тонами на разных местах группы. Из одного расположения тонов делался положительный условный раздражитель, из другого отрицательный (дифференцировка). Эти дифференцировки, как сообщено в ранних лекциях, давались звуковому анализатору труднее, чем дифференцировки отдельных тонов. Кроме дифференцировок комплексных раздражителей, данных собак имелись дифференцировки и отдельных тонов. Затем у собак удалялись полностью височные доли полушарий. У пятг: собак, служивших для этих опытов, получилсясовершенно однообразный результат. В то время как дифференцировка отдельных тонов в ранее, до операции, испытанных пределах (до одного тона) рано или поздно восстанавливалась, ни малейшего намека на возвращение дифференцировок комплексных раздражений увидеть не удалось, хотя большинство собак подвергались опытам до 2-3 месяцев после операции, а одна собака даже спустя почти 3 года после операции. У последней собаки, оставшейся после работы Кудрина, была удалена, как указано выше, вся задняя половина полушарий на линии прямо позади g. sygmoideus до вершины g. sylviaticus и потом до основания мозга по fissura fossae Sylvii. Эта операция была закончена 5 мая 1909 года. Данное исследование было предпринято к концу 1911 года. Я опишу этот опыт. У этой собаки теперь были образованы пищевые рефлексы на ряд восходящих тонов (духовые камертоны 290, 325, 370 и 413 колебаний в секунду) и отдельно на тон тонвариатора Макса Коля в 1200 колебаний в секунду. Рефлексы образовались довольно скоро. Затем была предпринята выработка дифференцировки на тон в 1066 колебаний в секунду (через этапы тонов в 600 и 900 колебаний в секунду). Она была достигнута. Дифференцировка на нисходящий ряд вышеприведенных тонов нимало не обозначилась, несмотря на повторение 400 раз восходящего ряда и 150 нисходящего; точно так же не существовала все 3 года и реакция на кличку.
Вот опыт 15 III 1912.
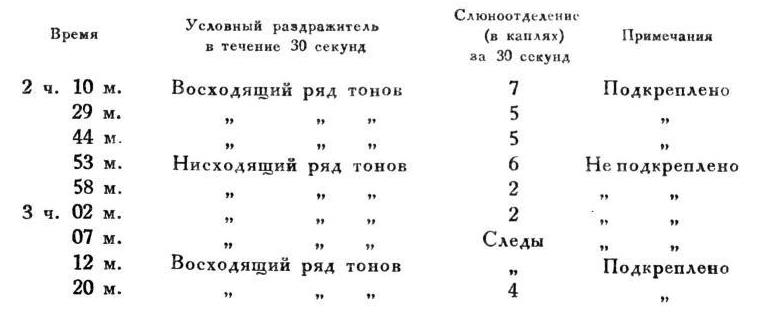 Сюда же, как мне кажется, можно отнести и следующий, хотя и единичный, наш опыт (Эльяссона). У собаки был выработан условный пищевой рефлекс на аккорд из тонов фисгармонии фа1 (F) доз (с') + соль (8") (от 85 до 768 колебаний в секунду). Когда рефлекс достиг максимальной величины, были испробованы отдельные тоны, входящие в аккорд. Все они также действовали, только значительно слабее, чем аккорд, но приблизительно одинаково между собой. Действие промежуточных тонов, соответственно, было еще слабее. После удаления передних половин височных долей отношения существенно изменились: соль4 и соседние с ним тоны потеряли свое действие, хотя рефлекс на аккорд восстановился уже на пятый день после операции. Низкий же тон, входящий в состав аккорда, стал действовать даже с особенной силой, его эффект часто равнялся эффекту всего аккорда. Что значил столь неожиданный и вместе резкий факт? Первое, что приходило в голову, это то, что высокие тоны специально пострадали от операции. Но это оказалось решительно неверным. Когда тон соль4 стали отдельно комбинировать с едой, он очень быстро превратился в хороший условный раздражитель. Мы перешли было к дальнейшим вариациям опыта, но, к большому нашему сожалению, собака скоро погибла. Как понимать этот факт? Свести на влияние разницы силы тонов в аккорде нельзя, так как высокий тон скорее был сильнее, чем низкий и средний. Всего проще принимать, что в слуховом анализаторе существует особый отдел (слуховая сфера Мунка), где специально осуществляются синтез и анализ комплексных звуковых раздражений, как последовательных, так и одновременных. Этот отдел вполне бы аналогировал с бесспорно доказанной так называемой проекцией сетчатки, особым отделом в коре оптического анализатора. По этому представлению надо было бы допустить, что в этом особом отделе звукового анализатора в коре имеются рецепторные клетки с приводами от всех частей периферического звукового аппарата, причем между клетками, благодаря здесь особенно благоприятной конструкции, могут происходить разнообразные и тонкие связи, образовываться наиболее сложные комплексы звуковых раздражений и осуществляться их анализы. Частичное разрушение этого отдела должно вести к выпадению из комплексов отдельных раздражений, а полное должно исключать вообще высший синтез и анализ раздражений. Так как по удалении височных долей полностью звуковые условные рефлексы продолжают существовать (Калишер и мы) и даже способны к элементарному дифференцированию, а при удалении всей коры полушарий они исчезают и навсегда, - то является неизбежным заключение, что, кроме специального отдела звукового анализатора, в коре должны быть рассеянные элементы этого анализатора в гораздо большем районе полушарий, может быть даже во всей массе их. Эти элементы по их конструкции уже более неспособны кложным соединениям, в них возможны только более элементарные синтез и анализ. Не исключается возможность, что это упрощение, ограничение деятельности звукового анализатора, идет тем дальше, чем его отдельные элементы более далеко отстоят от коркового, так сказать, ядра этого анализатора.
Какое представление о конструкции мозгового конца звукового анализатора, как, вероятно, и всякого другого, мне кажется, всего легче, с одной стороны, охватило бы весь наличный фактический материал, с другой - открывало бы огромнейший горизонт для дальнейших исследований. Оно хорошо отвечало бы широкому распространению каждого анализатора, как увидим дальше, между другими анализаторами, далеко за пределы принимаемых ранее локализаций, и делало бы понятным существование особых ядер анализаторов, где благодаря густому и исключительному сконцентрированию элементов данного анализатора делалась бы возможной высшая синтетическая и анализаторная деятельность его. Оно же без затруднения позволяло бы уяснить себе и механизм постепенного усовершенствования под влиянием практики функции остатков анализатора, сначала, после операций над ядром анализатора, очень ограниченной, а затем и предел этого усовершенствования. У последней собаки, описанной выше, элементарный анализ тонов, вероятно, достиг высшей степени, а потеря высшего синтеза и анализа оказалась невозвратемой и после трехгодичной практики.
Предлагаемое представление о распределении анализаторов в больших полушариях естественно ведет, ради фактической проверки его, к длиннейшему ряду экспериментальных задач. Если говорить о звуковом анализаторе, то до частичных операций должны быть применены многочисленные положительные как элементарные, так и комплексные раздражители, а также разнообразные дифференцировки. Рядом с этим должны быть обследованы общие свойства элементарных раздражителей: возбудимость, условия перехода положительного раздражителя в тормозной, подвижность тормозного процесса и т. д., чтобы после операции можно было с большей или меньшей вероятностью определить: что произошло насчет изменения этих общих свойств и что есть специально результат разрушения некоторых конструкций. При исследовании восстановляютейся звуковой функции после операции нужно, как уже сказано выше, возможно тщательно проследить этапы этого восстановления. Конечно, для осуществления всего этого прежде всего необходимо обеспечить продолжительное и вполне здоровое существование животного после мозговой операции, что, к сожалению, теперь большей частью есть только pium desiderium.
Сюда же, как мне кажется, можно отнести и следующий, хотя и единичный, наш опыт (Эльяссона). У собаки был выработан условный пищевой рефлекс на аккорд из тонов фисгармонии фа1 (F) доз (с') + соль (8") (от 85 до 768 колебаний в секунду). Когда рефлекс достиг максимальной величины, были испробованы отдельные тоны, входящие в аккорд. Все они также действовали, только значительно слабее, чем аккорд, но приблизительно одинаково между собой. Действие промежуточных тонов, соответственно, было еще слабее. После удаления передних половин височных долей отношения существенно изменились: соль4 и соседние с ним тоны потеряли свое действие, хотя рефлекс на аккорд восстановился уже на пятый день после операции. Низкий же тон, входящий в состав аккорда, стал действовать даже с особенной силой, его эффект часто равнялся эффекту всего аккорда. Что значил столь неожиданный и вместе резкий факт? Первое, что приходило в голову, это то, что высокие тоны специально пострадали от операции. Но это оказалось решительно неверным. Когда тон соль4 стали отдельно комбинировать с едой, он очень быстро превратился в хороший условный раздражитель. Мы перешли было к дальнейшим вариациям опыта, но, к большому нашему сожалению, собака скоро погибла. Как понимать этот факт? Свести на влияние разницы силы тонов в аккорде нельзя, так как высокий тон скорее был сильнее, чем низкий и средний. Всего проще принимать, что в слуховом анализаторе существует особый отдел (слуховая сфера Мунка), где специально осуществляются синтез и анализ комплексных звуковых раздражений, как последовательных, так и одновременных. Этот отдел вполне бы аналогировал с бесспорно доказанной так называемой проекцией сетчатки, особым отделом в коре оптического анализатора. По этому представлению надо было бы допустить, что в этом особом отделе звукового анализатора в коре имеются рецепторные клетки с приводами от всех частей периферического звукового аппарата, причем между клетками, благодаря здесь особенно благоприятной конструкции, могут происходить разнообразные и тонкие связи, образовываться наиболее сложные комплексы звуковых раздражений и осуществляться их анализы. Частичное разрушение этого отдела должно вести к выпадению из комплексов отдельных раздражений, а полное должно исключать вообще высший синтез и анализ раздражений. Так как по удалении височных долей полностью звуковые условные рефлексы продолжают существовать (Калишер и мы) и даже способны к элементарному дифференцированию, а при удалении всей коры полушарий они исчезают и навсегда, - то является неизбежным заключение, что, кроме специального отдела звукового анализатора, в коре должны быть рассеянные элементы этого анализатора в гораздо большем районе полушарий, может быть даже во всей массе их. Эти элементы по их конструкции уже более неспособны кложным соединениям, в них возможны только более элементарные синтез и анализ. Не исключается возможность, что это упрощение, ограничение деятельности звукового анализатора, идет тем дальше, чем его отдельные элементы более далеко отстоят от коркового, так сказать, ядра этого анализатора.
Какое представление о конструкции мозгового конца звукового анализатора, как, вероятно, и всякого другого, мне кажется, всего легче, с одной стороны, охватило бы весь наличный фактический материал, с другой - открывало бы огромнейший горизонт для дальнейших исследований. Оно хорошо отвечало бы широкому распространению каждого анализатора, как увидим дальше, между другими анализаторами, далеко за пределы принимаемых ранее локализаций, и делало бы понятным существование особых ядер анализаторов, где благодаря густому и исключительному сконцентрированию элементов данного анализатора делалась бы возможной высшая синтетическая и анализаторная деятельность его. Оно же без затруднения позволяло бы уяснить себе и механизм постепенного усовершенствования под влиянием практики функции остатков анализатора, сначала, после операций над ядром анализатора, очень ограниченной, а затем и предел этого усовершенствования. У последней собаки, описанной выше, элементарный анализ тонов, вероятно, достиг высшей степени, а потеря высшего синтеза и анализа оказалась невозвратемой и после трехгодичной практики.
Предлагаемое представление о распределении анализаторов в больших полушариях естественно ведет, ради фактической проверки его, к длиннейшему ряду экспериментальных задач. Если говорить о звуковом анализаторе, то до частичных операций должны быть применены многочисленные положительные как элементарные, так и комплексные раздражители, а также разнообразные дифференцировки. Рядом с этим должны быть обследованы общие свойства элементарных раздражителей: возбудимость, условия перехода положительного раздражителя в тормозной, подвижность тормозного процесса и т. д., чтобы после операции можно было с большей или меньшей вероятностью определить: что произошло насчет изменения этих общих свойств и что есть специально результат разрушения некоторых конструкций. При исследовании восстановляютейся звуковой функции после операции нужно, как уже сказано выше, возможно тщательно проследить этапы этого восстановления. Конечно, для осуществления всего этого прежде всего необходимо обеспечить продолжительное и вполне здоровое существование животного после мозговой операции, что, к сожалению, теперь большей частью есть только pium desiderium.
Лекция двадцатая. Патологические состояния больших полушай как результат оперативных воздействий на них: нарушения деятельности светового анализатора; г) нарушения деятельности кожно-механического анализатора; д) нарушения после удаления лобных долей; e) нарушения кожно-температурного анализатора; ж) нарушения после удаления g. pyriformis; з) нарушения двигательного анализатора
Мм. гг.! Перехожу к деятельности светового анализатора. Хотя деятельность этого анализатора обследовала у нас сравнительно меньше, однако имеющийся физиологии материал позволяет усмотреть очень сходственные отношения его деятельности с деятельностью звукового анализатора в случае удаления соответственных отделов больших полушарий. Опять уже из опытов Гольца над его собакой без больших полушарий следовало, что ориентировочная, самая общая, двигательная реакция на свет принадлежит и подкорковым узлам. Но и только. Указаний других авторов на существование у собак без больших полушарий каких-либо других зрительных функций не имеется. На нашей собаке (Зеленого) даже и упомянутая элементарная реакция не выступала отчетливо. Значит, у нас есть основание говорить о всем остальном объеме зрительной деятельности нашего животного как о свойственном исключительно большим полушариям, подобно деятельности ротового и звукового анализаторов, относительно которых такое заключение экспериментально достаточно обосновано. Если даже и ротовой анализатор, несмотря на его сохранившуюся и без полушарий анализаторную функцию (наша собака без больших полушарий отвергала многие несъедобные вещества), был не в состоянии вступать во временные, условные связи, то тем более не может быть речи об условных рефлексах на световые раздражения через посредство подкорковых узлов. Прежде всего мы, как и все ранние исследователи (до М. А. Минковского включительно) больших полушарий, видели (и это должно считаться точно установленным фактом), что при известных нарушениях затылочных долей полушарий на одной стороне или на обеих происходит соответственно или на одном глазу, или на обоих совершенно точное ограничение поля зрения то с боков, то сверху, то снизу. Таким образом предметы, попадающие в уцелевшие части поля зрения, вызывают у животного соответствующие действия, и они же, чуть сдвинутые, остаются без обычной реакции со стороны животного. Из этого факта необходимо следует, что в затылочных долях имеется ядро зрительного анализатора, делающее возможным существование самых сложных комплексных зрительных раздражений и, следовательно, сложнейшего анализа их. На всех наших, довольно многочисленных, собаках после удаления их затылочных долей на обоих сторонах, как ни долго они жили после операции (одна 3 года), ни разу не было обнаружено предметного зрения. Ни люди, ни другие животные, ни пища не различались оптическим анализатором таких собак. Мы много раз разбрасывали на полу куски пищи или вешали их на нитках на разной высоте, и ни одна из собак без затылочных долей, как они ни были голодны, ни разу не направила к ним своих движений в силу зрительных раздражений, исключительно ориентируясь относительно них при помощи запаховых и кожномеханических раздражений. Уже из факта различно ограниченного поля зрения при различных нарушениях затылочных долей следует, что при потере предметного зрения сущность дела в отсутствии высшего синтеза и анализа именно световых раздражений, не в нарушении дополнительных актов глазной деятельности, как аккомодация и адаптация. Наши собаки без затылочных долей одинаково не различали предметов, как больших, так и малых, как на далеком, так и на близком расстоянии, как при сильном, так и при среднем и слабом освещении. Итак, центральная часть, ядро оптического анализатора, орган высшего синтеза и анализа световых раздражений, находится в затылочных долях полушарий. Но здесь не весь оптический анализатор. Он распространен гораздо шире, может быть, даже по всей массе больших полушарий. Еще со славной эпохи в физиологии больших полушарий (с семидесятых годов) несколькими исследователями утверждалось положение, что и передние доли полушарий имеют отношение к зрению, но отрицательный характер факта не позволял видеть в нем достаточное доказательство. Было возможно все свести на отдаленное задерживающее действие. Мы можем, однако, тепривести положительные факты, что оптический анализатор, и даже еще с значительной функцией, действительно существует и передней половине больших полушарий, отграниченнией, начинающейся наверху прямо позади g. sygmoideus, идущей косо кзади к переднему углу g. sylviaticus и затем до основания полушарий по fissura fossae Sylvii. У всех наших собак без затылочных долей (как и Калишера) легко образовывались условные рефлексы на общее освещение комнаты, в которой находилась собака, причем выступало отчетливо дифференцирование довольно тонких интенсивностей этого освещения. Этот факт дает простое и чисто естественно-научное объяснение тому, что было названо Мунком психической слепотой. После удаления затылочных долей значительно поврежденный оптический анализатор мог войти в условную связь только этой элементарной функцией, именно реакцией на колебания интенсивности световых раздражений. Благодаря этому собака в освещенном пространстве могла обходить затененные предметы и уходить в открытую дверь, как в светлое место. Ввиду этих фактов, если говорить психологически, правильнее было бы сказать, что собака именно понимала, но плохо, недостаточно видела. Но, конечно, такая фомулировка совершенно излишня. Ясно, что существо дела заключается в ограничении анализаторной функции. И дальнейшее изучение фактов вполне подтвердило научную плодотворность такой чисто объективной точки зрения на предной нашей собаки, располагавшей только передними долями полушарий по вышеуказанной линии, можно было образовать условный рефлекс и на более высшую функцию светового анализатора. Это та же собака, которая только что была упомянута как прожившая после операции 3 года, а также была приведена в конце прошлой лекции как пример потери навсегда анализа комплексных звуковых раздражений. Я опишу подробно все наши опыты на ней со световыми раздражениями (опыты Кудрина). Операция на обоих полушариях была произведена с промежутками в месяц. Вторая операция была исполнена 5 мая 1909 года. До операции выработанный пищевой рефлекс на вспыхивание в затемненной комнате лампы в 100 свечей после второй операции на пятый день восстановился отчетливо, а к одиннадцатому достиг и даже несколько превзошел дооперационный размер. После того этот рефлекс был оставлен, исследование продолжалось с звуковыми раздражениями. 7 сентября того же года начали образовывать рефлекс на движение освещенной фигуры креста (отбрасываемой проекционным аппаратом) на экране затемненной комнате. Ко Конечно, рефлекс скоро появился и через неделю был значительным. C 28 сентября отбрасываемую фигуру креста оставили неподвижной. Рефлекс продолжал существовать, несколько уменьшившись в размере. Теперь была предпринята дифференцировка фигуры круга от фигуры креста той же величины площади и того же освещения, т. е. появление креста сопровождалось кормлением, круга - нет. С седьмого раза применения круга начало обнаруживаться его дифференцирование, но опыты были прерваны и возобновились лишь полгода спустя. Рефлекс на фигуру креста уцелел. Дифференцировка быстро появилась и скоро сделалась полной. Вот конечный результат этих опытов.
 Вскрытие собаки 3 года спустя после мозговых операций вполне подтвердило полноту операций: от задней половины полушарий не осталось ни малейшего следа.
Не может подлежать сомнению факт, что у данной собаки оставшаяся в передней половине полушарий часть светового анализатора была способна образовывать условные раздражители не только из колебании интенсивности освещения, но и из различных форм освещенных и затемненных предметов. А вместе с тем, как уже было сказано, у этой собаки, как и у всех других, при удалении затылочных долей до конца жизни не наблюдалось условных рефлексов на отдельные предметы. Имея в виду, что у данной собаки впервые испытанное образование условных рефлексов на разные формы удалось уже спустя только 4 месяца после операции, значит, вероятно, могло произойти и раньше, а затем спустя еще 2/2 года не появилось предметного зрения, нужно заключить, что состояние светового анализатора к 3 годам после операции было окончательным, бесповоротным. При этом можно было бы обратить внимание еще на следующее. Почему же, раз у нашей собаки наблюдалось различение форм на станке, она не различала предметов по форме на свободе? Но есть огромная разница в обстановке опыта на станке и на свободе среди разных предметов. На свободе масса форм, и притом постоянно более или менее изменяющихся для глаза животного вследствие как движения некоторых предметов, так и движений самого животного относительно всех предметов. К тому же и резкость границ форм также не остается постоянной, завися от колебаний освещения при перемещении некоторых объектов и животного относительно их. В станке при нашем опыте - явно все иначе, все чрезвычайно упрощено сравнительно с условиями обычной действительности. Вероятно, потребовалась бы очень постепенная и долговременная практика, чтобы остающаяся функция различения форм получила жизненное практическое значение для животного.
На основании сообщенного главнейшие этапы нарушенной деятельности светового и звукового анализаторов могут до известной степени считаться соответственными между собой. Минимальное нарушение в световом анализаторе есть ограничение поля нормального зрения, в звуковом, еели доверять нашему единственному опыту, есть выпадение из звуковых комплексов отдельных звуков. Более значительнарушением светового анализатора является отсутствие различения предметов, т. е. комплексов из формы, светотени и окраски (когда цветовая раздражимость встречается у отдельных собак), иначе говоря, потеря высшего синтеза и анализа световых раздражителей. Совершенно таким же нарушеникового анализатора, очевидно, надо считать отсутствие различения комплексов из разных звуков, например слов, значить опять же потерю высшего синтеза и анализа звуковых раздражений. Максимальное нарушение обоих анализаторов, не говоря о полном их разрушении, выражается различением только интенсивности как световых, так и звуковых раздражений. В середине между крайними этапами нарушения стоят случаи, когда вместе c разными интенсивностями в световом анализаторе различаются еще и разные группироветящихся пунктов, а в звуковом - разные роды звуков: удары, шумы и разные тоны.
Дальше перед нами стоял вопрос: не аналогично ли распределение кожно-механического анализатора в массе полушарий описанному распределению в ней светового и звукового анализаторов, т. е. не имеется ли и при этом анализаторе, кроме специального высшей деятельностью района, более широко рассеянных кожно-механических рецепторных элементов с более ограниченными функциями? Не имея еще окончательного ответа на этот вопрос, мы, однако, на основании нашего материала склонны к решению его в положительном смысле. Вот наш старый и текущий, сюда относящийся запас опытов, представляющий некоторый интерес и помимо ответа на вышепоставленный вопрос. Давно уже (опыты Тихомирова) мы видели, что удаление передних частей полушарий ведет за собой исчезание условных кожномеханических рефлексов при сохранении рефлексов на агенты, относящиеся к другим анализаторам. Позднейших опытах (опыты Красногорского) было точно констатировано (согласно установленным данным), что двигательная область более или менее отграничена от кожно-механического анализатора, его специального района и что отдельным участкам кожи соответствуют определенные части этого районa. Привожу пример. У собаки, кроме условных рефлексов на агенты из других анализаторов, имеется генерализованный по всей кожной поверхности кислотный кожно-механический условный рефлекс. Теперь у нее экстирпируются gg. coronarius и ectosylvius с левой стороны. На четвертый день после операции условные рефлексы в области других анализаторов уже налицо. Условный кожно-механический рефлекс появляется на восьмой день, но только на левой стороне, и быстро достигает нормального размера. На десятый он есть и на правой стороне, но лишь на коже середины туловища. На передней и задней конечностях и на плечевом и тазовом поясах туловища он полностью отсутствует, причем граница районов с рефлексами и без них очень резка. Отсутствие рефлексов продолжается до девяностого дня после операции, а затем они начинают восстанавливаться последовательно сверху вниз. При этих «опытах особенный интерес, помимо топографии, совпадающей с банковской, заключается в следующем. В то время как условные рефлексы на указанных местах кожи потеряли свое положительное действие, они обнаруживали резкое тормозящее действие (что указано уже раньше в лекции о сне). Это выражалось в том, что, повидимому, недействительное раздражение этих мест, комбинированное с раздражением других отделов кожи, дающим положительный эффект, и с положительно действующим раздражением агентов из других анализаторов, уменьшало или уничтожало положительный эффект этих последних. Кроме того, повторное раздражение этих мест, и особенно несколько удлиненное, каждый раз вело быстро к развитию сонливости и к полному усыплению животного, которое до этого в станке никогда не только не спало, но и не было сонливым. Сон наступал только при этих раздражениях. Без них в той же обстановке животное оставалось бодрым. В более недавнее время эти опыты были повторены вообще с тем же результатом (опыты Разенкова). У данного животного, пока не применялось раздражение пострадавших от мозговой операции мест кожи (разрушались те же извилины, но частично), рефлексы, относящиеся к другим анализаторам, были. После этого раздражения животное на все остальное время опыта становилось сонливым и все остальные рефлексы делались нулевыми. Являлся интерес исследовать: нельзя ли каким-нибудь способом открыть в этом раздражении элемент положительного действия? Он оказался (опыты того же Разенкова). До сих пор условное раздражение изолированно, т. е. до присоединения безусловного раздражителя, продолжалось 30 секунд. В начале новой вариации опыте изолированное раздражение недействительных мест кожи несколько раз применялось каждый раз только по 5 секунд, конце опыта, как обычно, продолжалось 30 секунд. При этих условиях можно было видеть и положительное действие этого раздражения. Эффект наступал скоро, был небольшим, а главное, еще во время раздражения уже исчезал, когда при других раздражителях он обыкновенно нарастал к концу изолированного раздражения. Вот относящийся сюда опыт.
У собаки положительными условными раздражителями были: удары метронома, свисток, вспыхивание лампочки и механическое раздражение кожи. В данноме раздражался оказавшийся после операции недействительным пункт кожи на предплечье. Отделение слюны отмечалось в делениях трубки; 5 делений которой отвечают 1 капле. Отделение у собаки постоянно небольшое. Судорог у собаки после операции замечено не было.
Вскрытие собаки 3 года спустя после мозговых операций вполне подтвердило полноту операций: от задней половины полушарий не осталось ни малейшего следа.
Не может подлежать сомнению факт, что у данной собаки оставшаяся в передней половине полушарий часть светового анализатора была способна образовывать условные раздражители не только из колебании интенсивности освещения, но и из различных форм освещенных и затемненных предметов. А вместе с тем, как уже было сказано, у этой собаки, как и у всех других, при удалении затылочных долей до конца жизни не наблюдалось условных рефлексов на отдельные предметы. Имея в виду, что у данной собаки впервые испытанное образование условных рефлексов на разные формы удалось уже спустя только 4 месяца после операции, значит, вероятно, могло произойти и раньше, а затем спустя еще 2/2 года не появилось предметного зрения, нужно заключить, что состояние светового анализатора к 3 годам после операции было окончательным, бесповоротным. При этом можно было бы обратить внимание еще на следующее. Почему же, раз у нашей собаки наблюдалось различение форм на станке, она не различала предметов по форме на свободе? Но есть огромная разница в обстановке опыта на станке и на свободе среди разных предметов. На свободе масса форм, и притом постоянно более или менее изменяющихся для глаза животного вследствие как движения некоторых предметов, так и движений самого животного относительно всех предметов. К тому же и резкость границ форм также не остается постоянной, завися от колебаний освещения при перемещении некоторых объектов и животного относительно их. В станке при нашем опыте - явно все иначе, все чрезвычайно упрощено сравнительно с условиями обычной действительности. Вероятно, потребовалась бы очень постепенная и долговременная практика, чтобы остающаяся функция различения форм получила жизненное практическое значение для животного.
На основании сообщенного главнейшие этапы нарушенной деятельности светового и звукового анализаторов могут до известной степени считаться соответственными между собой. Минимальное нарушение в световом анализаторе есть ограничение поля нормального зрения, в звуковом, еели доверять нашему единственному опыту, есть выпадение из звуковых комплексов отдельных звуков. Более значительнарушением светового анализатора является отсутствие различения предметов, т. е. комплексов из формы, светотени и окраски (когда цветовая раздражимость встречается у отдельных собак), иначе говоря, потеря высшего синтеза и анализа световых раздражителей. Совершенно таким же нарушеникового анализатора, очевидно, надо считать отсутствие различения комплексов из разных звуков, например слов, значить опять же потерю высшего синтеза и анализа звуковых раздражений. Максимальное нарушение обоих анализаторов, не говоря о полном их разрушении, выражается различением только интенсивности как световых, так и звуковых раздражений. В середине между крайними этапами нарушения стоят случаи, когда вместе c разными интенсивностями в световом анализаторе различаются еще и разные группироветящихся пунктов, а в звуковом - разные роды звуков: удары, шумы и разные тоны.
Дальше перед нами стоял вопрос: не аналогично ли распределение кожно-механического анализатора в массе полушарий описанному распределению в ней светового и звукового анализаторов, т. е. не имеется ли и при этом анализаторе, кроме специального высшей деятельностью района, более широко рассеянных кожно-механических рецепторных элементов с более ограниченными функциями? Не имея еще окончательного ответа на этот вопрос, мы, однако, на основании нашего материала склонны к решению его в положительном смысле. Вот наш старый и текущий, сюда относящийся запас опытов, представляющий некоторый интерес и помимо ответа на вышепоставленный вопрос. Давно уже (опыты Тихомирова) мы видели, что удаление передних частей полушарий ведет за собой исчезание условных кожномеханических рефлексов при сохранении рефлексов на агенты, относящиеся к другим анализаторам. Позднейших опытах (опыты Красногорского) было точно констатировано (согласно установленным данным), что двигательная область более или менее отграничена от кожно-механического анализатора, его специального района и что отдельным участкам кожи соответствуют определенные части этого районa. Привожу пример. У собаки, кроме условных рефлексов на агенты из других анализаторов, имеется генерализованный по всей кожной поверхности кислотный кожно-механический условный рефлекс. Теперь у нее экстирпируются gg. coronarius и ectosylvius с левой стороны. На четвертый день после операции условные рефлексы в области других анализаторов уже налицо. Условный кожно-механический рефлекс появляется на восьмой день, но только на левой стороне, и быстро достигает нормального размера. На десятый он есть и на правой стороне, но лишь на коже середины туловища. На передней и задней конечностях и на плечевом и тазовом поясах туловища он полностью отсутствует, причем граница районов с рефлексами и без них очень резка. Отсутствие рефлексов продолжается до девяностого дня после операции, а затем они начинают восстанавливаться последовательно сверху вниз. При этих «опытах особенный интерес, помимо топографии, совпадающей с банковской, заключается в следующем. В то время как условные рефлексы на указанных местах кожи потеряли свое положительное действие, они обнаруживали резкое тормозящее действие (что указано уже раньше в лекции о сне). Это выражалось в том, что, повидимому, недействительное раздражение этих мест, комбинированное с раздражением других отделов кожи, дающим положительный эффект, и с положительно действующим раздражением агентов из других анализаторов, уменьшало или уничтожало положительный эффект этих последних. Кроме того, повторное раздражение этих мест, и особенно несколько удлиненное, каждый раз вело быстро к развитию сонливости и к полному усыплению животного, которое до этого в станке никогда не только не спало, но и не было сонливым. Сон наступал только при этих раздражениях. Без них в той же обстановке животное оставалось бодрым. В более недавнее время эти опыты были повторены вообще с тем же результатом (опыты Разенкова). У данного животного, пока не применялось раздражение пострадавших от мозговой операции мест кожи (разрушались те же извилины, но частично), рефлексы, относящиеся к другим анализаторам, были. После этого раздражения животное на все остальное время опыта становилось сонливым и все остальные рефлексы делались нулевыми. Являлся интерес исследовать: нельзя ли каким-нибудь способом открыть в этом раздражении элемент положительного действия? Он оказался (опыты того же Разенкова). До сих пор условное раздражение изолированно, т. е. до присоединения безусловного раздражителя, продолжалось 30 секунд. В начале новой вариации опыте изолированное раздражение недействительных мест кожи несколько раз применялось каждый раз только по 5 секунд, конце опыта, как обычно, продолжалось 30 секунд. При этих условиях можно было видеть и положительное действие этого раздражения. Эффект наступал скоро, был небольшим, а главное, еще во время раздражения уже исчезал, когда при других раздражителях он обыкновенно нарастал к концу изолированного раздражения. Вот относящийся сюда опыт.
У собаки положительными условными раздражителями были: удары метронома, свисток, вспыхивание лампочки и механическое раздражение кожи. В данноме раздражался оказавшийся после операции недействительным пункт кожи на предплечье. Отделение слюны отмечалось в делениях трубки; 5 делений которой отвечают 1 капле. Отделение у собаки постоянно небольшое. Судорог у собаки после операции замечено не было.

 Того же, т. е небольшого положительного действия при раздражении обыкновенно недействительных мест кожи, можно было достигнуть и некоторыми другими приемами: при помощи положительной индукции, посредством растормаживания и впрыскиванием раствора кофеина. Перед нами, очевидно, как уже сказано раньше при другом случае, явление максимальной раздражительной слабости.
Во всех наших опытах до текущих включительно (опыты Федорова) обыкновенно недействительные места с течением времени, рано или поздно, получают снова свои нормальную функцию, как это заявлялось и раньше разными авторами. Перед нами поднимался вопрос, ставившийся и другими исследователями: каким образом, на счет чего происходит это восстановление? Конечно, прежде всего можно было думать о прямых путях, кроме перекрещенных, соединяющих участки кожи с полушариями. Ради этого у собак удалялось одно полушарие целиком и исследовались условные кожно-механические рефлексы с противоположной стороны, пока животные оставались в живых (некоторые из них жили более года и без судорог). Опыты в последнее время пока исполнены на четырех собаках, большей частью с пищевыми условными рефлексами, частью с оборонительным на вливание кислоты и на слабый электрический ток, приложенный к коже на стороне, противоположной операции (опыты Фурсикова и Быкова). Полученный результат оставался во всех случаях отрицательным, несмотря на разные вариации опытов. Для повышения возбудимости применялось впрыскивание стрихнина и кофеина (в опытах Фурсикава), а также испытывалось различным образом возможное тормозящее действие раздражений кожи на поврежденной стороне (в опытах Быкова). Последнее настойчиво применялось ввиду выше сообщенных данных относительно кожного анализатора при частичном нарушении полушария. Тормозящее действие не проявлялось на разных положительных условных рефлексах, и между прочими на кожно-механических здоровой стороны, ни последовательно, ни при одновременном применении. Точно так же не оказалось ни малейшего снотворного действия раздражений кожи на поврежденной стороне. Тем менее, конечно, эти раздражения могли сделаться положительно действующими при употреблении приемов, которыми достигалось активирование, как только что указано выше, недействительных кожно-механических раздражений при частичном нарушении кожного анализатора. Те раздражения были, следовательно, вполне индифферентными в отношении условнорефлекторной деятельности; иначе говоря, по нашим теперешним опытам не оказывалось прямых путей, связывающих кожу половины тела с полушарием той же стороны.
Когда таким образом было исключено участие в восстановлении временно исчезавших рефлексов с прямых (не перекрещенных) путей, оставалась возможность замещения деятельности удаленных частей оставшимися в том же полушарии, как это предполагалось и испытывалось другими исследователями. Ввиду этого прежде всего мы удаляли возможно больше передних извилин одного полушария. Восстановление сначала надолго исчезавших условных кожно-механических рефлексов с течением времени все же происходило. Но и теперь можно было думать -о прилежащих к району первой операции частях. Однако добавочное разрушение этих частей (опыты Юрман) почти совсем не оказало действия на восстановленную деятельность анализатора. Следовательно, возмещение произошло за счет отдаленных элементов. При таком исходе дела для нас, во-первых, обострился вопрос о безупречности нашей методики кожно-механического раздражения, возможно осложняемой звуковым компонентом. Был сконструирован прибор, работа которого, по крайней мере для нашего уха, была беззвучной. Но и этого без контроля было мало. Поэтому мы между кожей и прибором помещали среду, которая исключала механическое раздражение кожи прибором, но не должна была устранять звуковой компонент прибора, если он был. Тогда рефлексов при действии прибора не появлялось. Следовательно, при обычной работеприбора рефлексы возбуждаются действительно воздействием его на кожу, а не на ухо собаки. Во-вторых, по аналогии со световыми и звуковыми анализаторами надо было предполагать, что оставшиеся после первой операции части кожного анализатора владеют только ограниченной функцией сравнительно с первоначально удаленным районом. Мы рассчитывали проверить это опытами с условными комплексными раздражениями. кожи, именно выработав на нормальной коже дифференцировку на разное направление двигающегося механического раздражения кожи и испытывая ее потом на местах, временно лишавшихся условной рефлекторной деятельности после первой операции. Опыты только производятся. Но и все опыты относительно восстановления кожных рефлексов после экстирпации, ввиду разногласия с одной нашей же старой работай, мы продолжаем повторять.
Кроме сообщенного относительно кожно-механического анализатора, мы располагаем еще опытами над связью этих анализаторов обеих сторон между собой. Как упоминалось и раньше, y многих наших соработников (Красногорский, Анреп и др.) резко выступал факт, что условные кожно-механические рефлексы, выработанные на разных местах кожи одной половины тела, удивительно точно воспроизводятся сами собой сразу на симметричных местах кожи другой половины тела. Это одинаково касается как положительных, так и отрицательных условных рефлексов. На основании этого факта надо было ожидать, что дифференцирование симметричных мест кожи обеих половин тела должно быть более или менее трудным процессом, что и оказалось на деле (опыты Быкова, Подкопаева и Григорович). Как происходит этот неожиданный факт? Естественно было думать o комиссуральной связи. И действительно, перерезка corpus. callosum уничтожала это факт. После перерезки условные кожномеханические рефлексы на обеих сторонах стали совершенно независимыми друг от друга (опыты Быкова). Применялисьусловные кожно-механические рефлексы: пищевой и оборонительные на вливание в рот кислоты и на приложенный к коже слабый электрический ток. Ток брался такой силы, чтобы он вызывал только отдергивание ноги и исследовательский рефлекс без сильной реакции в виде борьбы с прибором и криков животного. Опыты очень разнообразились, но результат их всегда был один. Выработанные рефлексы оставались на одной стороне. На другой приходилось вырабатывать их особо. Без затруднений получились противоположные рефлексы на симметричных. местах обеих половин тела.
Приведу пример. У собаки пищевые рефлексы. На правой половине при раздражении бедра - положительный рефлекс, при раздражении плеча - отрицательный, на левой - наоборот.
Того же, т. е небольшого положительного действия при раздражении обыкновенно недействительных мест кожи, можно было достигнуть и некоторыми другими приемами: при помощи положительной индукции, посредством растормаживания и впрыскиванием раствора кофеина. Перед нами, очевидно, как уже сказано раньше при другом случае, явление максимальной раздражительной слабости.
Во всех наших опытах до текущих включительно (опыты Федорова) обыкновенно недействительные места с течением времени, рано или поздно, получают снова свои нормальную функцию, как это заявлялось и раньше разными авторами. Перед нами поднимался вопрос, ставившийся и другими исследователями: каким образом, на счет чего происходит это восстановление? Конечно, прежде всего можно было думать о прямых путях, кроме перекрещенных, соединяющих участки кожи с полушариями. Ради этого у собак удалялось одно полушарие целиком и исследовались условные кожно-механические рефлексы с противоположной стороны, пока животные оставались в живых (некоторые из них жили более года и без судорог). Опыты в последнее время пока исполнены на четырех собаках, большей частью с пищевыми условными рефлексами, частью с оборонительным на вливание кислоты и на слабый электрический ток, приложенный к коже на стороне, противоположной операции (опыты Фурсикова и Быкова). Полученный результат оставался во всех случаях отрицательным, несмотря на разные вариации опытов. Для повышения возбудимости применялось впрыскивание стрихнина и кофеина (в опытах Фурсикава), а также испытывалось различным образом возможное тормозящее действие раздражений кожи на поврежденной стороне (в опытах Быкова). Последнее настойчиво применялось ввиду выше сообщенных данных относительно кожного анализатора при частичном нарушении полушария. Тормозящее действие не проявлялось на разных положительных условных рефлексах, и между прочими на кожно-механических здоровой стороны, ни последовательно, ни при одновременном применении. Точно так же не оказалось ни малейшего снотворного действия раздражений кожи на поврежденной стороне. Тем менее, конечно, эти раздражения могли сделаться положительно действующими при употреблении приемов, которыми достигалось активирование, как только что указано выше, недействительных кожно-механических раздражений при частичном нарушении кожного анализатора. Те раздражения были, следовательно, вполне индифферентными в отношении условнорефлекторной деятельности; иначе говоря, по нашим теперешним опытам не оказывалось прямых путей, связывающих кожу половины тела с полушарием той же стороны.
Когда таким образом было исключено участие в восстановлении временно исчезавших рефлексов с прямых (не перекрещенных) путей, оставалась возможность замещения деятельности удаленных частей оставшимися в том же полушарии, как это предполагалось и испытывалось другими исследователями. Ввиду этого прежде всего мы удаляли возможно больше передних извилин одного полушария. Восстановление сначала надолго исчезавших условных кожно-механических рефлексов с течением времени все же происходило. Но и теперь можно было думать -о прилежащих к району первой операции частях. Однако добавочное разрушение этих частей (опыты Юрман) почти совсем не оказало действия на восстановленную деятельность анализатора. Следовательно, возмещение произошло за счет отдаленных элементов. При таком исходе дела для нас, во-первых, обострился вопрос о безупречности нашей методики кожно-механического раздражения, возможно осложняемой звуковым компонентом. Был сконструирован прибор, работа которого, по крайней мере для нашего уха, была беззвучной. Но и этого без контроля было мало. Поэтому мы между кожей и прибором помещали среду, которая исключала механическое раздражение кожи прибором, но не должна была устранять звуковой компонент прибора, если он был. Тогда рефлексов при действии прибора не появлялось. Следовательно, при обычной работеприбора рефлексы возбуждаются действительно воздействием его на кожу, а не на ухо собаки. Во-вторых, по аналогии со световыми и звуковыми анализаторами надо было предполагать, что оставшиеся после первой операции части кожного анализатора владеют только ограниченной функцией сравнительно с первоначально удаленным районом. Мы рассчитывали проверить это опытами с условными комплексными раздражениями. кожи, именно выработав на нормальной коже дифференцировку на разное направление двигающегося механического раздражения кожи и испытывая ее потом на местах, временно лишавшихся условной рефлекторной деятельности после первой операции. Опыты только производятся. Но и все опыты относительно восстановления кожных рефлексов после экстирпации, ввиду разногласия с одной нашей же старой работай, мы продолжаем повторять.
Кроме сообщенного относительно кожно-механического анализатора, мы располагаем еще опытами над связью этих анализаторов обеих сторон между собой. Как упоминалось и раньше, y многих наших соработников (Красногорский, Анреп и др.) резко выступал факт, что условные кожно-механические рефлексы, выработанные на разных местах кожи одной половины тела, удивительно точно воспроизводятся сами собой сразу на симметричных местах кожи другой половины тела. Это одинаково касается как положительных, так и отрицательных условных рефлексов. На основании этого факта надо было ожидать, что дифференцирование симметричных мест кожи обеих половин тела должно быть более или менее трудным процессом, что и оказалось на деле (опыты Быкова, Подкопаева и Григорович). Как происходит этот неожиданный факт? Естественно было думать o комиссуральной связи. И действительно, перерезка corpus. callosum уничтожала это факт. После перерезки условные кожномеханические рефлексы на обеих сторонах стали совершенно независимыми друг от друга (опыты Быкова). Применялисьусловные кожно-механические рефлексы: пищевой и оборонительные на вливание в рот кислоты и на приложенный к коже слабый электрический ток. Ток брался такой силы, чтобы он вызывал только отдергивание ноги и исследовательский рефлекс без сильной реакции в виде борьбы с прибором и криков животного. Опыты очень разнообразились, но результат их всегда был один. Выработанные рефлексы оставались на одной стороне. На другой приходилось вырабатывать их особо. Без затруднений получились противоположные рефлексы на симметричных. местах обеих половин тела.
Приведу пример. У собаки пищевые рефлексы. На правой половине при раздражении бедра - положительный рефлекс, при раздражении плеча - отрицательный, на левой - наоборот.
 Все четыре рефлекса нужно было вырабатывать отдельно Точно так же всякие посторонние раздражения кожи одной половины тела (тепло в 50°С, слабый электрический ток в случае пищевых и кислотных рефлексов), вызывая исследовательский. рефлекс, тормозили условные кожно-механические рефлексы только той же половины тела, совершенно не влияя на такие же рефлексы другой половины тела. Все эти опыты исполнены на 3 животных.
Рядом с специальными опытами над кожным анализатором могут быть упомянуты наши опыты (опыты Бабкина) с удалением только лобных долей. Удалялись лобные доли с обеих сторон по sulcus praecruciatus и sulcus praesylvius до основания мозга, следовательно, с разрушением lobi olfactorii. Иногда нож при операции и почти постоянно последующий патологический процесс, как показывало потом вскрытие, захватывали и дальнейшие кзади лежащие извилины. ы произведены на. 4 собаках. Во всех случаях условные рефлексы с глаза и уха после операции восстанавливаются или образуются более или менее быстро. Нарушения, и значительные, были только со стороны кожного анализатора и движения. Большей частью нельзя было образовать, пока животные жили (от одного месяца до шести, умирая во время судорог), условных положительных кожно-механических рефлексов специально c туловища. C конечностей иногда вырабатывались. Отрицательные же рефлексы в виде условного тормоза могли быть получены со всех мест кожи. Рядом с этим почти постоянно существовала общая повышенная возбудимость кожной поверхности, так что некоторые собаки не выносили петель на ногах на станке во время опыта и стояли на станке спокойно только без них. Также наблюдались и временные двигательные расстройства в виде измененной позы животных (опущенная вниз голова и выгнутая кверху спина) и паретических явлений в конечностях вместе с подергиваниями. Особенно были сильны расстройства в движениях рта. В ближайшее время после операции животные с трудом захватывали в особенности твердую пищу, так что приходилось кормить их с руки. Обшее же поведение животного, за исключением перечисленных частностей, не представляло ничего особенного.
Из небольшого числа наших опытов словными рефлексами на агенты кожно-температурного анализатора, повидимаму, следует, что этот анализатор и кожио-механический топографически в коре не совсем совпадают. При удалении g. praecruciatus (опыты Шишло), в то время как условный кожномеханический рефлекс на задней конечности восстановился быстро (около недели), температурный (нлод и на температуру 47.5° C) отстал в его появлении очень резко, на 4 недели.
При помощи метода условных рефлексов мы рассчитывали получить данные относительно указываемых некоторыми авторами отношений g. pyriformis к запаховому анализатору (опыты Заводского). На животных предварительно были установлены многочисленные двигательные и секреторные рефлексы, как безусловные, так и условные, последние как натуральные, так и искусственные. Из всех рефлексов после обоюдостороннего и полного удаления g. pyriformis и прилежащей части cornus ammonis как раз ранее всего появились именно запаховые рефлексы. Движения ноздрями на запах уже имелись на второйтретий день. На третий-четвертый дни собака из бумажных свертков только при помощи носа безошибочно выбирала содержащие мясо и колбасу. Начиная с шестого дня появился слюнной рефлекс на запах мясного порошка. Искусственный пищевой рефлекс на камфору отчетливо выступил с четырнадцатого дня после операции, раньше всех остальных условных искусственных рефлексов с кожи, уха и глаза. Искусственные запаховые рефлексы появлялись при первой пробе, т. e. восстанавливались, а не образовывались вновь.
Наконец мы поставили перед собой вопрос относительно натуры так называемой двигательной области коры полушарий. Что она такое? Такой ли анализатораздражений, идущих из скелетно-двигательного аппарата организма при его сложной и тонкой работе, как другие области коры суть анализаторы раздражений, падающих на организм извне, или это совершенно отличная от всех остальных область эффекторного характера; иначе сказать, отвечает ли она по физиологической функции задней или передней половине спинного мозга? Как известно, этот вопрос почти так же стар, как и самый факт констатирования двигательной области, и до сих пор имеет немало защитников решения его в первом смысле. Мы рассчитывали получить лишний материал для решения его при помощи метода условных рефлексов, делая из определенного акта движения условный раздражитель и устанавливая опытно топографию его в коре. Вот как были исполнены наши опыты (опыты Красногорского). Так как к этим опытам привязывается важный вопрос об общей конституции больших полушарий и так как они довольно сложны сравнительно со всеми остальными опытами над условными рефлексами по их постановке, то я считаю себя вправе передать эти опыты возможно подробно и обильно снабдить описание их протоколами.
Условный пищевой раздражитель был сделан из сгибания голеностопного и предплюсно-фалангового суставов. Сгибание производилось следующим образом. Бедро и голень одной из задних конечностей были фиксированы гипсовой съемной шиной, укрепленной на металлической подставке, привинченной к станку. Движение производилось в голеностопном суставе. Когда пользовались предплюсно-фаланговым суставом, также на особой подставке фиксировалась плюсна и предплюсна. Сгибание производилось прибором или рукой.
Когда рефлекс образовался на сгибание в голеностопном суставе левой конечности, он был испытан на правой. Он воспроизвелся и здесь сам собой, как мы с этим встречались при кожных условных раздражениях. Затем была предпринята выработка дифференцировки на левой же конечности сгибания фалангового сустава от сгибания голеностопного, причем сопровождалось подкармливанием животного только сгибание фалангового. Дифференцирование было достигнуто после 42 подкреплений сгибания фалангового сустава и 74 повторений сгибания голеностопного сустава без сопровождения безусловным раздражителем. Дифференцирование также само собой оказалось и на правой конечности. Но это, конечно, не было еще то, что было нам нужно. При сгибании непременно участвовало и кожно-механическое раздражение, и оно одно могло обусловливать условный эффект. Необходимо было отдифференцировать его от акта сгибания. Поэтому применялись всевозможные формы механического раздражения кожи: прикосновение, давление, захватывание, ритмическое натягивание кожи на одной стороне сустава и складывание на противоположной (как это приблизительно происходит при сгибании). Всего сильнее действовал последний прием. Повторяемые, конечно при отсутствии подкармливания животного, одни, без сгибания, все эти приемы сделались, наконец, совершенно недействительными. Сгибалие продолжало действовать. Но и это не казалось убедительным в отношении непременного участия в нашем условном раздражителе именно самого акта сгибания. Было возможно, что мы всеми нашими вариациями механического раздражения не воспроизводили его полностью, как оно происходит при сгибании. Необходимо было другое, более бесспорное доказательство, что акт сгибания действительно сделан нами условным раздражителем. Мы думали иметь это доказательство при полном исключении кожного компонента посредством удаления части кожного анализатора, о которой мы знали, что в ней локализировались кожные раздражения задней конечности, именно gg. coronarius и ectosylvius. Предварительно были выработаны условные пищевые рефлексы на механическое раздражение кожи задней конечности в пяти местах и на тон в 500 колебаний в секунду. После операции на левой стороне первый условный рефлекс на тон появился на седьмой день. Первая проба сгибания на правой стороне на восьмой день после операции оказалась недействительной. Вторая же в том же опыте уже дала капли в течение 30 секунд. На десятый день получились 3 капли. В этот же день испытанное действие одновременно пятью приборами для механического раздражения кожи на той же конечности оказалось нулевым в отношении слюноотделения. На двенадцатый день, когда рефлекс на сгибание достиг 5 капель, как механическое раздражение пятью приборчиками, так и натягивание и складывание кожи на суставе оставались нулевыми. На тринадцатый день натягивание и складывание кожи на суставе тормозили одновременно с ним примененное раздражение тоном; значит, механическое раздражение действовало не положительно, а тормозящим образом. На пятнадцатый и шестнадцатый дни, когда левая задняя конечность освободилась от тормозящего действия операции, и механическое раздражение ее одной давало значительный слюнной эффект, то же механическое раздражение ее одновременно с натягиванием и складыванием кожи на правом фаланговом суставе становилось нулевым. Следовательно, на правой конечности, продолжающей оставаться под влиянием операции, кожно-механическое раздражение было тормозящим, а не положительным. Сгибание же неизменно сопровождалось слюнос делением.
Привожу протоколы некоторых из этих опытов.
Все четыре рефлекса нужно было вырабатывать отдельно Точно так же всякие посторонние раздражения кожи одной половины тела (тепло в 50°С, слабый электрический ток в случае пищевых и кислотных рефлексов), вызывая исследовательский. рефлекс, тормозили условные кожно-механические рефлексы только той же половины тела, совершенно не влияя на такие же рефлексы другой половины тела. Все эти опыты исполнены на 3 животных.
Рядом с специальными опытами над кожным анализатором могут быть упомянуты наши опыты (опыты Бабкина) с удалением только лобных долей. Удалялись лобные доли с обеих сторон по sulcus praecruciatus и sulcus praesylvius до основания мозга, следовательно, с разрушением lobi olfactorii. Иногда нож при операции и почти постоянно последующий патологический процесс, как показывало потом вскрытие, захватывали и дальнейшие кзади лежащие извилины. ы произведены на. 4 собаках. Во всех случаях условные рефлексы с глаза и уха после операции восстанавливаются или образуются более или менее быстро. Нарушения, и значительные, были только со стороны кожного анализатора и движения. Большей частью нельзя было образовать, пока животные жили (от одного месяца до шести, умирая во время судорог), условных положительных кожно-механических рефлексов специально c туловища. C конечностей иногда вырабатывались. Отрицательные же рефлексы в виде условного тормоза могли быть получены со всех мест кожи. Рядом с этим почти постоянно существовала общая повышенная возбудимость кожной поверхности, так что некоторые собаки не выносили петель на ногах на станке во время опыта и стояли на станке спокойно только без них. Также наблюдались и временные двигательные расстройства в виде измененной позы животных (опущенная вниз голова и выгнутая кверху спина) и паретических явлений в конечностях вместе с подергиваниями. Особенно были сильны расстройства в движениях рта. В ближайшее время после операции животные с трудом захватывали в особенности твердую пищу, так что приходилось кормить их с руки. Обшее же поведение животного, за исключением перечисленных частностей, не представляло ничего особенного.
Из небольшого числа наших опытов словными рефлексами на агенты кожно-температурного анализатора, повидимаму, следует, что этот анализатор и кожио-механический топографически в коре не совсем совпадают. При удалении g. praecruciatus (опыты Шишло), в то время как условный кожномеханический рефлекс на задней конечности восстановился быстро (около недели), температурный (нлод и на температуру 47.5° C) отстал в его появлении очень резко, на 4 недели.
При помощи метода условных рефлексов мы рассчитывали получить данные относительно указываемых некоторыми авторами отношений g. pyriformis к запаховому анализатору (опыты Заводского). На животных предварительно были установлены многочисленные двигательные и секреторные рефлексы, как безусловные, так и условные, последние как натуральные, так и искусственные. Из всех рефлексов после обоюдостороннего и полного удаления g. pyriformis и прилежащей части cornus ammonis как раз ранее всего появились именно запаховые рефлексы. Движения ноздрями на запах уже имелись на второйтретий день. На третий-четвертый дни собака из бумажных свертков только при помощи носа безошибочно выбирала содержащие мясо и колбасу. Начиная с шестого дня появился слюнной рефлекс на запах мясного порошка. Искусственный пищевой рефлекс на камфору отчетливо выступил с четырнадцатого дня после операции, раньше всех остальных условных искусственных рефлексов с кожи, уха и глаза. Искусственные запаховые рефлексы появлялись при первой пробе, т. e. восстанавливались, а не образовывались вновь.
Наконец мы поставили перед собой вопрос относительно натуры так называемой двигательной области коры полушарий. Что она такое? Такой ли анализатораздражений, идущих из скелетно-двигательного аппарата организма при его сложной и тонкой работе, как другие области коры суть анализаторы раздражений, падающих на организм извне, или это совершенно отличная от всех остальных область эффекторного характера; иначе сказать, отвечает ли она по физиологической функции задней или передней половине спинного мозга? Как известно, этот вопрос почти так же стар, как и самый факт констатирования двигательной области, и до сих пор имеет немало защитников решения его в первом смысле. Мы рассчитывали получить лишний материал для решения его при помощи метода условных рефлексов, делая из определенного акта движения условный раздражитель и устанавливая опытно топографию его в коре. Вот как были исполнены наши опыты (опыты Красногорского). Так как к этим опытам привязывается важный вопрос об общей конституции больших полушарий и так как они довольно сложны сравнительно со всеми остальными опытами над условными рефлексами по их постановке, то я считаю себя вправе передать эти опыты возможно подробно и обильно снабдить описание их протоколами.
Условный пищевой раздражитель был сделан из сгибания голеностопного и предплюсно-фалангового суставов. Сгибание производилось следующим образом. Бедро и голень одной из задних конечностей были фиксированы гипсовой съемной шиной, укрепленной на металлической подставке, привинченной к станку. Движение производилось в голеностопном суставе. Когда пользовались предплюсно-фаланговым суставом, также на особой подставке фиксировалась плюсна и предплюсна. Сгибание производилось прибором или рукой.
Когда рефлекс образовался на сгибание в голеностопном суставе левой конечности, он был испытан на правой. Он воспроизвелся и здесь сам собой, как мы с этим встречались при кожных условных раздражениях. Затем была предпринята выработка дифференцировки на левой же конечности сгибания фалангового сустава от сгибания голеностопного, причем сопровождалось подкармливанием животного только сгибание фалангового. Дифференцирование было достигнуто после 42 подкреплений сгибания фалангового сустава и 74 повторений сгибания голеностопного сустава без сопровождения безусловным раздражителем. Дифференцирование также само собой оказалось и на правой конечности. Но это, конечно, не было еще то, что было нам нужно. При сгибании непременно участвовало и кожно-механическое раздражение, и оно одно могло обусловливать условный эффект. Необходимо было отдифференцировать его от акта сгибания. Поэтому применялись всевозможные формы механического раздражения кожи: прикосновение, давление, захватывание, ритмическое натягивание кожи на одной стороне сустава и складывание на противоположной (как это приблизительно происходит при сгибании). Всего сильнее действовал последний прием. Повторяемые, конечно при отсутствии подкармливания животного, одни, без сгибания, все эти приемы сделались, наконец, совершенно недействительными. Сгибалие продолжало действовать. Но и это не казалось убедительным в отношении непременного участия в нашем условном раздражителе именно самого акта сгибания. Было возможно, что мы всеми нашими вариациями механического раздражения не воспроизводили его полностью, как оно происходит при сгибании. Необходимо было другое, более бесспорное доказательство, что акт сгибания действительно сделан нами условным раздражителем. Мы думали иметь это доказательство при полном исключении кожного компонента посредством удаления части кожного анализатора, о которой мы знали, что в ней локализировались кожные раздражения задней конечности, именно gg. coronarius и ectosylvius. Предварительно были выработаны условные пищевые рефлексы на механическое раздражение кожи задней конечности в пяти местах и на тон в 500 колебаний в секунду. После операции на левой стороне первый условный рефлекс на тон появился на седьмой день. Первая проба сгибания на правой стороне на восьмой день после операции оказалась недействительной. Вторая же в том же опыте уже дала капли в течение 30 секунд. На десятый день получились 3 капли. В этот же день испытанное действие одновременно пятью приборами для механического раздражения кожи на той же конечности оказалось нулевым в отношении слюноотделения. На двенадцатый день, когда рефлекс на сгибание достиг 5 капель, как механическое раздражение пятью приборчиками, так и натягивание и складывание кожи на суставе оставались нулевыми. На тринадцатый день натягивание и складывание кожи на суставе тормозили одновременно с ним примененное раздражение тоном; значит, механическое раздражение действовало не положительно, а тормозящим образом. На пятнадцатый и шестнадцатый дни, когда левая задняя конечность освободилась от тормозящего действия операции, и механическое раздражение ее одной давало значительный слюнной эффект, то же механическое раздражение ее одновременно с натягиванием и складыванием кожи на правом фаланговом суставе становилось нулевым. Следовательно, на правой конечности, продолжающей оставаться под влиянием операции, кожно-механическое раздражение было тормозящим, а не положительным. Сгибание же неизменно сопровождалось слюнос делением.
Привожу протоколы некоторых из этих опытов.
 Эти опыты дают право сделать два заключения: первое, что один двигательный акт без участия кожного компонента может быть сделан условным раздражителем, и второе, что раздражение от двигательного акта и кожно-механическое раздражение в коре имеют различные области их приложения. Мы знаем область для кожно-механического раздражения. Где же область для двигательного акта? Ответы дают опыты на другой нашей собаке.
У нас имелась собака с основательно удаленным два месяца с лишком тому назад g. sygmoideus dexter, y которой все время после операции оставались резкие двигательные расстройства в обеих конечностях левой стороны. Повреждение g. sygmoideus sinister было очень незначительно и ничем не выражалось на конечностях правой стороны. Кожно-механические рефлексы были нормальны на всей поверхности тела. На правой стороне было приступлено к выработке пищевого условного рефлекса на сгибание фалангового сустава. Он был готов быстро. Теперь его стали дифференцировать от кожного компонента, применяя вышеупомянутые вариации механического раздражения кожи на суставе, конечно не сопровождая их безусловным рефлексом. Через месяц дифференцировка образовалась, хотя и не всегда полная.
Вот пример ее.
Эти опыты дают право сделать два заключения: первое, что один двигательный акт без участия кожного компонента может быть сделан условным раздражителем, и второе, что раздражение от двигательного акта и кожно-механическое раздражение в коре имеют различные области их приложения. Мы знаем область для кожно-механического раздражения. Где же область для двигательного акта? Ответы дают опыты на другой нашей собаке.
У нас имелась собака с основательно удаленным два месяца с лишком тому назад g. sygmoideus dexter, y которой все время после операции оставались резкие двигательные расстройства в обеих конечностях левой стороны. Повреждение g. sygmoideus sinister было очень незначительно и ничем не выражалось на конечностях правой стороны. Кожно-механические рефлексы были нормальны на всей поверхности тела. На правой стороне было приступлено к выработке пищевого условного рефлекса на сгибание фалангового сустава. Он был готов быстро. Теперь его стали дифференцировать от кожного компонента, применяя вышеупомянутые вариации механического раздражения кожи на суставе, конечно не сопровождая их безусловным рефлексом. Через месяц дифференцировка образовалась, хотя и не всегда полная.
Вот пример ее.
 Натягивание и складывание кожи на фаланговом суставе левой стороны тоже оказывалось совсем недействительным в силу вышеуказанных отношений. Но здесь не было рефлекса и на сгибание. Когда же сгибание было подкреплено, вслед за этим и одно механическое раздражение приобретало слюногонное действие. Опыт надо было понять так, что сгибание на левой стороне само по себе не действовало, но подкрепление его растормаживало неизбежный при сгибании кожный компонент. A дальше, несмотря на настойчиво продолжаемую процедуру дифференцирования, не удалось получить условный рефлекс на сгибание отдельно от раздражения кожи. Как только уничтожался эффект от одного раздражения кожи, исчезал рефлекс и от сгибания. При подкреплении возвращался рефлекс с кожи. На правой же стороне дифференцировка между сгибанием и кожным раздражением выступала постоянно полной отчетливостью.
Вот пример.
Натягивание и складывание кожи на фаланговом суставе левой стороны тоже оказывалось совсем недействительным в силу вышеуказанных отношений. Но здесь не было рефлекса и на сгибание. Когда же сгибание было подкреплено, вслед за этим и одно механическое раздражение приобретало слюногонное действие. Опыт надо было понять так, что сгибание на левой стороне само по себе не действовало, но подкрепление его растормаживало неизбежный при сгибании кожный компонент. A дальше, несмотря на настойчиво продолжаемую процедуру дифференцирования, не удалось получить условный рефлекс на сгибание отдельно от раздражения кожи. Как только уничтожался эффект от одного раздражения кожи, исчезал рефлекс и от сгибания. При подкреплении возвращался рефлекс с кожи. На правой же стороне дифференцировка между сгибанием и кожным раздражением выступала постоянно полной отчетливостью.
Вот пример.
 Сгибание на обеих сторонах во всех случаях этого опыта сопровождалось подкармливанием, раздражение кожи - нет.
Эт Эти опыты показывают, что g. sygmoideus есть область приложения раздражений, идущих из скелетно-двигательного аппарата при его работе. К сожалению, мы потом не продолжали и не разнообразили описанные опыты. Конечно, требуется их повторение и подтверждение. Если опираться на них, то следует принимать, что двигательная область коры есть такой же анализатор скелетно-двигательной энергии организма, как другие области ее суть анализаторы разных видов внешней энергии, действующей на организм. При таком взгляде на дело полушария представляют собой грандиозный анализатор как внешнего мира, так и внутреннего мира организма. Конечно, то, что надо принять относительно скелетно-двигательной деятельности, есть основание распространить на гораздо большую деятельность организма, если не на всю. Огромную роль внушения с ее чрезвычайными случаями, как, например, мнимая беременность, можно физиологически понять только при допущении коркового анализатора, пусть обычно мало расчлененного и неопределенного, анализатора деятельности всего организма.
Сгибание на обеих сторонах во всех случаях этого опыта сопровождалось подкармливанием, раздражение кожи - нет.
Эт Эти опыты показывают, что g. sygmoideus есть область приложения раздражений, идущих из скелетно-двигательного аппарата при его работе. К сожалению, мы потом не продолжали и не разнообразили описанные опыты. Конечно, требуется их повторение и подтверждение. Если опираться на них, то следует принимать, что двигательная область коры есть такой же анализатор скелетно-двигательной энергии организма, как другие области ее суть анализаторы разных видов внешней энергии, действующей на организм. При таком взгляде на дело полушария представляют собой грандиозный анализатор как внешнего мира, так и внутреннего мира организма. Конечно, то, что надо принять относительно скелетно-двигательной деятельности, есть основание распространить на гораздо большую деятельность организма, если не на всю. Огромную роль внушения с ее чрезвычайными случаями, как, например, мнимая беременность, можно физиологически понять только при допущении коркового анализатора, пусть обычно мало расчлененного и неопределенного, анализатора деятельности всего организма.
Лекция двадцать первая. Патологические состояния больших пполушарий как результат оперативных воздействий на них: проба вывести отклонения от нормы в поведении животного из частных нарушений коры больших полушарий
Мм. гг.! Мы знаем, что удаление всей массы полушарий у собаки превращает ее в относительно проступ рефлекторную машину, располагающую сравнительно небольшой суммой внешних безусловных рефлексов и лишенную той сложности и тонкости соотношения ее работы с внешним миром, основу которых составляют бесчисленные условные рефлексы, являющиеся верховной функцией полушарий. С другой стороны, у нас имеются некоторые сведения о значении отдельных частей полушарий, анализаторов, совокупная деятельность которых и обусловливает полное уравновешивание организма с окружающей средой, иначе говоря, определяет поведение животного. Для приближения к более или менее полному пониманию физиологической деятельности полушарий во всей ее целостности предоставляется, между прочим, небезвыгодным утилизировать, т. е. наблюдать и анализировать состояние животного как при удачном, т. е. без осложнений, удаленииспециально больших порций массы полушарий, так и при случаях последовательного и значительного нарушения полушарий разращением рубцовой ткани или другими побочными обстоятельствами операции. Другими словами, это значит - стараться все более и более при всяком случае сводить изменения общего поведения животного к тем или другим поломам, нарушениям механизма больших полушарий. Этому я и посвящу настоящую лекцию, переходя от более простых случаев к более сложным. У одной собаки (опыты Орбели) были удалены верхние половины обоих полушарий на высоте вершина g. sylviaticus. Удаление на каждой стороне произведено одним разрезом, так что удаленный кусок точно свидетельствует о размерах дефекта на массе полушарий. Через 2 недели после второй операции (операции на обеих сторонах исполнены с большим промежутком) собака представляла состояние, которое оставалось затем неизменным до конца ее исследования (4 месяца). Это и теперь было в высшей степени подвижное животное, быстро отзывающееся на кличку, легко и круто поворачивающееся и направляющееся на зов. При обычном взгляде нельзя было бы отличить ее от нормального животного. Только при внимательном наблюдении замечалась некоторая атактичность в конечностях, они подбрасывались и ударялись по полу при беге больше, чем обыкновенно, а при ходьбе по гладкому или мокрому полу часто скользили. Кроме того, собака проделывала особенные движения головой, когда направлялась куда-нибудь. Только изредка она наталкивалась на предметы, стоящие на пути ее движения. Но как только происходило последнее, выступала поражающая ненормальность собаки. Она оказывалась совершенно беспомощной при столкновении даже с довольно тонков ножкой стола. Она долгое время толкалась о нее, пока случайным боковым отклонением не миновала ее, и только таким образом получала возможность продолжать дальнейшее движение. Положенная передней половиной тела на стул, она при зове или беспорядочным движением сваливалась со стула боком, или, двигаясь вперед, укладывалась животом на стуле и, болтая ногами в воздухе, не могла ничего больше сделать, и т. д. и т. д. в этом роде Условные рефлексы, бывшие до второй операции, скоро восстановились после нее, и могли быть легко образованы новые на запах и освещение. Не восстановился кожно-механический условный рефлекс, и не мог быть образован температурный (на охлаждение). Безусловные рефлексы на разные раздражения кожи (встряхивание, отдергивание конечностей, визжание, поворот головой к месту действия приборчиков для механического раздражения и охлаждения) существовали. Как понимать механизм отклонений от нормы в поведении этой собаки, имея в виду функции анализаторов? Деятельность запахового и звукового анализаторов была налицо в полном объеме. То же с небольшим ограничением нужно сказать и относительно светового анализатора. Обращая внимание на полную ориентировку собаки глазом и особенные движения ее головой при передвижениях, нужно принять, что у собаки уцелела после операции небольшая нижняя часть специального района светового анализатора, которая при соответственных постановках головы делала возможным высший синтез и анализ световых раздражений. Радикально пострадал двигательный анализатор. Общая локомоторная деятельность, совершающаяся насчет частей головного мозга, лежащих ниже полушарий, явно совершенно сохранилась, а подробная и тонкая деятельность скелетной мускулатуры, по крайней мере части ее, основанная на условных рефлексах, исчезла. Что касается кожного анализатора, то этот пункт сейчас нам представляется неясным. Конечно, факт потери двигательной ориентировки при встрече собакой препятствий на пути ее движений вполне согласовался бы с отсутствием правильных сигналов с кожи. Но отсутствие условных кожно-механических рефлексов при данной операции стоит в некотором противоречии с другими нашими опытами относительно топографии кожного анализатора. Требуется нарочитая проверка относительно этого пункта, специально требуется испытание всех пунктов кожи. Итак, в целом поведение собаки как в общей природной, так и специально в социальной среде осталось нормальным, за исключением частного дефекта в обхождении с встречающимися механическими препятствиями. Оперированная таким образом собака, лишенная совершенно определенной и значительной массы больших полушарий, представляет большой интерес для более подробного и более точного анализа ее, чем исполненный над ней нами в ранний период нашей работы. У ряда других собак, которые уже упоминались раньше, была удалена задняя большая половина полушарий по указанной выше линии, начинающейся вверху позади g. sygmoideus и кончающийся внизу по fissura fossae Sylvil. Я возвращаюсь к этим собакам сейчас, чтобы привлечь внимание на такие стороны их поведения, которые не нашли себе места в прежнем изложении. Напоминаю, что у них отсутствовал высший синтез и анализ как звуковых, так и световых раздражений, но существовал анализ разных родов звука и анализ интенсивности освещения и форм. Прямо после окончательной операции они почти сплошь спали, а затем во все продолжение жизни (одна 3 года) главным образом проводили время во сне и посему при хорошем кормлении быстро жирели. Когда были голодны, исключительно при помощи запахового и кожного анализаторов, как уже сказано раньше, находили пищу. Особенно была поразительна соединенная работа кожно-механического и двигательного анализаторов. Когда животное находилось среди разбросанных по полу и подвешенных на нитках на разной высоте кусочков мяса, то малейшего прикосновения к куску мяса какой-нибудь частью тела было достаточно, чтобы вызвать точнейшее движение тела в пункт местонахождения этого куска и захватывание его ртом. Наконец особенно должно быть отмечено совершенно индифферентное отношение таких собак к другим собакам и людям, не исключая хозяина-экспериментатора. Нетрудно представить себе большую жизненную пассивность (уже отмеченную раньше Кольцом), большую наклонность ко сну этих животных, раз у них отпала в значительной степени деятельность главнейших дистантных анализаторов - светового и звукового. Точно так же является совершенно понятной чрезвычайная изощренность деятельностей сохранившихся в целости запахового, кожного и двигательного анализаторов. Но останавливает на себе внимание упомянутая последняя особенность. Что значит она? Есть ли это частное явление их общей жизненной пассивности, общего упадка энергии, или, что вероятнее, специальный результат выпадения комплексных световых и звуковых раздражений как преимущественных социальных раздражений, а также, может быть, и цепных условных рефлексов? При теперешнем состоянии наших сведений о высшей нервной деятельности этот вопрос мог бы подлежать серьезному исследованию, и он его заслуживает. Затем я перехожу к самому сложному и самому поучительному нашему случаю - к случаю с удалением передних половин обоих полушарий по вышеуказанной линии. Поведение этих животных в высшей степени отклоняется от нормы, и анализ его является чрезвычайно интересным. Поэтому я остановлюсь на нем особенно подробно. У нас имелось два таких животных, и они жили подолгу (около года). Операция у обеих собак была исполнена в два приема, сперва на одной стороне, затем, с промежутком в несколько месяцев, на другой. Я опишу сперва наиболее полно первую собаку (опыты В. А. Демидова), а затем сделаю нужные добавления относительно второй (опыты H. М. Сатурнова и С. П. Кураева). По После второй операции собака почти сплошь спала, просыпаясь только перед мочеиспусканием и испражнением. Пища вкладывалась животному через желудочную фистулу, наложенную до второй мозговой операции. Только с начала третьей недели после операции собака начала вставать и становиться на ноги, но стояла на ногах короткое время, а потом, покачиваясь, падала. Приблизительно через месяц она стала ходить, но при этом пальцы часто подвертывались, а конечности перекрещивались. Еще через месяц ходьба и бег сделались почти нормальными, но при крутых и быстрых поворотах собака едва удерживалась на ногах. При встрече с препятствиями собака делала беспорядочные движения. То двигаясь вперед, то подаваясь назад, то отклоняясь в сторону, иногда случайно сама преодолевала препятствия, а в большинстве случаев ей надо было помогать. Кроме того, обращало на себя внимание вообще невозможное комбинирование собакой в одно и то же время разных движений, в результате чего наступала потеря равновесия и падение животного. Эта особенность оставалась у собаки без изменения до конца жизни. Только через 2 недели после операции она сама начала лакать молоко, когда ее рот приводили в соприкосновение с ним. И потом всю жизнь она приступала к еде только тогда, когда пища прикасалась к слизистой губ, щек и особенно языка. Прикосновение пищи к коже около рта акта еды не вызывало. В позднейшее время, когда собака была голодна, она приходила в возбуждение и хватала ртом все, что находилось поблизости, даже собственную лапу, укусив которую, визжала. С самого начала еды собакой отчетливо отличались съедобные вещества от несъедобных, как песок, или содержащих примеси, например хинина, значительного процента кислоты, соли и т. д. Двигательная реакция на прикосновение к коже появилась тоже около 2 недель после операции. Затем кожная возбудимость все росла. Через 2-3 месяца прикосновение к животному, при помещении его в станок, при снятии со станка, даже простое поглаживание приводило собаку в сильнейшее возбуждение. Она рвалась, лаяла и скалила зубы. Точно так же при движениях, при которых она соприкасалась с разными предметами, на дворе, когда ветер шевелил ее волосы или капли дождя падали на нее, тоже наступало двигательное возбуждение. Интересно, что во время таких возбуждений поглаживание по голове или шее, наоборот, успокаивало животное, и оно даже иногда при этом погружалось в сон. 1ри чесании определенных мест кожи точно выступал чесательный рефлекс; при этом часто приходилось наблюдать появление или усиление рефлекса, если чесание было слабо, благодаря одновременно производимому звуковому раздражению явление так называемого Bahnungsreflex'a. Двигательная реакция на звук - поднятие и трясение ушей - появилась только через 14/2 месяца после операции. Затем реакция на звуки все усиливалась и иногда становилась чрезвычайной; даже при относительно слабых звуках собака впадала в сильное возбуждение. При действии сильного света она закрывала глаза и отворачивала голову. На запахи не было никогда и никакой реакции, так как у этой собаки были удалены bulbus и tractus olfactorii с обеих сторон. Полового рефлекса при нарочитых испытаниях наблюдать не удалось. Никаких специальных отношений, ни положительных, ни отрицательных, ни к другим животным, ни к людям никогда определенно не замечалось. С собакой случались как бы беспричинные припадки, выражавшиеся в дрожи всего тела, клонических судорогах челюстей с насильственным поворотом головы в одну сторону и кончавшиеся извержением мочи и кала. Судорог туловища и конечностей при этом не было, и животное не падало. Припадок продолжался 1-2 минуты. После него животное делалось возбужденным: бегало, металось и лаяло, затем успокаивалось, становилось сонливым и скоро глубоко засыпало. Таким образом, судя по деятельности скелетной мускулатуры, наша собака оказывалась совершенно лишенной высшей нервной деятельности, представляя собой очень упрощенную, далеко не совершенную рефлекторную машину. Походя во многом на собаку совсем без полушарий, она даже уступает ей в отношении локомоции, так как собака без полушарий скорее после операции начинает вставать на ноги и ходить и лучше сохраняет равновесие при разных движениях, чем наша. От условнорефлекторной деятельности, повидимому, не осталось и следа. Но так ли это? Попробуем анализировать ее состояние при помощи другого индикатора, помимо скелетной мускулатуры. Обратимся к нашему всегдашнему в описываемых исследованиях участнику высшей нервной деятельности - слюнной железе. Безусловный слюнной рефлекс прямо после операции тоже совершенно исчезал, но затем скоро восстанавливался, сначала с некоторыми отступлениями от нормы, но потом с течением времени изгладившимися. Положительных слюнных рефлексов от раздражения с глаза, уха и кожи получить не удалось, несмотря на энергичные меры, при этом принимаемые. Так, только под звук бульканья собака долгое время получала всю ежедневную пищу. Тем не менее при пятисоткратном применении этого приема бесспорного условного слюнного рефлекса не получилось. Тогда мы испытали раздражения с рецепторной поверхности, о которой уже раньше было сообщено, что она при различных нарушениях больших полушарий оказывается устойчивее всех других поверхностей, именно с ротовой поверхности, в виде водяного условного рефлекса. Эти опыты я и передам теперь в подробностях. Из раннего сообщения мы знаем, что значительный слюнной рефлекс на воду, введенияю в рот, получается только после предварительного введения в рот отвергаемых веществ в виде водяных растворов, например кислоты, так как вода сама по себе или совсем не вызывает слюноотделения, или только минимальное. При многократном введении кислоты нашей собаке вода впервые обнаружила значительное слюногонное действие около месяца спустя после операции. 50 дней спустя после операции были начаты систематические опыты с водяным рефлексом. Когда кислота вливалась часто, изо дня в день, то в день опыта вода, примененная в опыте как первый раздражитель, т. е. без предварительного введения кислоты, вызывала обильное слюноотделение, до и более капель за минуту. Это слюноотделение исчезало, если повторно вливалась одна вода. Вот примеры таких опытов.
 Очевидно, это есть угасание условного водяного рефлекса. Условный водяной рефлекс так же легко, как и вообще условные рефлексы, тормозился всякими посторонними рефлексами (внешнее торможение).
Вот примеры.
Очевидно, это есть угасание условного водяного рефлекса. Условный водяной рефлекс так же легко, как и вообще условные рефлексы, тормозился всякими посторонними рефлексами (внешнее торможение).
Вот примеры.
 Но это (внешнее) торможение не характерно для условных рефлексов, так как так же тормозятся и все безусловные рефлексы. Поэтому было предпринятобразование условного тормоза, т. е. вида внутреннего торможения, условного отрицательного раздражителя. что взять тормозным агентом? Расчет был на то, что, может быть, агенты, относящиеся к другим анализаторам, не делаясь положительноми условными раздражителями, окажутся отрицательными, чему несколько примеров уже встречалось в прошлых лекциях. Расчет оправдался. Звуковые и световые раздражители могли сделаться условными тормозами. Вливание воды, одновременное со звучанием определенного тона, стало постоянной тормозной комбинацией при 64-м повторении.
Вот примеры.
Но это (внешнее) торможение не характерно для условных рефлексов, так как так же тормозятся и все безусловные рефлексы. Поэтому было предпринятобразование условного тормоза, т. е. вида внутреннего торможения, условного отрицательного раздражителя. что взять тормозным агентом? Расчет был на то, что, может быть, агенты, относящиеся к другим анализаторам, не делаясь положительноми условными раздражителями, окажутся отрицательными, чему несколько примеров уже встречалось в прошлых лекциях. Расчет оправдался. Звуковые и световые раздражители могли сделаться условными тормозами. Вливание воды, одновременное со звучанием определенного тона, стало постоянной тормозной комбинацией при 64-м повторении.
Вот примеры.

 То же было достигнуто при сильном освещении комнаты при вливании воды, когда кислота вливалась в затемненной комнате. Теперь условное торможение развилось быстрее. Вот опыт только при 16-м повторении вливания воды щением.
То же было достигнуто при сильном освещении комнаты при вливании воды, когда кислота вливалась в затемненной комнате. Теперь условное торможение развилось быстрее. Вот опыт только при 16-м повторении вливания воды щением.
 Наконец было испытано растормаживание. И оно вполне ясно выступало при соответствующих условиях. Вот опыт, поставленный в многолюдном заседании в Обществе русских врачей в С.-Петербурге 19 марта 1909 года.
Наконец было испытано растормаживание. И оно вполне ясно выступало при соответствующих условиях. Вот опыт, поставленный в многолюдном заседании в Обществе русских врачей в С.-Петербурге 19 марта 1909 года.
 Раздражение мясом растормозило угашенный водяной рефлекс только на ближайшее время его последействия, а позже угашение восстанавливалось до известного срока, как это обыкновенно и наблюдается.
Таким образом наличность условнорефлекторной деятельности была обнаружена и у этой собаки.
Теперь перехожу к другой подобной собаке. Эта собака была оперирована с некоторым видоизменением, именно при удалении передней половины обоих полушарий, по указанной линии кзади, были возможно пощажены bulbus и tractus olfactorii, так как представлялось нужным для большей убедительности рядом с уцелевшим условным водяным рефлексом иметь и условный рефлекс на запах.
Оказалось то, что мы ожидали. У этой собаки, кроме условного водяного рефлекса, восстановился и бывший до операции условный слюнной пищевой рефлекс на запах камфоры. Конечно, был и натуральный слюнной условный рефлекс на еду. Это же вело и к тому, что собака тянулась или двигалась по направлению к еде и могла случайно хватать еду, находящуюся близко от нее. Но этим и некоторыми другими второстепенными особенностями и ограничивалась разница поведении этой собаки от первой. Отношение собаки к другим животным и людям оставалось тоже совершенно безразличным. Беспомощность среди механических препятствий и локомоция недостатками были выражены так же ярко. Вообще и эта собака была такой же инвалидной, неспособной жить без помощи со стороны, как и первая. Положительных условных рефлексов на раздражения, относящиеся к другим анализаторам, получить было нельзя.
Остается еще прибавить, что у обеих собак при вскрытии после смерти оставшаяся задняя половина полушарий оказалась очень резко атрофированной.
Мне кажется, что поведение этих собак более или менее ясно, т. е. доступно анализу с точки зрения выпадения функции разрушенных и нарушенных анализаторов, как это обнаружилось в результате испытания условных рефлексов и данными вскрытия. Все корковые концы анализаторов, кроме ротового у первой и ротового и запахового у второй, или не функционировали совсем, или только ограниченно, в виде торможения. Следовательно, собаки были лишены огромной массы раздражений со стороны внешнего мира, обыкновенно определяющих нормальную и сложную деятельность животных. У периой собаки оставался в действии только анализатор, наиболее ограниченный в его отношениях с окружающей средой. У другой, кроме того, действовал и дистантный, особенно развитый у собак, анализатор запаховый. Но на этот раз он работал слабо и не очень постоянно. Можно думать, что это произошло потому, что или и этот анализатор также несколько пострадал при операции, или он постоянно более или менее находился под тормозящим влиянием других нарушенных анализаторов, неспособных на положительную деятельность, a отвечающих нашние раздражения иррадиирующим тормозным процессом. С другой стороны, обычный и главнейший рабочий приборганизма - скелетно-двигательная система - был лишен возможности совершать свою работу в точном соответствии с внешними условиями. Его работа в норме определяется тесно ассоциированным Функционированием двух анализаторов: внешнего кожного, сигнализирующего подробно внешние, механические отношения животного в окружающей среде, и внутреннего двигательного, детально анализирующего и сложно синтезирующего соответственные двигательные акты. При глубоком нарушении того и другого точных соответственных двигательных реакций, конечно, быть не может. Есть основания думать, что некоторые разрозненные части только что упомянутых анализаторов уцелели в коре. Так, может быть, надо понять, что с одних мест кожного покрова при раздражении вызываются безусловные рефлексы явно насчет работы низших двигательных отделов центральной нервной системы, например оборонительный, чесательный и т. д., с других (при поглаживании кожи головы и шеи) наступают, вероятно, условно-тормозящие (нейтрализующие возбуждение животного) рефлексы. Это же представление хорошо бы объяснило и разницу между описываемыми собаками и собаками совсем без больших полушарий, которые гораздо скорее после операции начинают вставать, стоять и ходить и вообще менее обнаруживают беспорядка в движениях, чем первые. На остатки двигательного анализатора указывает и то, что у занимающих нас теперь собак бывали эпилептоидные припадки в виде судорог головных, шейных и иногда туловищных мускулов, но никогда мускулов конечностей. Что касается отсутствия специально социальных рефлексов, то могу сослаться на то, что сказано мной раньше. Вероятно, для них главным образом необходимы комплексные раздражения, которых у этих собак, конечно, не могло быть, раз не было даже и положительных элементарных. Таким образом у первой собаки только орган второстепенный по физиологической роли и упрощенный, что касается соотношений с внешним миром, слюнная железа при содействии связанного с ним тоже ограниченного, но уцелевшего ротового анализатора свидетельствует о все еще продолжающем действовать механизме коры полушарий, высшей нервной деятельности животного.
Я остановлюсь еще на одной собаке, анализ которой тоже занял у нас немало времени. Она представила резкое отклонение от нормального поведения не сразу после операции вследствие удаления некоторой части полушарий, а под влиянием последующего разрастания рубцовой ткани, сопровождавшегося частыми судорожными припадками, но не сильными. Только последние припадки, особенно сильные, 2 года с лишком спустя после мозговой операции привели ее к гибели. Это была молодая, очень подвижная собака с сильной нервной системой. Работа над ней начата с образования у нее исключительно кожно-механических и кожно-температурных рефлексов, которые, как сообщено раньше, особенно располагают корковые клетки к переходу в тормозное состояние, так что животное под их влиянием обыкновенно очень скоро делается сонливым и погружается в сон. см не менее эта собака все время в станке оставалась бодрой. Операция была исполнена в два приема - 9 марта и 28 апреля 1910 г., сперва на одной стороне, потом на другой: были повреждены gg. postcruciati. Только теперь на короткое время при раздражении мест кожи, несколько пострадавших от операции, развилась сонливость, но введением лишнего сильного условного раздражителя на звонок она была скоро устранена. Небольшие дефекты, наступившие сейчас же после операции, скоро почти изгладились, и собака вообще держалась как нормальная. 11 мая 1910 г. у нее был первый судорожный припадок. Первая работа на ней закончилась к этому сроку, и она оставалась затем при лаборатории, значительное время не подвергаемая опытам. Летом повторялисьдорожные припадки, а осенью и зимой ухаживающим за животным служителем было отмечено, что собака делается особенной, именно: при прикосновении к ней она приходит в сильное возбуждение, рычит, лает, скалит зубы, чего раньше с ней никогда не случалось. С начала января 1911 г. она поступает на исследование к новому нашему сотруднику (Сатурнову). Теперь ее общее поведение представлялось в следующем виде. Вынутая из клетки и поставленная на пол, при сильном возбуждении она скоро успокаивается и остается стоять на одном и том же месте по нескольку десятков минут, иногда по часу, двигая только головой и шеей туда и сюда, обнюхивая воздух. Затем она делает движение вперед или по кругу и скоро кончает тем, что или мочится, или испражняется. Это, очевидно, и обусловило ее передвижение. А потом опять продолжается стояние на одном и том же месте. Когда приходит обычное время еды, она также начинает ходить и принюхивается. Подставляемая ей чашка с пищей также заставляет ее тянуться к пище и двигаться за ней, если ее отодвигают. Акт еды совершенно правильный. Стоит собака твердо, не качаясь, но при движении передние конечности несколько выбрасываются, а на гладком полу и при бысворотах слегка скользят. Однако падений почти никогда не бывает. На препятствия иногда собака натыкается, иногда же обходит. Полового рефлекса, несмотря на повторение опытов, видеть не удавалось. На кличку не реагирует ибще никаких обычных отношений ни к другим собакам, ни к людям не обнаруживает. Прикосновение к коже собаки на всех местах тела, но особенно на голове и шее (вероятно, специально шевеление волос), чем бы оно ни было произведено, -- рукой, предметом, на который животное наткнулась, другим животным, ветром, каплями дождя, - непременно сейчас же вызывает сильнейшее возбуждение, выражающееся в рычании, лае, скалении зубов и в общих движениях. Обыкновенно при этом собака поднимает голову кверху и почти никогда не направляет ее к тому месту, где произошло прикосновение. Вместе с тем постоянное прикосновение и давление на кожу, например приборами, прикрепляемыми к коже собаки, петлями на ногах, когда собака стоит на станке, оставляют собаку в покое. Как уже сказано, все время, пока собака жила, у нее периодически повторялись общие судорожные припадки, после которых она обыкновенно довольно быстро оправлялась.
Нам предстояло исследовать условнорефлекторную деятельность странного животного и попытаться таким образом, может быть, разъяснить ближайший нервный механизм ее поведения. Условнорефлекторная деятельность оказалась налицо. Условные рефлексы были пищевые. Очень легко и скоро восстановился давний рефлекс на звонок. После того был образован новый звуковой рефлекс на тон органной трубы в 300 колебаний в секунду. За ним был получен рефлекс на камфору. Точно так же выработалась дифференцировка к тону из терции и условный тормоз из ударов метронома к рефлексу на камфору. Но первому исследователю больной собаки (Сатурнову) не удалось восстановить ранних (бывших до заболевания) кожно-механических условных рефлексов. При этом исследовании было отмечено, что положительные условные раздражители очень наклонны переходить в тормозные и что тормозной процесс очень инертен, очень долго давая себя знать на положительных раздражителях.
Следующий исследователь собаки (Кураев), оценив только что указанную особенность собаки работая главным образом с коротким изолированным условным раздражением, без труда получил и условные кожно-механические рефлексы. Но тормозная тенденция нервной системы все нарастала вместе с повторением и усилением судорожных припадков, пока сильные припадки 9 мая 1912 г., продолжавшиес небольшими паузами почти 12 часов, не убили собаку.
Вскрытие мозга показало следующее. По снятии рубца дефект мозгового вещества захватил следующие извилины: заднюю часть g. postcruciatus, передние отделы g. suprasplenialis, entolateralis и ectolateralis, g. suprasylvius medius, верхнюю половину g. coronarius и отчасти g. ectosylvius medius. Кроме того, оказались резко атрофированными затылочные и височные доли. Размер мозга здесь уменьшен, и мозговые извилины уплощены. Передние доли полушарий, повидимому, остались совершенно неизмененными.
Как понимать с точки зрения этих нарушений отклонения от нормы в поведении собаки? Вскрытие собаки показало, что разрушение и нарушение полушарий захватило главным образом заднюю половину больших полушарий, только слегка коснувшись передней. Таким образом собака должна была быть подобной собаке без задней половины полушарий, что и сказывалось на условных рефлексах. К сожалению, мы не исследовали ее относительно комплексных звуковых и световых раздражителей. На их отсутствие указывало то, что собака не реагировала на кличку и не обнаруживала специальных отношений ни к другим собакам, ни к людям. Последнее могло зависеть также и от отсутствия у такой собаки цепных рефлексов. Далее некоторая неисправность в ходьбе, очевидно, зависела от небольшого повреждения двигательной области. Представляются трудными для удовлетворительного объяснения стояние собаки на одном месте и чрезвычайная реакция на прикосновение к коже. Что касается первого, то относить ли это на счет несомненного преобладания тормозного процесса в коре (под влиянием повторяющихся взрывов возбуждения) или рассматривать это как результат некоторого частичного повреждения кожного анализатора, тем более, что голова и шея сохранили свою обычную подвижность? Вместе с тем остается непонятным действие прикосновения к коже собаки. Корковый ли это рефлекс или низших отделов мозга? Раз условные кожно-механические рефлексы существуют, последнее маловероятно. Но тогда как согласить повышенную возбудимость кожного анализатора c преобладанием тормозного процесса в коре? Для решения этих вопросов требовались многие вариации опытов, которые в свое время не были сделаны.
Приводя описанные в этой лекции случаи, я отнюдь не имел претензии дать удовлетворительные объяснения нервного механизма всех отклонений от нормы в поведении собак после мозговых операций. Моя цель состояла только в том, чтобы поставить перед физиологией полушарий вопросы относительно этого механизма как законные и показать некоторую возможность решения их.
В целом приведенные в последних трех лекциях опыты с условными рефлексами главным образом являются подтверждением ранних и позднейших исследований других авторов относительно того же предмета, только сольшими прибавлениями с нашей стороны и с постановкой некоторых новых задач в этой области. Но наши факты решительно противоречат учению об отдельных ассоциационных центрах вли вообще о какомто особом отделе полушарий с верховной нервной функцией, против чего восставал уже и Мунк.
Раздражение мясом растормозило угашенный водяной рефлекс только на ближайшее время его последействия, а позже угашение восстанавливалось до известного срока, как это обыкновенно и наблюдается.
Таким образом наличность условнорефлекторной деятельности была обнаружена и у этой собаки.
Теперь перехожу к другой подобной собаке. Эта собака была оперирована с некоторым видоизменением, именно при удалении передней половины обоих полушарий, по указанной линии кзади, были возможно пощажены bulbus и tractus olfactorii, так как представлялось нужным для большей убедительности рядом с уцелевшим условным водяным рефлексом иметь и условный рефлекс на запах.
Оказалось то, что мы ожидали. У этой собаки, кроме условного водяного рефлекса, восстановился и бывший до операции условный слюнной пищевой рефлекс на запах камфоры. Конечно, был и натуральный слюнной условный рефлекс на еду. Это же вело и к тому, что собака тянулась или двигалась по направлению к еде и могла случайно хватать еду, находящуюся близко от нее. Но этим и некоторыми другими второстепенными особенностями и ограничивалась разница поведении этой собаки от первой. Отношение собаки к другим животным и людям оставалось тоже совершенно безразличным. Беспомощность среди механических препятствий и локомоция недостатками были выражены так же ярко. Вообще и эта собака была такой же инвалидной, неспособной жить без помощи со стороны, как и первая. Положительных условных рефлексов на раздражения, относящиеся к другим анализаторам, получить было нельзя.
Остается еще прибавить, что у обеих собак при вскрытии после смерти оставшаяся задняя половина полушарий оказалась очень резко атрофированной.
Мне кажется, что поведение этих собак более или менее ясно, т. е. доступно анализу с точки зрения выпадения функции разрушенных и нарушенных анализаторов, как это обнаружилось в результате испытания условных рефлексов и данными вскрытия. Все корковые концы анализаторов, кроме ротового у первой и ротового и запахового у второй, или не функционировали совсем, или только ограниченно, в виде торможения. Следовательно, собаки были лишены огромной массы раздражений со стороны внешнего мира, обыкновенно определяющих нормальную и сложную деятельность животных. У периой собаки оставался в действии только анализатор, наиболее ограниченный в его отношениях с окружающей средой. У другой, кроме того, действовал и дистантный, особенно развитый у собак, анализатор запаховый. Но на этот раз он работал слабо и не очень постоянно. Можно думать, что это произошло потому, что или и этот анализатор также несколько пострадал при операции, или он постоянно более или менее находился под тормозящим влиянием других нарушенных анализаторов, неспособных на положительную деятельность, a отвечающих нашние раздражения иррадиирующим тормозным процессом. С другой стороны, обычный и главнейший рабочий приборганизма - скелетно-двигательная система - был лишен возможности совершать свою работу в точном соответствии с внешними условиями. Его работа в норме определяется тесно ассоциированным Функционированием двух анализаторов: внешнего кожного, сигнализирующего подробно внешние, механические отношения животного в окружающей среде, и внутреннего двигательного, детально анализирующего и сложно синтезирующего соответственные двигательные акты. При глубоком нарушении того и другого точных соответственных двигательных реакций, конечно, быть не может. Есть основания думать, что некоторые разрозненные части только что упомянутых анализаторов уцелели в коре. Так, может быть, надо понять, что с одних мест кожного покрова при раздражении вызываются безусловные рефлексы явно насчет работы низших двигательных отделов центральной нервной системы, например оборонительный, чесательный и т. д., с других (при поглаживании кожи головы и шеи) наступают, вероятно, условно-тормозящие (нейтрализующие возбуждение животного) рефлексы. Это же представление хорошо бы объяснило и разницу между описываемыми собаками и собаками совсем без больших полушарий, которые гораздо скорее после операции начинают вставать, стоять и ходить и вообще менее обнаруживают беспорядка в движениях, чем первые. На остатки двигательного анализатора указывает и то, что у занимающих нас теперь собак бывали эпилептоидные припадки в виде судорог головных, шейных и иногда туловищных мускулов, но никогда мускулов конечностей. Что касается отсутствия специально социальных рефлексов, то могу сослаться на то, что сказано мной раньше. Вероятно, для них главным образом необходимы комплексные раздражения, которых у этих собак, конечно, не могло быть, раз не было даже и положительных элементарных. Таким образом у первой собаки только орган второстепенный по физиологической роли и упрощенный, что касается соотношений с внешним миром, слюнная железа при содействии связанного с ним тоже ограниченного, но уцелевшего ротового анализатора свидетельствует о все еще продолжающем действовать механизме коры полушарий, высшей нервной деятельности животного.
Я остановлюсь еще на одной собаке, анализ которой тоже занял у нас немало времени. Она представила резкое отклонение от нормального поведения не сразу после операции вследствие удаления некоторой части полушарий, а под влиянием последующего разрастания рубцовой ткани, сопровождавшегося частыми судорожными припадками, но не сильными. Только последние припадки, особенно сильные, 2 года с лишком спустя после мозговой операции привели ее к гибели. Это была молодая, очень подвижная собака с сильной нервной системой. Работа над ней начата с образования у нее исключительно кожно-механических и кожно-температурных рефлексов, которые, как сообщено раньше, особенно располагают корковые клетки к переходу в тормозное состояние, так что животное под их влиянием обыкновенно очень скоро делается сонливым и погружается в сон. см не менее эта собака все время в станке оставалась бодрой. Операция была исполнена в два приема - 9 марта и 28 апреля 1910 г., сперва на одной стороне, потом на другой: были повреждены gg. postcruciati. Только теперь на короткое время при раздражении мест кожи, несколько пострадавших от операции, развилась сонливость, но введением лишнего сильного условного раздражителя на звонок она была скоро устранена. Небольшие дефекты, наступившие сейчас же после операции, скоро почти изгладились, и собака вообще держалась как нормальная. 11 мая 1910 г. у нее был первый судорожный припадок. Первая работа на ней закончилась к этому сроку, и она оставалась затем при лаборатории, значительное время не подвергаемая опытам. Летом повторялисьдорожные припадки, а осенью и зимой ухаживающим за животным служителем было отмечено, что собака делается особенной, именно: при прикосновении к ней она приходит в сильное возбуждение, рычит, лает, скалит зубы, чего раньше с ней никогда не случалось. С начала января 1911 г. она поступает на исследование к новому нашему сотруднику (Сатурнову). Теперь ее общее поведение представлялось в следующем виде. Вынутая из клетки и поставленная на пол, при сильном возбуждении она скоро успокаивается и остается стоять на одном и том же месте по нескольку десятков минут, иногда по часу, двигая только головой и шеей туда и сюда, обнюхивая воздух. Затем она делает движение вперед или по кругу и скоро кончает тем, что или мочится, или испражняется. Это, очевидно, и обусловило ее передвижение. А потом опять продолжается стояние на одном и том же месте. Когда приходит обычное время еды, она также начинает ходить и принюхивается. Подставляемая ей чашка с пищей также заставляет ее тянуться к пище и двигаться за ней, если ее отодвигают. Акт еды совершенно правильный. Стоит собака твердо, не качаясь, но при движении передние конечности несколько выбрасываются, а на гладком полу и при бысворотах слегка скользят. Однако падений почти никогда не бывает. На препятствия иногда собака натыкается, иногда же обходит. Полового рефлекса, несмотря на повторение опытов, видеть не удавалось. На кличку не реагирует ибще никаких обычных отношений ни к другим собакам, ни к людям не обнаруживает. Прикосновение к коже собаки на всех местах тела, но особенно на голове и шее (вероятно, специально шевеление волос), чем бы оно ни было произведено, -- рукой, предметом, на который животное наткнулась, другим животным, ветром, каплями дождя, - непременно сейчас же вызывает сильнейшее возбуждение, выражающееся в рычании, лае, скалении зубов и в общих движениях. Обыкновенно при этом собака поднимает голову кверху и почти никогда не направляет ее к тому месту, где произошло прикосновение. Вместе с тем постоянное прикосновение и давление на кожу, например приборами, прикрепляемыми к коже собаки, петлями на ногах, когда собака стоит на станке, оставляют собаку в покое. Как уже сказано, все время, пока собака жила, у нее периодически повторялись общие судорожные припадки, после которых она обыкновенно довольно быстро оправлялась.
Нам предстояло исследовать условнорефлекторную деятельность странного животного и попытаться таким образом, может быть, разъяснить ближайший нервный механизм ее поведения. Условнорефлекторная деятельность оказалась налицо. Условные рефлексы были пищевые. Очень легко и скоро восстановился давний рефлекс на звонок. После того был образован новый звуковой рефлекс на тон органной трубы в 300 колебаний в секунду. За ним был получен рефлекс на камфору. Точно так же выработалась дифференцировка к тону из терции и условный тормоз из ударов метронома к рефлексу на камфору. Но первому исследователю больной собаки (Сатурнову) не удалось восстановить ранних (бывших до заболевания) кожно-механических условных рефлексов. При этом исследовании было отмечено, что положительные условные раздражители очень наклонны переходить в тормозные и что тормозной процесс очень инертен, очень долго давая себя знать на положительных раздражителях.
Следующий исследователь собаки (Кураев), оценив только что указанную особенность собаки работая главным образом с коротким изолированным условным раздражением, без труда получил и условные кожно-механические рефлексы. Но тормозная тенденция нервной системы все нарастала вместе с повторением и усилением судорожных припадков, пока сильные припадки 9 мая 1912 г., продолжавшиес небольшими паузами почти 12 часов, не убили собаку.
Вскрытие мозга показало следующее. По снятии рубца дефект мозгового вещества захватил следующие извилины: заднюю часть g. postcruciatus, передние отделы g. suprasplenialis, entolateralis и ectolateralis, g. suprasylvius medius, верхнюю половину g. coronarius и отчасти g. ectosylvius medius. Кроме того, оказались резко атрофированными затылочные и височные доли. Размер мозга здесь уменьшен, и мозговые извилины уплощены. Передние доли полушарий, повидимому, остались совершенно неизмененными.
Как понимать с точки зрения этих нарушений отклонения от нормы в поведении собаки? Вскрытие собаки показало, что разрушение и нарушение полушарий захватило главным образом заднюю половину больших полушарий, только слегка коснувшись передней. Таким образом собака должна была быть подобной собаке без задней половины полушарий, что и сказывалось на условных рефлексах. К сожалению, мы не исследовали ее относительно комплексных звуковых и световых раздражителей. На их отсутствие указывало то, что собака не реагировала на кличку и не обнаруживала специальных отношений ни к другим собакам, ни к людям. Последнее могло зависеть также и от отсутствия у такой собаки цепных рефлексов. Далее некоторая неисправность в ходьбе, очевидно, зависела от небольшого повреждения двигательной области. Представляются трудными для удовлетворительного объяснения стояние собаки на одном месте и чрезвычайная реакция на прикосновение к коже. Что касается первого, то относить ли это на счет несомненного преобладания тормозного процесса в коре (под влиянием повторяющихся взрывов возбуждения) или рассматривать это как результат некоторого частичного повреждения кожного анализатора, тем более, что голова и шея сохранили свою обычную подвижность? Вместе с тем остается непонятным действие прикосновения к коже собаки. Корковый ли это рефлекс или низших отделов мозга? Раз условные кожно-механические рефлексы существуют, последнее маловероятно. Но тогда как согласить повышенную возбудимость кожного анализатора c преобладанием тормозного процесса в коре? Для решения этих вопросов требовались многие вариации опытов, которые в свое время не были сделаны.
Приводя описанные в этой лекции случаи, я отнюдь не имел претензии дать удовлетворительные объяснения нервного механизма всех отклонений от нормы в поведении собак после мозговых операций. Моя цель состояла только в том, чтобы поставить перед физиологией полушарий вопросы относительно этого механизма как законные и показать некоторую возможность решения их.
В целом приведенные в последних трех лекциях опыты с условными рефлексами главным образом являются подтверждением ранних и позднейших исследований других авторов относительно того же предмета, только сольшими прибавлениями с нашей стороны и с постановкой некоторых новых задач в этой области. Но наши факты решительно противоречат учению об отдельных ассоциационных центрах вли вообще о какомто особом отделе полушарий с верховной нервной функцией, против чего восставал уже и Мунк.
Лекция двадцать вторая. Общая характеристика данного исследованиия: его задача, его трудности и наши ошибки
Мм. гг.! При научном изучении животных явлений есть несколько плоскостей, на которых можно вести это изучение. Можно иметь в виду непременную физико-химическую основу жизненных явлений и методами физики и химии анализировать элементарное жизненное явление. Дальше, считаясь как с фактом с эволюцией живого вещества, можно стараться свести деятельность сложных конструкций живого вещества на свойства элементарных форм его. Наконец, охватывая деятельность сложных конструкций во всем их действительном объеме, можно отыскивать строгие правила этой деятельности или, что то же, констатировать все те условия, которые точно определяют течение деятельности во всех ее моментах и вариациях. Плоскость, на которой мы стояли в описанном исследовании, есть, очевидно, последняя плоскость. На этой плоскости наше внимание не занимал вопрос о том: что такое раздражение и торможение в их последнем глубоком основании? Мы брали их как два фактических данных, два основных свойства нашей сложной конструкции, два главнейших проявления деятельности этой конструкции. Мы не ставили себе задачей деятельность полушарий свести на элементарные свойства нервной ткани, как они установлены для нервного волокна. Мы даже не останавливались подробно на вопросе о возможном размещении двух основных явлений этой деятельности -- раздражения и торможения - по двум элементам нашей конструкции: нервным клеткам и соединительным пунктам или путям между нервными клетками, удовлетворяясь временным предположением, что то и другое есть функции нервных клеток. Что изучение условных рефлексов есть изучение деятельности клеток коры больших полушарий, конечно, не может подлежать сомнению. За это бесспорно говорит огромнейшая масса отдельных раздражений, воспринимаемых полушариями как из внешнего, так и из внутреннего мира, для которых соответственно изолированные пункты представлены сотнями миллионов и миллиардами клеток коры иорые отпадают вместе удалением больших полушарий. А затем является вероятным, что раздражение и торможение, так постоянно и тесно переплетающиеся между собой, беспрерывно сменяющие друг друга, суть функции нервных клеток, представляя собой только разные фазы физико-химического процесса, происходящего в этих клетках под влиянием многочисленных раздражений, поступающих в них как из внешнего, так и из внутреннего мира, как из всех отделов тела, так и специально из других пунктов полушарий. Наша основная задача: регистрация и характеристика отдельных актов деятельности коры, определение точных условий их наличности и их систематизация, иначе говоря, определение условий колебаний величины раздражительного и тормозного процессов и установление взаимных отношений этих процессов. Таким образом наша работа по своему характеру аналогична работе Шеррингтона и его сотрудников над спинным мозгом. Нельзя не видеть очень много пунктов, где наши факты, полученные при исследовании больших полушарий, совпадают с фактами физиологии спинного мозга, что свидетельствует об естественной общности основных отношений и там и здесь. Но работа над деятельностью больших полушарий и в тех границах, которые мы себе поставили, является исключительно трудной. В силу чрезвычайной реактивности коры полушарий и грандиозной массы раздражений, постоянно на нее падающих, деятельность ее характеризуют две основные черты: чрезвычайная обусловленность и естественно с ней связанная текучесть явлений, составляющих эту деятельность. Почти ни при одном явлении нельзя быть уверенным в обладании всеми условиями его существования. Малейшее колебание внешней среды или внутреннего мира, часто едва уловимое или совершенно не подозреваемое, резко меняет ход явлений. Понятно, что при таком положении дела здесь в особенности горько дают себя знать обычные слабости мысли: стереотипность и предвзятость. Мысль не может, так сказать, угнаться за разнообразием отношений. Поэтому так часто приходится ошибаться при этой работе. Я уверен, что и в переданном материале немало промахов, и даже больших. Но, посягая на такую сложность, не стыдно и ошибаться. Вот почему я так долго медлил с систематическим изложением нашей столь продолжительной работы. В силу указанных свойств предмета постоянно возникают новые вопросы, а позади остается еще много нерешенных вопросов. От намеченных задач нас часто отвлекает неожиданное и нередко смущает многое непонятное с принятой нами точки зрения. Эту общую характеристику работы над большими полушариями я и хочу дать в этой заключительной лекции, опираясь опять же на наш фактический материал, не нашедший себе места в предшествующих лекциях. До какой степени явления в исследуемой нами области сложны и как они тонко и многообразно обусловлены, я думаю, особенно ярко показывают два следующих наблюдения, относяшиеся к последнему периоду нашей работы. Вот собака, о которой была не раз речь в предшествующих лекциях, подвергшаяся сильному влиянию чрезвычайного наводнения и представившая после применения определенного нашего приема изолированное функциональное поражение в звуковом анализаторе. Целый месяц сплошь при решении некоторой задачи, именно при выработке дифференцировки на высоту тона, она держалась вполне нормально. При продолжительности изолированного условного раздражения только в 10 секунд секреторный эффект достигал 5 капель. Отчетливо выступала разница между сильными и слабыми раздражителями. Подаваемую после условных раздражителей пищу собака брала сразу и ела жадно. В станке стояла спокойно. Теперь вводится, повидимому, очень маленькое изменение в опыте. Изолированное условное раздражение удлиняется на 5 секунд. И сейчас же вся условнорефлекторная деятельность резко нарушается. Вот подлинные цифры опыта.
 Первый раздражитель во втором опыте дает секреторный эффект больший, чем обычный, что и понятно, так как изолированное условное раздражение удлинено на 5 секунд. Пищевая реакция на условный раздражитель - живая, и пища сейчас же съедается. Все обещает обыкновенный опыт. Никаких указаний на какую-нибудь экстренную ненормальность этого дня нет. Но уже в паузе обращает на себя внимание какое-то беспокойство животного. А затем выступает явная парадоксальная фаза. На сильные раздражители секреторная реакция исчезает, и подаваемую пищу собака в первый раз берет с опозданием, а во второй раз и совсем не берет. На слабый же раздражитель секреция есть, хотя и уменьшенная, и пища берется сразу. На другой день при обычной продолжительности условных раздражений в 10 секунд положение дела еще более ухудшается. На все раздражители секреторный эффект отсутствует, пищу при сильных условных раздражителях собака не берет, при слабом ест. На третий день все вернулось к норме, лишь тон, примененный первым в день с удлинением изолированного условного раздражения, имел ослабленный секреторный эффект, вдвое против нормальной величины. Опыт был повторен с тем же результатом. Какая тонкость реагирования, какое значительное действие от ничтожного изменения условий опыта!
Но дело не ограничивается одной реактивностью. На другой собаке, тоже упоминавшейся раньше, исключительно тормозимой (Умнице), такое же изменение условий опыта сопровождалось обратным результатом. Так как эта собака на станке при спокойной обстановке скоро делалась сонливой и все рефлексы не только на условные раздражители, но и на подаваемую пищу совершенно отсутствовали, то был применен испытанный возбуждающий прием: кратчайший переход (0.5-1 секунда) от начала действия условного раздражителя к подаче пищи. Через 3 недели практики такого приема у собаки рассеялась сонливость, и она всякий раз брала подаваемую пищу и жадно ее ела. Когда затем изолированное условное раздражение было удлинено на 5 5 секунд, оказывался и условный секреторный эффект. Но он держался несколько дней, а затем исчезал, и собака опять становилась сонливой. Стоило, однако, удлинить условные раздражения до 10 секунд - снова собака делалась бодрее, и опять появлялся секреторный эффект, и - что существенно - он был уже и в первые 5 секунд. Далее повторялась старая история. После немногих дней собака впадала в прежнюю сонливость, и секреторного условного рефлекса более нело. Удлинение условного раздражения до 15 секунд имело то же действие, что первое удлинение до 10 секунд: бодрое состояние собаки и значительный условный секреторный эффект, начинавшийся уже в первые 5 секунд. Так же шло дело и при удлинении условного раздражителя до 20 и 25 секунд.
Привожу протокол опытов.
Первый раздражитель во втором опыте дает секреторный эффект больший, чем обычный, что и понятно, так как изолированное условное раздражение удлинено на 5 секунд. Пищевая реакция на условный раздражитель - живая, и пища сейчас же съедается. Все обещает обыкновенный опыт. Никаких указаний на какую-нибудь экстренную ненормальность этого дня нет. Но уже в паузе обращает на себя внимание какое-то беспокойство животного. А затем выступает явная парадоксальная фаза. На сильные раздражители секреторная реакция исчезает, и подаваемую пищу собака в первый раз берет с опозданием, а во второй раз и совсем не берет. На слабый же раздражитель секреция есть, хотя и уменьшенная, и пища берется сразу. На другой день при обычной продолжительности условных раздражений в 10 секунд положение дела еще более ухудшается. На все раздражители секреторный эффект отсутствует, пищу при сильных условных раздражителях собака не берет, при слабом ест. На третий день все вернулось к норме, лишь тон, примененный первым в день с удлинением изолированного условного раздражения, имел ослабленный секреторный эффект, вдвое против нормальной величины. Опыт был повторен с тем же результатом. Какая тонкость реагирования, какое значительное действие от ничтожного изменения условий опыта!
Но дело не ограничивается одной реактивностью. На другой собаке, тоже упоминавшейся раньше, исключительно тормозимой (Умнице), такое же изменение условий опыта сопровождалось обратным результатом. Так как эта собака на станке при спокойной обстановке скоро делалась сонливой и все рефлексы не только на условные раздражители, но и на подаваемую пищу совершенно отсутствовали, то был применен испытанный возбуждающий прием: кратчайший переход (0.5-1 секунда) от начала действия условного раздражителя к подаче пищи. Через 3 недели практики такого приема у собаки рассеялась сонливость, и она всякий раз брала подаваемую пищу и жадно ее ела. Когда затем изолированное условное раздражение было удлинено на 5 5 секунд, оказывался и условный секреторный эффект. Но он держался несколько дней, а затем исчезал, и собака опять становилась сонливой. Стоило, однако, удлинить условные раздражения до 10 секунд - снова собака делалась бодрее, и опять появлялся секреторный эффект, и - что существенно - он был уже и в первые 5 секунд. Далее повторялась старая история. После немногих дней собака впадала в прежнюю сонливость, и секреторного условного рефлекса более нело. Удлинение условного раздражения до 15 секунд имело то же действие, что первое удлинение до 10 секунд: бодрое состояние собаки и значительный условный секреторный эффект, начинавшийся уже в первые 5 секунд. Так же шло дело и при удлинении условного раздражителя до 20 и 25 секунд.
Привожу протокол опытов.
 в следующем опытном дне условные раздравения удлиняются до 20 секунд.
в следующем опытном дне условные раздравения удлиняются до 20 секунд.
 Таким образом каждое небольшое удлинение изолированного условного раздражения производило временное возбуждение собаки: сонливое состояние рассеивалось, и секреторный условный эффект появлялся в такой близкий к началу условного раздражения момент, в какой его не было до этого удлинения. Следовательно, он был результатом возбуждения коры, а не следствие только увеличения времени до его обнаружения.
Итак, на двух собаках одно и то же совершенно незначительное изменение в опыте обусловило совершенно противоположный эффект. У одной оно вызвало торможение, у другой, наоборот, оно устраняло торможение. Надо думать, основание этои противоположности состояло в том, что первое животное находилось в бодром (возбужденном) состоянии, второе в сонном (заторможенном).
Привожу пример, где, очевидно, основное отношение явлений испытывает отклонение вследствие влияние пока неизвестных второстепенных условий. Как уже многократно указывалось в предшествующих лекциях, условные раздражители из внешних агентов, относящихся к разным анализаторам, при всех прочих равных условиях, дают разные по величине эффекты. Долгое время только под влиянием предвзятой мысли, что разница в наших ощущениях света, звука и т. д. должна же иметь какоелибо материальное основание в разнице соответственных клеток коры, раз нервный процесс во всех нервах современной физиологией признается тождественным, мы были наклонны и разницу в величине условного эффекта тоже отнести насчет индивидуальности клеток разных анализаторов. Но некоторые факты уже в течение этих лекций потребовали от нас нарочитого экспериментального исследования этого пункта. При этом оказалось, как уже было сообщено ранее, что разница в величине условного эффекта при агентах из разных анализаторов несомненно существенно зависит от количества энергии, посылаемой каждым агентом в кору полушарий. Как помните, этот результат был получен следующим образом: мы давно знали, что условный комплексный раздражитель, состоящий из двух обычно применямых нами агентов и принадлежащих к разным анализаторам, действует преимущественно или даже почти исключительно одним из компонентов, как показывает испытание условного действия каждого из компонентов в отдельности, например наши обычные звуковые агенты всегда в комплексе замаскировывали световые, кожно-механический, кожно-температурный и т. д. Но, нарочито чрезвычайно ослабляя звуковой агент и усиливая световой, мы получили обратное отношение, чем и было доказано определяющее значение в факте количества энергии. Однако основной факт связи величины условного эффекта с количеством энергии иногда, хотя и не часто, у некоторых животных не обнаруживается, и мы до сих пор не можем определенно указать специальные условия этих отклонений, хотя и направляем усиленное внимание на этот пункт. Отдельные условия более или менее намечаются, но вся совокупность их остается незахваченной. Отклонения, видимо, связаны с общим характером нервной системы: возбудимым или тормозимым. У тормозных собак обычная связь величины эффекта с силой раздражения выступает особенно резко и почти без исключения, не считая явно патологических состояний. Затем определенное значение имеет продолжительность изолированного условного раздражения. Если эта продолжительность не велика, разница может не оказаться. Начинаясь одинаково при сильных и слабых раздражениях, эффект при слабых раздражениях обычно не так сильно нарастает, а иногда и совсем не увеличивается в течение раздражения, как это почти всегда происходит при сильных раздражениях. Но у очень возбудимых и жадных до еды собак, наоборот, укорачивание изолированного условного раздражения способствует обнаружению обычной зависимости величины эффекта от силы раздражения, которая незаметна при более продолжительном условном раздражении.
Опять же, вследствие чрезвычайной обусловленности явлений, составляющих деятельность больших полушарий, и при многих, повидимому, простых и, казалось бы, хорошо обследованных фактах приходится чувствовать все еще неудовлетворительность нашего теперешнего физиологического анализа. Возьмем один из наших недавних случаев (опыты Н. А. Подкопаева и С. Н. Выржиковского). У собаки образовывались условные рефлексы на разные агенты при следующей вариации условий. Первый индифферентный агент один раз применялся в сопровождении безусловного раздражителя), в другой раз один, - и это повторялось несколько раз. Условный рефлекс образовался довольно скоро (на двадцатый раз). При следуюшем агенте пицца давалась при одном разе из трех. Рефлекс тоже образовался, даже скорее (на седьмом разе), но собака при этом становилась очень возбужденной. Наконец агент, за которым еда следовала только один раз при четырех его повторениях,совсем не сделался условным раздражителем, причем собака делалась несколько сонливой. При этом агент в целом был применен 240 раз, из них только 60 раз в сопровождении еды. Попробуем, пользуясь нашими другими основными фактами, понять механизм этого факта: почему условный рефлекс не образовался в данном случае или по крайней мере его образование так замедлялось, если допустить, что при еще большем числе он мог бы образоваться? Основной механизм образования условного рефлекса есть встреча, совпадение во времени раздражения определенного пункта коры полушарий с более сильным раздражением другого пункта, вероятно коры же, в силу чего между этими пунктами более или менее скоро проторивается более легкий путь, образуется соединение. И обратно, если эта встреча долго не повторяется, проторенность исчезает, соединение разрывается. Но когда проторенность дошла до ее максимума, она и без дальнейшей практики может держаться месяцы и годы. Понятно, что при соответствующем условии, т. е. при встрече раздражений, проторенность должна образовываться при первой встрече и постоянно увеличиваться, суммироваться при следующих встречах. Тогда является законный вопрос: почему же условный рефлекс не получился в последней вариации описываемого опыта? Ведь встреча происходила 60 раз, когда при обыкновенных условиях достаточно 3-5 раз в лучшем случае, а в данном опыте при первой вариации он образовался после 20 раз, что обыкновенно и бывает при образовании первых условных рефлексов у собаки при нашей обстановке. Конечно, прежде всего приходит мысль о влиянии промежутка времени между повторениями встречи. Но это не имеет значения в нашем случае. При тех же промежутках, если бы промежуток не был заполнен повторением индифферентного раздражителя без солровождения едой, условный рефлекс несомненно бы образовался. Значит, помеха не в этом. Неизбежно принимать, что именно частое повторение индифферентного агента без условного раздражителя создало препятствие для образования условного рефлекса. Как представить себе это препятствие? Как мы уже знаем, всякий новый раздражитель, вызывающий сначала исследовательский рефлекс, перестает вызывать его. если затем этим раздражителем не производится на животное никакого дальнейшего действия. И это прекращение действия основано на развитни торможения в клетке, куда адресовалось раздражение. Следовательно, при трех повторениях агента в третьей вариации без присоединения еды и могло происходить это торможение соответствующей клетки, и, таким образом, для соединения с клеткой пищевого раздражения в ней не было процесса раздражения. Предположение кажется достаточно обоснованным, но оно не может быть признано в данном случае за реализованное. Когда агент в третьей вариации был повторен 240 раз, он был испытан относительно его свойства. На очень коротком расстоянии времени после него (30 секунд) применялся какойнибудь из бывших у собаки условных рефлексов - он отнюдь не оказывался заторможенным. Следовательно, агент не был тормозным и не производил последовательного торможения. Но его тормозное действие должно быть очень концентрированным вследствие долгого употребления. И это не подтвердилось. Когда после 240 раз этот агент начали сопровождать едой сплошь, он уже на третий раз дал значительновный секреторный эффект. Нужно еще прибавить, что в первый день, когда агент был применен в комбинации с пищей два раза, ранние условные рефлексы были малы, т. е. возбудимость собаки была невысока. Таким образом скорость образования нового условного рефлекса была максимальной, и думать о тормозном состоянии клетки индифферентного агента нет основания. Мы испытали предположения, истекающие из известных нам фактов, и причину явления не нашли. Мы будем испытывать ее дальше и, надо думать, найдем. Но сделанное до сих пор показывает, что у нас еще нет сейчас полного знания всех тех условий, которые определяют деятельность механизма образования условного рефлекса. Приведенные во второй лекции условия, как они ни были достаточны для получения всех рефлексов, бывших до сих пор в нашем распоряжении, значит, еще не полны, раз, имея их в голове, мы не можем сразу понять результата анализируемого случая. Есть еще какое-то просматриваемое нами условие. Значит, опять чрезвычайная обусловленность явлений.
Если не может быть речи в настоящее время о понимании всей деятельности больших полушарий из деталей физико-химического процесса, что очевидно; если нет, по всей вероятности, верных шансов для понимания этой деятельности из элементарных свойств нервной ткани; если мы даже относительно самого основного явления этой деятельности не имеем еще полного знания, то что же остается для нашего исследования, к чему сводится наша работа? Очевидно, приведение бесчисленной массы разнообразных отдельных явлений все к меньшему числу общих основных явлений и должно составить ближайшую и реальнейшую задачу теперешней стадии физиологического изучения больших полушарий. Этим мы и занимаемся теперь, то приближаясь несколько к цели в отдельных случаях, то стоя перед неразрешенными, но атакуемыми вопросами, то ошибаясь.
В начале этой работы, опираясь на резкие, но внешние черты, мы различали три вида торможения: внешнее, внутреннее и сонное. Накопляющиися фактический материал позволил нам, как вы уже знаете, слить последние два вида, сведя их внешнюю разницу на второстепенную подробность. Оба они оказались одним и тем же внутренним торможением, раз раздробленным и местно разграниченным, в другой раз слитым и широко распространенным. Когда при дальнейшем изучении условных рефлексов мы познакомились с явлениями взаимной индукции, естественно. бросилось в глаза подобие факта отрицательной индукции с фактом внешнего торможения. Теперь представилась вероятной тождественность в основе процесса при всех наших старых видах торможения, почему мы и приступили к собиранию дальнейших данных в этом направлении. Эти данные частью были сообщены раньше, частью я их приведу теперь. В лекции об индукции был описан опыт, в котором условный оборонительный рефлекс на механическое раздражение кожи на одном определенном месте тормозил в периоде последействия условный пищевой рефлекс на механическое раздражение кожи на других местах. При этом одна подробность свидетельствовала, что это торможение происходило обоими пунктами (пунктами раздражения и торможения), именно в коре полушарий. В лекции о переходных фазах к полному внутреннему торможению было указано на некоторых примерах, что и в периоде внешнего торможения можно наблюдать те же фазы. Сейчас я могу прибавить новые соображения и факты в пользу заключения о тождестве внешнего и внутреннего торможения. В вышеупомянутом факте замаскирования, подавления в комплексном условном раздражителе сильным раздражителем слабого необходимо видеть обнаружение внешнего торможения. Клетка сильного раздражителя тормозит клетку слабого раздражителя, и потому эта последняя клетка может войти только в слабую связь с клеткой безусловного раздражителя, так как есть основание считать чтопень проторенности соединительного пути отвечающей силе условного раздражителя. А чем бы в данном случае от этого внешнего торможения отличалась отрицательная индукция, когда пункт положительного условного раздражителя усиливает или даже восстановляет тормозное состояние пункта условного тормозного раздражителя? Наконец к доказательству устанавливаемого нами положения могут служить и следующие факты. Уже давно многими нашими сотрудниками (Минтовт, Кржышковский, Лепорский) указывалось, что при образовании условного тормоза для быстрого и полного торможения имеет большое значение, чтобы агент, из которого будет вырабатываться условный тормоз, не был значительно физически слабее, чем агент, являющийся положительным условным раздражителем, участником тормозной комбинации. Недавние нарочитые опыты (Фурсикова) вполне подтвердили эти указания, прибавив факт, что часто внешнее торможение, с которого начинается действие нового агента, как вызывающего исследовательский рефлекс, незаметно переходит в тормозное действие комбинации, как внутреннее торможение. И все же остается вопрос: достаточно ли это для отождествления внешнего и внутреннего торможения?
В девятнадцатой лекции был поднят вопрос о трех видах внешних раздражителей, вызывающих прямо тормозное состояние коры больших полушарий: очень слабых, очень сильных и необыкновенных, причем было предположительно указано биологическое значение этого факта. Вопрос о физиологическом механизме действия этих различных раздражителей был отложен до настоящей лекции. Но и здесь сейчас я не нахожу возможным ставить и обсуждать вопрос о торможении во всем его объеме. Как ни значителен наш экспериментальный материал, он явно недостаточен, чтобы составить общее и определенное представление о торможении и об его отношении к раздражению. Объяснения, хорошо подходящие к одной группе фактов, не могут, однако, захватить их все. Многое факты упорно не поддаются теоретическому анализу, и представление об их механизме приходилось несколько раз менять на протяжении нашей работы, не находя его удовлетворительным. И тут, как и вообще в нашей работе, пока остается только группировать фактический материал. Мы стоим перед массой нерешенных вопросов: о растормаживании, положительном действии тормозных условных раздражителей, в некоторых случаях о прямо тормозящем действии положительных условных раздражителей при нарушении коры полушарий, о преимущественном тормозящем действии слабых и сильных положительных раздражителей, и т. д. и т. д. Мы во многих случаях не можем сказать: какие из фактов близки или тождественны, или же совершенно отдельны, радикально различны. Для иллюстрации трудности таких случаев рассмотрю следующий пример. Почему новые явления или новая связь явлений действует на животное тормозящим образом? Какой физиологический механизм этого задерживания? Мы подаем собаке после условного раздражителя корм другим способом, чем раньше, например вместо чашки с пищей, выдвигаемой из-за экрана, сбрасываем ее через трубку из резервуара, стоящего наверху, в чашку, находящуюся перед собакой, и многие из собак тогда ее упорно не берет, и наши условные рефлексы исчезают. Наступает, конечно, торможение. На что его свести, с каким из наших других фактов его поставить рядом? Может быть, с фактом, упомянутым в тринадцатой лекции, когда при резком изменении обычного порядка в применении ряда различных условных раздражителей в течение опытов наступает, особенно у некоторых собак, более или менее значительное торможение всей условнорефлекторной деятельности, держащееся несколько дней, несмотря даже на возвращение к старому порядку? Можно представить себе то же самое и относительно всей окружающей собаку действительности. Постоянно повторяющиеся и в определенном следовании перед животным внешние явления создают в полушариях определенную, так сказать, стереотипную деятельность. Всякое новое явление, или даже и старые, но в новой связи, нарушая этим стереотип, вызывают торможение, как и в наших опытах с изменением порядка условных раздражителей. Но тогда встает следующий вопрос: как, каким ближайшим физиологическим процессом происходит торможение в последнем случае, или, правильнее сказать, с каким в свою очередь другим случаем торможения тождествен этот случай торможения? Не есть ли это результат тормозящего действия исследовательского рефлекса, возникающего при новом колебании внешней среды? Или же это совершенно отдельные, самостоятельные явления? За первое бы говорила резко выраженная у очень тормозимых собак связь чрезвычайно чувствительного ориентировочного рефлекса с сильным последовательным торможением (факт, выступавший исключительно отчетливо y Sмницы), и при таких собаках такой механизм является вероятным. Но в других случаях нет внешних указаний на ориентировочный рефлекс и наступает прямо тормозное состояние.
Этот пример показывает, как мы еще далеки от удовлетворительного группирования наших отдельных фактов.
Вот почему я пока отказываюсь высказываться за ту или другую из существующих теорий торможения или выставлять нобую. Пока мы пользуемся временными предположениями для систематизирования нашего фактического материала и проектырования новых опытов.
В заключение я перехожу к нашим ошибкам, которых мы делали немало в прошлом и которые постепенно поправляли и поправляем. Сейчас сперва опишу одну, которая была открыта и исправлена при повторении старых опытов в течение этих лекции, и другую, вероятную, которую исследуем в настоящее время.
В четвертой лекции, в которой говорилось о восстановлении угашенного условного рефлекса, были приведены три различных случая этого восстановления: самостоятельное по истечении известного срока, прочное, но более или менее медленно наступающее, и ускоренное, раз посредством подкрепления, т. е. применения специального рефлекса, в другой раз вследствие применения каких-нибудь посторонних рефлексов. Эти два последние случая описаны там как существенно различные между собой. При первом из них принималось не только быстрое, но и прочное восстановление. При втором случае с посторонними рефлексами было установлено, что восстановление хотя и быстро наступало, но было временным и скоро исчезало по прекращении постороннего рефлекса и его последействия, сменяясь опять торможением, продолжавшимся до конца его обычного срока, как если бы никакого постороннего рефлекса и не было. Поэтому это восстановление получило особенное название «растормаживания». При этом там было указано на трудность понимать эту разницу, раз угасание не было окончательным разрушением условного рефлекса. В последнее время при получении новых данных относительно взаимодействия условного и безусловного раздражителей мы сочли надобным переделать возможно точно старые опыты и убедились в ошибочности их (новый опыт Подкопаева). Восстановление угашенного рефлекса при применении как специального, так и всякого постороннего рефлекса только временное, т. е. после обоих рефлексов в ближайшее время угашенный условный раздражитель действует положительно, а несколько позже опять лишается этого действия до его самостоятельного восстановления. Когда условный рефлекс - пищевой, а посторонний - кислотный, т. е. оба химические, то временное восстановление, растормаживание происходит совершенно однообразно, что касается его течения во времени. Опыты были поставлены на двух собаках и дали один и тот же результат. Даю подробное описание опыта на одной из собак. При угасании условного пищевого рефлекса на удары метронома до нуля этот нуль держался до 20 минут после последнего повторения условного раздражителя без подкрепления. Затем начиналось постепенное самостоятельное восстановление, которое к 30 минутам достигало 40 0/ % первоначальной величины условного эффекта до угасания. Когда при первом нуле угасания условный раздражитель сейчас же подкреплялся и через 20 минут от нуля он испытывался опять, то оставался еще нулевым. Когда же в другом опыте после такого же подкрепления он применялся спустя только 10 10 минут после угасания, от него имелся положительный эффект. Совершенно так же шел опыт, когда вместо пищи в тот же момент вливалась в рот собаки кислота и в те же сроки испытывался условный раздражитель.
Вот подлинные цифры, взятые из некоторых опытов для примера.
Условный раздражитель при первом применении в опытный день дал 6 капель за 20 секунд; он же после конца угасания, тотчас же подкрепленный и испытанный 10 минут спустя, считая от конца угасания, дал 3 капли.
В другой день условный раздражитель при первом применении дал 7 капель; он же после конца угасания, тотчас же подкрепленный и испытанный 20 минут спустя, считая от конца угасания, остался без малейшего эффекта.
Ус Условный раздражитель при первом применении в опытный день дал 5 капель за 20 секунд; он же после конца угасания тотчас же сопровождался вливанием кислоты в рот собаки и испытанный 10 минут спустя, считая от конца угасания, дал 2 капли.
В другой день условный раздражитель при первом применении дал 5 капель; он же после конца угасания, тотчас же сопровождаемый вливанием кислоты и испытанный 20 минут спустя, считая от конца угасания, остался без малейшего эффекта.
Максимум растормаживания наступает в более ранние сроки после конца угасания, чем 10 минут.
Очевидно, прежняя ошибка была основана частью на неточности опытов, когда сравнение сроков восстановления при специальном рефлексе и посторонних рефлексах чаще всего происходило при применении таких посторонних рефлексов, как звуковые, зрительные и другие, имевших короткое последействие, а не химических с их длинным последействием. Частью же к старому заключению располагала мысль, что безусловный раздражитель, с которым сочетался условный, должен находиться в особенном отношении к последнему в случае его восстановления после угасания. Теперешний же результат представляет лишнее основание для принятия, что торможение происходит именно в нервной клетке, а не в соединительном пункте или пути между клеткой условного раздражителя и клеткой специального безусловного раздражителя. Иначе было бы трудно себе представить, как восстановляющее действие обоих этих раздражителей могло оказаться совершенно одинаковым.
Другая ошибка обнаруживается сейчас, и я нахожу возможным говорить о ней сейчас, хотя переисследование предмета только начинается, с одной стороны, ввиду особенной важности пункта, к которому она относится, с другой - опять же ради общей характеристики нашей области, изобилующей исключительными трудностями при установлении точных фактических отношений.
Во второй лекции приведены были, казалось, все условия, при которых образуются условные рефлексы, и все, что мешает их образованию. На втором месте после первого условия совпадения во времени индифферентного агента и безусловного раздражителя - было поставлено непременное, хотя бы и очень короткое, предшествование по времени будущего условного раздражителя перед безусловным. На многих вобаках после многочисленных повторений (даже 300-400 раз) комбинации индифферентного агента с безусловным раздражителем, причем комбинация начиналась с безусловного раздражителя и к нему индифферентный агент присоединялся только спустя 5-10 секунд, условный рефлекс не вырабатывался, хотя у тех же собак при других разных агентах, сочетаемых обычным образом с безусловным раздражителем, условные рефлекса были готовы через 7-20 повторений. Естественно было думать, что сильный безусловный раздражитель, сосредоточенный в определенном месте полушарий механизмом внешнего торможения, обусловливал такое тормозное состояние в остальных частях полушарий, что раздражения, падающие на эти части, оказывались недействительными, совершенно так, как это мы знаем по себе, когда, чем-нибудь сильно занятые, мы не слышим и не видим, что происходит около нас. Таким образом факт казался очевидным и понятным как с нашей точки зрения, так и с общежитейской. Это и сделало то, что до самого последнего времени у нас не было относительно него ни малейшего сомнения. Теперь положение дела изменилось. Когда мы недавно задались вопросом: каким образом раннее подкрепление условного раздражения, т. е. возможное укорочение его изолированного действия, мешает развитию тормозного состояния в пункте его приложения в коре, то новым варьированием опытов были приведены к неожиданному для нас факту. Оказалось, что безусловный раздражитель тормозит и выработанный условный раздражитель. Но тогда естественно и обратило на себя внимание следующее сопоставление фактов.
Имеется ли уже выработанный условный раздражитель, или это еще индифферентный агент, из которого будет только вырабал батываться этот условный раздражитель, - влияние на него безусловного раздражителя одно и то же. А вместе с тем влияние это выражается в том, что при малейшем изменении внешних условий, условий времени, происходит образование условного рефлекса, превращение индифферентного агента в определенный условный раздражитель. Приходилось признать, что механизм образования условного рефлекса и механизм внешнего торможения как-то сближаются или даже отождествляются, иначе говоря, что процесс внешнего торможения дает основание для образования связи между клетками. Это напоминало бы другой из наших фактов, именно образование вторичного условного рефлекса и развитие условного торможения, где также при общих внешних условиях, но при небольшой разнице в отношении времени, наступает то процесс раздражения, то процесс торможения. Если бы это соображение было верно, то надо было бы ждать, что и при покрытии, как мы выражаемся, с самого начала безусловным раздражителем индифферентного агента между пунктами их в коре вначале могла бы образоваться связь, мог бы произойти условный рефлекс, но который потом при повторении такой комбинации затормаживался. И это показали наши первые ориентировочные опыты. Опираясь на то, что полное покрытие выработанных условных раздражителей безусловным только постепенно ведет к уменьшению их эффекта, при слабых к более скорому и даже к нулю, а при сильных к более медленному и часто к незначительному, мы при новых комбинациях безусловного раздражителя с индифферентными агентами, - причем комбинация начиналась с безусловного раздражителя, - применяли эту комбинацию только малое число раз, чтобы не дать развиться торможению. Во многих случаях мы действительно получили то, что ожидали. После таких комбинаций индифферентный агент, испытанный отдельно, действовал как условный раздражитель (в опытах В. И. Павловой, Крепса, Подкопаева и Купалова). Когда мы с этим взглядом на дело и с первыми опытами в этом направлении обратились к нашим старым опытам (опыты Крестовникова), мы увидали, что, во-первых, почти всегда при них индифферентные агенты испытывались на их условное действие только после большого числа повторений комбинаций, в которых они присоединялись к безусловному раздражителю после начала его действия, а, вовторых, даже и при этом часто сильные индифферентные агенты при отдельной их пробе долгобнаруживали секреторный эффект. Но этот эффект истолковывался как случайный и посторонние, а не как условный, в силу предвзятой мысли, что, если бы он был действительно условным, он должен был бы c повторением комбинации увеличиваться, не уменьшаться и исчезать, как это было в тех опытах. Возможно, что и другое явление, именно отсутствие при пробах ясной условной двигательной реакции, при этой предвзятости тоже не вполне объективно оценивалось. На одной собаке должно было быть признано образование подлинного условного рефлекса, но оно также было сведено на постороннее условие, конечно с известным основанием, - но вполне ли достаточным? Тема будет обработана многосторонне и строго при теперешнем очень увеличенном запасе наших сведений о предмете и при большей специальной тренировке нашей мысли в новой области исследования. При полном оправдании наших соображений и при дальнейшем подтверждении наших теперешних ориентировочных опытов физиология больших полушарий овладела бы очень важным для приложения к человеку положением, что образование новых связей в коре может происходить не только в районах полушарий с оптимальной возбудимостью, но и в частях их, более или менее заторможенных.
в этой последней, также фактической, лекции я, однако, хотел, как только мог, передать основное впечатление от работы в нашей области. Как мне кажется, многочисленные факты, сообщенные во всех предшествующих лекциях, достаточно свидетельствуют, что в этой области возможна серьезная естественнонаучная работа, накопление точных фактических данных. Поэтому я не боялся дискредитировать наше научное предприятие тем, что привел в этой лекции. Как всегда, лучше, конечно, сознавать опасности, чем их игнорировать. Кроме того, мне хотелось предупредить будущих работников этой области о тех исключительных трудностях, которые они встретят в ней.
В общем же этот новый отдел физиологии поистине пленителен, удовлетворяя двум, всегда рядом идущим, тенденциям человеческого ума: стремлению к захватыванию все новых и новых истин и протесту против претензий как бы законченного где-нибудь знания. Здесь гора неизвестного явно надолго останется безмерно больше кусочков отторгнутого, познанного.
Таким образом каждое небольшое удлинение изолированного условного раздражения производило временное возбуждение собаки: сонливое состояние рассеивалось, и секреторный условный эффект появлялся в такой близкий к началу условного раздражения момент, в какой его не было до этого удлинения. Следовательно, он был результатом возбуждения коры, а не следствие только увеличения времени до его обнаружения.
Итак, на двух собаках одно и то же совершенно незначительное изменение в опыте обусловило совершенно противоположный эффект. У одной оно вызвало торможение, у другой, наоборот, оно устраняло торможение. Надо думать, основание этои противоположности состояло в том, что первое животное находилось в бодром (возбужденном) состоянии, второе в сонном (заторможенном).
Привожу пример, где, очевидно, основное отношение явлений испытывает отклонение вследствие влияние пока неизвестных второстепенных условий. Как уже многократно указывалось в предшествующих лекциях, условные раздражители из внешних агентов, относящихся к разным анализаторам, при всех прочих равных условиях, дают разные по величине эффекты. Долгое время только под влиянием предвзятой мысли, что разница в наших ощущениях света, звука и т. д. должна же иметь какоелибо материальное основание в разнице соответственных клеток коры, раз нервный процесс во всех нервах современной физиологией признается тождественным, мы были наклонны и разницу в величине условного эффекта тоже отнести насчет индивидуальности клеток разных анализаторов. Но некоторые факты уже в течение этих лекций потребовали от нас нарочитого экспериментального исследования этого пункта. При этом оказалось, как уже было сообщено ранее, что разница в величине условного эффекта при агентах из разных анализаторов несомненно существенно зависит от количества энергии, посылаемой каждым агентом в кору полушарий. Как помните, этот результат был получен следующим образом: мы давно знали, что условный комплексный раздражитель, состоящий из двух обычно применямых нами агентов и принадлежащих к разным анализаторам, действует преимущественно или даже почти исключительно одним из компонентов, как показывает испытание условного действия каждого из компонентов в отдельности, например наши обычные звуковые агенты всегда в комплексе замаскировывали световые, кожно-механический, кожно-температурный и т. д. Но, нарочито чрезвычайно ослабляя звуковой агент и усиливая световой, мы получили обратное отношение, чем и было доказано определяющее значение в факте количества энергии. Однако основной факт связи величины условного эффекта с количеством энергии иногда, хотя и не часто, у некоторых животных не обнаруживается, и мы до сих пор не можем определенно указать специальные условия этих отклонений, хотя и направляем усиленное внимание на этот пункт. Отдельные условия более или менее намечаются, но вся совокупность их остается незахваченной. Отклонения, видимо, связаны с общим характером нервной системы: возбудимым или тормозимым. У тормозных собак обычная связь величины эффекта с силой раздражения выступает особенно резко и почти без исключения, не считая явно патологических состояний. Затем определенное значение имеет продолжительность изолированного условного раздражения. Если эта продолжительность не велика, разница может не оказаться. Начинаясь одинаково при сильных и слабых раздражениях, эффект при слабых раздражениях обычно не так сильно нарастает, а иногда и совсем не увеличивается в течение раздражения, как это почти всегда происходит при сильных раздражениях. Но у очень возбудимых и жадных до еды собак, наоборот, укорачивание изолированного условного раздражения способствует обнаружению обычной зависимости величины эффекта от силы раздражения, которая незаметна при более продолжительном условном раздражении.
Опять же, вследствие чрезвычайной обусловленности явлений, составляющих деятельность больших полушарий, и при многих, повидимому, простых и, казалось бы, хорошо обследованных фактах приходится чувствовать все еще неудовлетворительность нашего теперешнего физиологического анализа. Возьмем один из наших недавних случаев (опыты Н. А. Подкопаева и С. Н. Выржиковского). У собаки образовывались условные рефлексы на разные агенты при следующей вариации условий. Первый индифферентный агент один раз применялся в сопровождении безусловного раздражителя), в другой раз один, - и это повторялось несколько раз. Условный рефлекс образовался довольно скоро (на двадцатый раз). При следуюшем агенте пицца давалась при одном разе из трех. Рефлекс тоже образовался, даже скорее (на седьмом разе), но собака при этом становилась очень возбужденной. Наконец агент, за которым еда следовала только один раз при четырех его повторениях,совсем не сделался условным раздражителем, причем собака делалась несколько сонливой. При этом агент в целом был применен 240 раз, из них только 60 раз в сопровождении еды. Попробуем, пользуясь нашими другими основными фактами, понять механизм этого факта: почему условный рефлекс не образовался в данном случае или по крайней мере его образование так замедлялось, если допустить, что при еще большем числе он мог бы образоваться? Основной механизм образования условного рефлекса есть встреча, совпадение во времени раздражения определенного пункта коры полушарий с более сильным раздражением другого пункта, вероятно коры же, в силу чего между этими пунктами более или менее скоро проторивается более легкий путь, образуется соединение. И обратно, если эта встреча долго не повторяется, проторенность исчезает, соединение разрывается. Но когда проторенность дошла до ее максимума, она и без дальнейшей практики может держаться месяцы и годы. Понятно, что при соответствующем условии, т. е. при встрече раздражений, проторенность должна образовываться при первой встрече и постоянно увеличиваться, суммироваться при следующих встречах. Тогда является законный вопрос: почему же условный рефлекс не получился в последней вариации описываемого опыта? Ведь встреча происходила 60 раз, когда при обыкновенных условиях достаточно 3-5 раз в лучшем случае, а в данном опыте при первой вариации он образовался после 20 раз, что обыкновенно и бывает при образовании первых условных рефлексов у собаки при нашей обстановке. Конечно, прежде всего приходит мысль о влиянии промежутка времени между повторениями встречи. Но это не имеет значения в нашем случае. При тех же промежутках, если бы промежуток не был заполнен повторением индифферентного раздражителя без солровождения едой, условный рефлекс несомненно бы образовался. Значит, помеха не в этом. Неизбежно принимать, что именно частое повторение индифферентного агента без условного раздражителя создало препятствие для образования условного рефлекса. Как представить себе это препятствие? Как мы уже знаем, всякий новый раздражитель, вызывающий сначала исследовательский рефлекс, перестает вызывать его. если затем этим раздражителем не производится на животное никакого дальнейшего действия. И это прекращение действия основано на развитни торможения в клетке, куда адресовалось раздражение. Следовательно, при трех повторениях агента в третьей вариации без присоединения еды и могло происходить это торможение соответствующей клетки, и, таким образом, для соединения с клеткой пищевого раздражения в ней не было процесса раздражения. Предположение кажется достаточно обоснованным, но оно не может быть признано в данном случае за реализованное. Когда агент в третьей вариации был повторен 240 раз, он был испытан относительно его свойства. На очень коротком расстоянии времени после него (30 секунд) применялся какойнибудь из бывших у собаки условных рефлексов - он отнюдь не оказывался заторможенным. Следовательно, агент не был тормозным и не производил последовательного торможения. Но его тормозное действие должно быть очень концентрированным вследствие долгого употребления. И это не подтвердилось. Когда после 240 раз этот агент начали сопровождать едой сплошь, он уже на третий раз дал значительновный секреторный эффект. Нужно еще прибавить, что в первый день, когда агент был применен в комбинации с пищей два раза, ранние условные рефлексы были малы, т. е. возбудимость собаки была невысока. Таким образом скорость образования нового условного рефлекса была максимальной, и думать о тормозном состоянии клетки индифферентного агента нет основания. Мы испытали предположения, истекающие из известных нам фактов, и причину явления не нашли. Мы будем испытывать ее дальше и, надо думать, найдем. Но сделанное до сих пор показывает, что у нас еще нет сейчас полного знания всех тех условий, которые определяют деятельность механизма образования условного рефлекса. Приведенные во второй лекции условия, как они ни были достаточны для получения всех рефлексов, бывших до сих пор в нашем распоряжении, значит, еще не полны, раз, имея их в голове, мы не можем сразу понять результата анализируемого случая. Есть еще какое-то просматриваемое нами условие. Значит, опять чрезвычайная обусловленность явлений.
Если не может быть речи в настоящее время о понимании всей деятельности больших полушарий из деталей физико-химического процесса, что очевидно; если нет, по всей вероятности, верных шансов для понимания этой деятельности из элементарных свойств нервной ткани; если мы даже относительно самого основного явления этой деятельности не имеем еще полного знания, то что же остается для нашего исследования, к чему сводится наша работа? Очевидно, приведение бесчисленной массы разнообразных отдельных явлений все к меньшему числу общих основных явлений и должно составить ближайшую и реальнейшую задачу теперешней стадии физиологического изучения больших полушарий. Этим мы и занимаемся теперь, то приближаясь несколько к цели в отдельных случаях, то стоя перед неразрешенными, но атакуемыми вопросами, то ошибаясь.
В начале этой работы, опираясь на резкие, но внешние черты, мы различали три вида торможения: внешнее, внутреннее и сонное. Накопляющиися фактический материал позволил нам, как вы уже знаете, слить последние два вида, сведя их внешнюю разницу на второстепенную подробность. Оба они оказались одним и тем же внутренним торможением, раз раздробленным и местно разграниченным, в другой раз слитым и широко распространенным. Когда при дальнейшем изучении условных рефлексов мы познакомились с явлениями взаимной индукции, естественно. бросилось в глаза подобие факта отрицательной индукции с фактом внешнего торможения. Теперь представилась вероятной тождественность в основе процесса при всех наших старых видах торможения, почему мы и приступили к собиранию дальнейших данных в этом направлении. Эти данные частью были сообщены раньше, частью я их приведу теперь. В лекции об индукции был описан опыт, в котором условный оборонительный рефлекс на механическое раздражение кожи на одном определенном месте тормозил в периоде последействия условный пищевой рефлекс на механическое раздражение кожи на других местах. При этом одна подробность свидетельствовала, что это торможение происходило обоими пунктами (пунктами раздражения и торможения), именно в коре полушарий. В лекции о переходных фазах к полному внутреннему торможению было указано на некоторых примерах, что и в периоде внешнего торможения можно наблюдать те же фазы. Сейчас я могу прибавить новые соображения и факты в пользу заключения о тождестве внешнего и внутреннего торможения. В вышеупомянутом факте замаскирования, подавления в комплексном условном раздражителе сильным раздражителем слабого необходимо видеть обнаружение внешнего торможения. Клетка сильного раздражителя тормозит клетку слабого раздражителя, и потому эта последняя клетка может войти только в слабую связь с клеткой безусловного раздражителя, так как есть основание считать чтопень проторенности соединительного пути отвечающей силе условного раздражителя. А чем бы в данном случае от этого внешнего торможения отличалась отрицательная индукция, когда пункт положительного условного раздражителя усиливает или даже восстановляет тормозное состояние пункта условного тормозного раздражителя? Наконец к доказательству устанавливаемого нами положения могут служить и следующие факты. Уже давно многими нашими сотрудниками (Минтовт, Кржышковский, Лепорский) указывалось, что при образовании условного тормоза для быстрого и полного торможения имеет большое значение, чтобы агент, из которого будет вырабатываться условный тормоз, не был значительно физически слабее, чем агент, являющийся положительным условным раздражителем, участником тормозной комбинации. Недавние нарочитые опыты (Фурсикова) вполне подтвердили эти указания, прибавив факт, что часто внешнее торможение, с которого начинается действие нового агента, как вызывающего исследовательский рефлекс, незаметно переходит в тормозное действие комбинации, как внутреннее торможение. И все же остается вопрос: достаточно ли это для отождествления внешнего и внутреннего торможения?
В девятнадцатой лекции был поднят вопрос о трех видах внешних раздражителей, вызывающих прямо тормозное состояние коры больших полушарий: очень слабых, очень сильных и необыкновенных, причем было предположительно указано биологическое значение этого факта. Вопрос о физиологическом механизме действия этих различных раздражителей был отложен до настоящей лекции. Но и здесь сейчас я не нахожу возможным ставить и обсуждать вопрос о торможении во всем его объеме. Как ни значителен наш экспериментальный материал, он явно недостаточен, чтобы составить общее и определенное представление о торможении и об его отношении к раздражению. Объяснения, хорошо подходящие к одной группе фактов, не могут, однако, захватить их все. Многое факты упорно не поддаются теоретическому анализу, и представление об их механизме приходилось несколько раз менять на протяжении нашей работы, не находя его удовлетворительным. И тут, как и вообще в нашей работе, пока остается только группировать фактический материал. Мы стоим перед массой нерешенных вопросов: о растормаживании, положительном действии тормозных условных раздражителей, в некоторых случаях о прямо тормозящем действии положительных условных раздражителей при нарушении коры полушарий, о преимущественном тормозящем действии слабых и сильных положительных раздражителей, и т. д. и т. д. Мы во многих случаях не можем сказать: какие из фактов близки или тождественны, или же совершенно отдельны, радикально различны. Для иллюстрации трудности таких случаев рассмотрю следующий пример. Почему новые явления или новая связь явлений действует на животное тормозящим образом? Какой физиологический механизм этого задерживания? Мы подаем собаке после условного раздражителя корм другим способом, чем раньше, например вместо чашки с пищей, выдвигаемой из-за экрана, сбрасываем ее через трубку из резервуара, стоящего наверху, в чашку, находящуюся перед собакой, и многие из собак тогда ее упорно не берет, и наши условные рефлексы исчезают. Наступает, конечно, торможение. На что его свести, с каким из наших других фактов его поставить рядом? Может быть, с фактом, упомянутым в тринадцатой лекции, когда при резком изменении обычного порядка в применении ряда различных условных раздражителей в течение опытов наступает, особенно у некоторых собак, более или менее значительное торможение всей условнорефлекторной деятельности, держащееся несколько дней, несмотря даже на возвращение к старому порядку? Можно представить себе то же самое и относительно всей окружающей собаку действительности. Постоянно повторяющиеся и в определенном следовании перед животным внешние явления создают в полушариях определенную, так сказать, стереотипную деятельность. Всякое новое явление, или даже и старые, но в новой связи, нарушая этим стереотип, вызывают торможение, как и в наших опытах с изменением порядка условных раздражителей. Но тогда встает следующий вопрос: как, каким ближайшим физиологическим процессом происходит торможение в последнем случае, или, правильнее сказать, с каким в свою очередь другим случаем торможения тождествен этот случай торможения? Не есть ли это результат тормозящего действия исследовательского рефлекса, возникающего при новом колебании внешней среды? Или же это совершенно отдельные, самостоятельные явления? За первое бы говорила резко выраженная у очень тормозимых собак связь чрезвычайно чувствительного ориентировочного рефлекса с сильным последовательным торможением (факт, выступавший исключительно отчетливо y Sмницы), и при таких собаках такой механизм является вероятным. Но в других случаях нет внешних указаний на ориентировочный рефлекс и наступает прямо тормозное состояние.
Этот пример показывает, как мы еще далеки от удовлетворительного группирования наших отдельных фактов.
Вот почему я пока отказываюсь высказываться за ту или другую из существующих теорий торможения или выставлять нобую. Пока мы пользуемся временными предположениями для систематизирования нашего фактического материала и проектырования новых опытов.
В заключение я перехожу к нашим ошибкам, которых мы делали немало в прошлом и которые постепенно поправляли и поправляем. Сейчас сперва опишу одну, которая была открыта и исправлена при повторении старых опытов в течение этих лекции, и другую, вероятную, которую исследуем в настоящее время.
В четвертой лекции, в которой говорилось о восстановлении угашенного условного рефлекса, были приведены три различных случая этого восстановления: самостоятельное по истечении известного срока, прочное, но более или менее медленно наступающее, и ускоренное, раз посредством подкрепления, т. е. применения специального рефлекса, в другой раз вследствие применения каких-нибудь посторонних рефлексов. Эти два последние случая описаны там как существенно различные между собой. При первом из них принималось не только быстрое, но и прочное восстановление. При втором случае с посторонними рефлексами было установлено, что восстановление хотя и быстро наступало, но было временным и скоро исчезало по прекращении постороннего рефлекса и его последействия, сменяясь опять торможением, продолжавшимся до конца его обычного срока, как если бы никакого постороннего рефлекса и не было. Поэтому это восстановление получило особенное название «растормаживания». При этом там было указано на трудность понимать эту разницу, раз угасание не было окончательным разрушением условного рефлекса. В последнее время при получении новых данных относительно взаимодействия условного и безусловного раздражителей мы сочли надобным переделать возможно точно старые опыты и убедились в ошибочности их (новый опыт Подкопаева). Восстановление угашенного рефлекса при применении как специального, так и всякого постороннего рефлекса только временное, т. е. после обоих рефлексов в ближайшее время угашенный условный раздражитель действует положительно, а несколько позже опять лишается этого действия до его самостоятельного восстановления. Когда условный рефлекс - пищевой, а посторонний - кислотный, т. е. оба химические, то временное восстановление, растормаживание происходит совершенно однообразно, что касается его течения во времени. Опыты были поставлены на двух собаках и дали один и тот же результат. Даю подробное описание опыта на одной из собак. При угасании условного пищевого рефлекса на удары метронома до нуля этот нуль держался до 20 минут после последнего повторения условного раздражителя без подкрепления. Затем начиналось постепенное самостоятельное восстановление, которое к 30 минутам достигало 40 0/ % первоначальной величины условного эффекта до угасания. Когда при первом нуле угасания условный раздражитель сейчас же подкреплялся и через 20 минут от нуля он испытывался опять, то оставался еще нулевым. Когда же в другом опыте после такого же подкрепления он применялся спустя только 10 10 минут после угасания, от него имелся положительный эффект. Совершенно так же шел опыт, когда вместо пищи в тот же момент вливалась в рот собаки кислота и в те же сроки испытывался условный раздражитель.
Вот подлинные цифры, взятые из некоторых опытов для примера.
Условный раздражитель при первом применении в опытный день дал 6 капель за 20 секунд; он же после конца угасания, тотчас же подкрепленный и испытанный 10 минут спустя, считая от конца угасания, дал 3 капли.
В другой день условный раздражитель при первом применении дал 7 капель; он же после конца угасания, тотчас же подкрепленный и испытанный 20 минут спустя, считая от конца угасания, остался без малейшего эффекта.
Ус Условный раздражитель при первом применении в опытный день дал 5 капель за 20 секунд; он же после конца угасания тотчас же сопровождался вливанием кислоты в рот собаки и испытанный 10 минут спустя, считая от конца угасания, дал 2 капли.
В другой день условный раздражитель при первом применении дал 5 капель; он же после конца угасания, тотчас же сопровождаемый вливанием кислоты и испытанный 20 минут спустя, считая от конца угасания, остался без малейшего эффекта.
Максимум растормаживания наступает в более ранние сроки после конца угасания, чем 10 минут.
Очевидно, прежняя ошибка была основана частью на неточности опытов, когда сравнение сроков восстановления при специальном рефлексе и посторонних рефлексах чаще всего происходило при применении таких посторонних рефлексов, как звуковые, зрительные и другие, имевших короткое последействие, а не химических с их длинным последействием. Частью же к старому заключению располагала мысль, что безусловный раздражитель, с которым сочетался условный, должен находиться в особенном отношении к последнему в случае его восстановления после угасания. Теперешний же результат представляет лишнее основание для принятия, что торможение происходит именно в нервной клетке, а не в соединительном пункте или пути между клеткой условного раздражителя и клеткой специального безусловного раздражителя. Иначе было бы трудно себе представить, как восстановляющее действие обоих этих раздражителей могло оказаться совершенно одинаковым.
Другая ошибка обнаруживается сейчас, и я нахожу возможным говорить о ней сейчас, хотя переисследование предмета только начинается, с одной стороны, ввиду особенной важности пункта, к которому она относится, с другой - опять же ради общей характеристики нашей области, изобилующей исключительными трудностями при установлении точных фактических отношений.
Во второй лекции приведены были, казалось, все условия, при которых образуются условные рефлексы, и все, что мешает их образованию. На втором месте после первого условия совпадения во времени индифферентного агента и безусловного раздражителя - было поставлено непременное, хотя бы и очень короткое, предшествование по времени будущего условного раздражителя перед безусловным. На многих вобаках после многочисленных повторений (даже 300-400 раз) комбинации индифферентного агента с безусловным раздражителем, причем комбинация начиналась с безусловного раздражителя и к нему индифферентный агент присоединялся только спустя 5-10 секунд, условный рефлекс не вырабатывался, хотя у тех же собак при других разных агентах, сочетаемых обычным образом с безусловным раздражителем, условные рефлекса были готовы через 7-20 повторений. Естественно было думать, что сильный безусловный раздражитель, сосредоточенный в определенном месте полушарий механизмом внешнего торможения, обусловливал такое тормозное состояние в остальных частях полушарий, что раздражения, падающие на эти части, оказывались недействительными, совершенно так, как это мы знаем по себе, когда, чем-нибудь сильно занятые, мы не слышим и не видим, что происходит около нас. Таким образом факт казался очевидным и понятным как с нашей точки зрения, так и с общежитейской. Это и сделало то, что до самого последнего времени у нас не было относительно него ни малейшего сомнения. Теперь положение дела изменилось. Когда мы недавно задались вопросом: каким образом раннее подкрепление условного раздражения, т. е. возможное укорочение его изолированного действия, мешает развитию тормозного состояния в пункте его приложения в коре, то новым варьированием опытов были приведены к неожиданному для нас факту. Оказалось, что безусловный раздражитель тормозит и выработанный условный раздражитель. Но тогда естественно и обратило на себя внимание следующее сопоставление фактов.
Имеется ли уже выработанный условный раздражитель, или это еще индифферентный агент, из которого будет только вырабал батываться этот условный раздражитель, - влияние на него безусловного раздражителя одно и то же. А вместе с тем влияние это выражается в том, что при малейшем изменении внешних условий, условий времени, происходит образование условного рефлекса, превращение индифферентного агента в определенный условный раздражитель. Приходилось признать, что механизм образования условного рефлекса и механизм внешнего торможения как-то сближаются или даже отождествляются, иначе говоря, что процесс внешнего торможения дает основание для образования связи между клетками. Это напоминало бы другой из наших фактов, именно образование вторичного условного рефлекса и развитие условного торможения, где также при общих внешних условиях, но при небольшой разнице в отношении времени, наступает то процесс раздражения, то процесс торможения. Если бы это соображение было верно, то надо было бы ждать, что и при покрытии, как мы выражаемся, с самого начала безусловным раздражителем индифферентного агента между пунктами их в коре вначале могла бы образоваться связь, мог бы произойти условный рефлекс, но который потом при повторении такой комбинации затормаживался. И это показали наши первые ориентировочные опыты. Опираясь на то, что полное покрытие выработанных условных раздражителей безусловным только постепенно ведет к уменьшению их эффекта, при слабых к более скорому и даже к нулю, а при сильных к более медленному и часто к незначительному, мы при новых комбинациях безусловного раздражителя с индифферентными агентами, - причем комбинация начиналась с безусловного раздражителя, - применяли эту комбинацию только малое число раз, чтобы не дать развиться торможению. Во многих случаях мы действительно получили то, что ожидали. После таких комбинаций индифферентный агент, испытанный отдельно, действовал как условный раздражитель (в опытах В. И. Павловой, Крепса, Подкопаева и Купалова). Когда мы с этим взглядом на дело и с первыми опытами в этом направлении обратились к нашим старым опытам (опыты Крестовникова), мы увидали, что, во-первых, почти всегда при них индифферентные агенты испытывались на их условное действие только после большого числа повторений комбинаций, в которых они присоединялись к безусловному раздражителю после начала его действия, а, вовторых, даже и при этом часто сильные индифферентные агенты при отдельной их пробе долгобнаруживали секреторный эффект. Но этот эффект истолковывался как случайный и посторонние, а не как условный, в силу предвзятой мысли, что, если бы он был действительно условным, он должен был бы c повторением комбинации увеличиваться, не уменьшаться и исчезать, как это было в тех опытах. Возможно, что и другое явление, именно отсутствие при пробах ясной условной двигательной реакции, при этой предвзятости тоже не вполне объективно оценивалось. На одной собаке должно было быть признано образование подлинного условного рефлекса, но оно также было сведено на постороннее условие, конечно с известным основанием, - но вполне ли достаточным? Тема будет обработана многосторонне и строго при теперешнем очень увеличенном запасе наших сведений о предмете и при большей специальной тренировке нашей мысли в новой области исследования. При полном оправдании наших соображений и при дальнейшем подтверждении наших теперешних ориентировочных опытов физиология больших полушарий овладела бы очень важным для приложения к человеку положением, что образование новых связей в коре может происходить не только в районах полушарий с оптимальной возбудимостью, но и в частях их, более или менее заторможенных.
в этой последней, также фактической, лекции я, однако, хотел, как только мог, передать основное впечатление от работы в нашей области. Как мне кажется, многочисленные факты, сообщенные во всех предшествующих лекциях, достаточно свидетельствуют, что в этой области возможна серьезная естественнонаучная работа, накопление точных фактических данных. Поэтому я не боялся дискредитировать наше научное предприятие тем, что привел в этой лекции. Как всегда, лучше, конечно, сознавать опасности, чем их игнорировать. Кроме того, мне хотелось предупредить будущих работников этой области о тех исключительных трудностях, которые они встретят в ней.
В общем же этот новый отдел физиологии поистине пленителен, удовлетворяя двум, всегда рядом идущим, тенденциям человеческого ума: стремлению к захватыванию все новых и новых истин и протесту против претензий как бы законченного где-нибудь знания. Здесь гора неизвестного явно надолго останется безмерно больше кусочков отторгнутого, познанного.
Лекция двадцать третья. Приложение экспериментальных данных, полученных на животных, к человеку
Мм. гг.! Если сведения, полученные на высших животных относительно функций сердца, желудка и других органов, так сходных с человеческими, можно применять к человеку только с осторожностью, постоянно проверяя фактичность сходства в деятельности этих органов у человека и животных, то какую же величайшую сдержанность надо проявить при переносе только что впервые получаемых точных естественно-научных сведений о высшей нервной деятельности животных на высшую деятельность человека. Ведь именно эта деятельность так поражающе резко выделяет человека из ряда животных, так неизмеримо высоко ставит человека над всем животным миром. Было бы большим легкомыслием первые шаги физиологии больших полушарий, полной по программе только, а, конечно, не по содержанию, считать уже за какое-то решение грандиозной задачи о высшем механизме человеческой натуры. Поэтому всякая узкая регламентация в настоящее время работы об этом предмете была бы только свидетельством чрезвычайной ограниченности мысли. Но, с другой стороны, временно, конечно, чрезвычайно упрощенное тестирование предмета со стороны естествознания не должно встречаться враждебно, что, к сожалению, также случается нередко. Сложное берется наукой только по частям и обрывками, но оно захватывается ею постепенно все более и более. Следовательно, будем надеяться и терпеливо ждать, когда точное и полное знание нашего высшего органа - головного мозга - сделается нашим подлинным достоянием, а с этим и главной основой прочного человеческого счастья. После того, что приведено во всех предшествующих лекциях, едва ли можно оспаривать, что самые общие основы высшей нервной деятельности, приуроченной к большим полушариям, одни и те же как у высших животных, так и у людей, а потому и элементарные явления этой деятельности должны быть одинаковыми у тех и у других как в норме, так и в патологических случаях. На нормальных случаях, как очевидных, я остановлюсь коротко, приведя лишь немногие отдельные примеры, главным же образом заиму теперь ваше внимание патологическими случаями. Очевидно, наше воспитание, обучение, дисциплинирование всякого рода, всевозможные привычки представляют собой длинные ряды условных рефлексов. Кто не знает, как установленные, приобретенные связи известных условий, т. е. определенных раздражений, с нашими действиями упорно воспроизводятся сами собой, часто даже несмотря на нарочитое противодействие с нашей стороны? Это одинаково касается как производства тех или других действий, так и выработанного их задерживания, т. e. как положительных, так и отрицательных рефлексов. Известно далее, как иногда нелегко развить нужное торможение в случае как отдельных лишних движений при играх, при манипуляциях в разных искусствах, так и действий. Точно так же практика давно научила, как исполнение трудных задач достигается только постепенными и осторожными подходами к ним. Все знают, как экстренные раздражения задерживают и расстраивают хорошо налаженную обычную деятельность и как путает и затрудняет изменение раз установленного порядка движений, действий и целого уклада жизни. Опять общеизвестно, что слабые и однообразные раздражения делают людей вялыми, сонливыми, а некоторых и прямо усыпляют. Так же хорошо знакомы всем разные случаи частичного бодрствования при обыкновенном сне, например случай спящей матери у больного ребенка, и т. д. и т. д. Это все факты, с которыми мы раньше в этих лекциях встречались на наших животных. Обращаюсь к патологическим случаям. Современная медицина различает нервные и душевные болезни, неврозы и психозы. Но различение это, конечно, совершенно условное. Точной разграничительной линии между теми и другими никто бы провести не мог, потому что ее нет и в действительности. Как можно было бы себе представить психическое расстройство без нарушения мозговой ткани, если не в структурном, то в функциональном отношении? Различие между нервным и психическим заболеванием есть различие или по сложности, или по тонкости нарушения нервной деятельности. К этому нас склоняют и опыты на наших животных. Пока мы имеем дело с животными, у которых наши разные приемы функционального характера или чрезвычайности жизненных условий (прошу припомнить случай с наводнением), или, наконец, незначительные операции на полушариях нарушают их нервную деятельность, мы можем понимать более или менее удовлетворительно механизм этих нарушений в терминах нервной физиологии. Но как только мы разрушили большие участки полушарий или делает это разрастающаяся рубцовая ткань, для нас возникает трудность всегда полно и ясно представить себе механизм наступающих расстройств в нервной деятельности, и мы прибегаем к предположениям, которыеще требуют доказательства их соответствия действительности. Очевидно, разница в нашем положении относительно предмета в том и другом случае имеет сбое основание в большей сложности нарушений в последнем случае и в недостаточности для них теперешнего физиологического анализа. Наверное, многие врачи психологи, смотря на тех и других животных, про первых сказали бы, что они нервнобольные, последних назвали бы душевнобольными. Мы же, отказываясь \входить воображаемый внутренний мир наших собак, повторили бы при этом, что перед нами нарушенная деятельность больших полушарий --- в первом случае меньше и проще, во втором случае больше и сложнее. Теперь сопоставим разные нервные расстройства у наших животных и у людей. На собаках мы познакомились с двумя условиями, производящими функционально нервные расстройства. Это - трудная встреча, столкновение раздражительного и тормозного процессов и сильные чрезвычайные раздражения. Они же составляют и обыкновенные причины нервных и психических заболеваний людей. Жизненные положения, возбуждающие нас в высшей степени, например в случае жестоких оскорблений, сильного горя, и вместе с тем обязывающие нас сдерживать, подавлять естественные реакции на них, часто и ведут к глубокому и долговременному нарушению нервного душевного равновесия. С другой стороны, люди также часто делаются нервно и психически больными, подвергаясь чрезвычайным опасностям, угрожающим им самим, а также их дорогим близким, или даже только присутствуя при страшных событиях, ни их самих, ни их близких непосредственно не касавшихся. При этом замечается, как правило, что те же обстоятельства остаются без этих последствий для других лиц, как говорят, не расположенных к заболеванию, т. е. с более сильной нервной системой. Совершенно то же самое наблюдалось и на наших собаках. В отношении таких заболеваний имеются очень большие различия между отдельными животными. У нас были собаки, у которых один из действительнейших приемов для нарушенервного равновесия, именно непосредственная смена тормозного ритма механических раздражений кожи на одном и том же месте на положительный, при ежедневном повторении в течение долгого срока времени, нисколько не влиял вредным образом на животное. У других нервное расстройство появлялось только после многократного повторения. У третьих же нервное состояние наступало уже после однократного применения приема. Точно так же, как это было упомянуто ранее, чрезвычайное наводнение произвело заболевание, очевидно аналогичное травматическому шоку людей, только у немногих собак, именно очень тормозимых. Далее только что упомянутый прием, как сообщалось раньше, обусловливает разные формы заболевания в связи с разными типами нервной системы: то в сторону преобладания раздражительного процесса у собак с более сильной нервной системой, то в сторону преобладания тормозного процесса у собак с более слабой нервной системой. Насколько позволительно судить, главным образом на основании обыденных наблюдений, мне кажется, что этим двум вариациям нарушения нервной деятельности у животных отвечают две невротических человеческих формы: именно неврастения и истерия, если первую характеризовать преобладанием процесса раздражения и слабостью тормозного, а вторую, наоборот, преобладанием тормозного и слабостью процесса раздражения. Есть практические основания признавать неврастеников, по крайней мере некоторых из них, за сильных, способных даже к очень большой работе, людей, истерики же, конечно, совершенно не жизненные субъекты, полные инвалиды. Что неврастеники вместе с тем имеют периоды бессилия, временной негодности, - это понятие, раз в другое время они так продолжительно возбудимы и продуктивны: нервная расточительность должна же быть возмещена. Можно бы сказать, что они представляют собой другую, именно более длинную против обычной, периодичность в смесе работы и отдыха, и поэтому у них сравнительно с обыкновенными уравновешенными людьми так утрированы периоды возбуждения и торможения. Что у истериков, с другой стороны, бывают припадки возбуждения, конечно, нисколько не говорит за силу их нервной системы. Это возбуждение бесцельно, бесплодно, так сказать грубо механично. В нашем наблюдательном материале на собаках, как мне кажется, имеются некоторые указания на происхождение и характер этого возбуждения. У нас была собака (описанная Фроловым) очень тормозимого типа, говоря обыденным языком, - в высшей степени трусливое, покорное существо. Эта собака служила для опытов над желудочной секрецией и должна была выстаивать в станке многие иасы. При этом замечалось, что она при этом никогда не спала, но стояла в бодрой позе удивительно покойно, почти не шевелясь, лишь изредка осторожно переступая ногами. Однако это не было оцепенение. На обращение к ней с кличкой она реагировала. Но в момент снятия ее со станка, когда начинали ее освобождать от привязи, она приходила в совершенно невероятное возбуждение: визжала и сильнейшим образом рвалась, так что могла опрокинуть и станок. И это возбуждение ничем нельзя было прекратить, ни громкими окриками, ни ударами, собака становилась совершенно неузнаваемой. Несколько минут прогулки на дворе - и собака делалась прежней: сама шла в экспериментальную комнату, вскакивала на станок и опять стояла неподвижной. Мочеиспускательный и испражнительный рефлексы всываемом факте существенной роли не играли. Подобный факт наблюдался иногда и у других собак, но никогда не выражался в такой исключительно резкой форме. Всего проще понять его как кратковременную положительную индукцию, взрыв возбуждения после продолжительного и напряженного торможения. Это могло бы быть одной из причин припадков возбуждения у истеричных при их частых и глубоких проявлениях торможения. Но вероятно соучастие и другой причины, демонстрированной другой нашей собакой (описанной Подкопаевым). Это - спокойное, уравновешенное животное, малоподвижное, на станок само никогда не вскакивает, в станке держится неподвижным, но отнюдь не спит, положительные и отрицательные условные рефлексы очень постоянны и точны. У нее с голени через туловище до запястья на одной половине тела расположен ряд приборчиков для механического раздражения кожи. Из раздражения голени сделан условный положительный пищевой раздражитель, из раздражения всех остальных мест - отрицательные раздражители. Последние вырабатывались скоро и были постоянными. Собака при всех кожных раздражениях оставалась спокойной, не делая совершенно местных движений, даже двигательная пищевая реакция при условном раздражении почти отсутствовала; еду брала не спеша. Выработка отрицательных рефлексов была начата с раздражения запястья как самого удаленного пункта от места положительного рефлекса. Такое положение оставалось некоторое время. Затем неожиданно раздражение на запястье начало сопровождаться двигательной реакцией в виде резкого отдергивания раздражаемой конечности. Иногда эти отдергивания совпадали с ритмом механических раздражений. Затем такие же местные реакции стали появляться последовательно на тормозных местах, все более и более приближающихся к месту положительного раздражения, причем двигательная реакция делалась все более обширной в виде переступания всех конечностей. Голова и шея оставались неподвижными, так сказать, безучастными к тому, что происходило в задней части тела. Слюноотделения при этом не было. Когда раздражение на бедре, ближайшем пункте к положительному, было сделана также положительным, описанная двигательная реакция на него совершенно исчезла. То же произошло и с другими местами, когда раздражения их также были превращены вожительные, кроме самых удаленных двух, которые при раздражении вместе с полным слюноотделительным эффектом все же обнаруживали местную оборонительную реакцию, хотя в ослабленной форме. Ход развития описываемого явления (не с начала опытов с каждым раздражителем, а лишь после выработки дифференцировки) и местный характер его дают основания для заключения, что это - спинномозговые рефлексы, появлявшиеся благодаря функциональному и притом частичному выключению коркового кожного анализатора. То же можно было бы принимать в некоторых случаях и у истеричных субъектов при их корковых торможениях. В нашем материале есть и другие случаи, которые тоже отвечают более или менее известным патологическим состояниям нервной системы у людей. Прошу припомнить собаку (Рикмана), которая была приведена в такое состояние, что совершенно не выносила условных раздражителей из физически сильных агентов, сейчас же непременно переходя в тормозное состояние, и лишь при слабых раздражениях могла быть и продолжаться условнорефлекторная деятельность. Едва ли будет натяжкой, если я этот случай, конечно не в полном размере, а лишь по его механизму, буду аналогировать со случаями продолжительного многогодового сна у людей, как это было описано Пьером Жанэ на одной молодой девушке и как это наблюдалось в одной из петербургских психиатрических больниц на взрослом мужчине. Дело шло о пациентах, которые, казалось, были погружены в постоянный сон. Они не делали никаких движений, ничего не говорили, приходилось искусственно их кормить и держать опрятными. Лишь ночью, когда затихала дневная жизнь с ее разнообразными и сильными раздражениями, для них являлась иногда возможность некоторой деятельности. Пациентка Пьера Жанэ ночью порой ела и даже писала. О петербургском больном тоже были сообщения, что он изредка ночью вставал. Когда этот больной почти после двадцатилетнего сна к старости (60 годам) начал освобождаться от него и мог говорить, он заявил, что часто и прежде и слышал и видел, что происходило около него, но не имел силы сделать какое-либо движение или что сказать. В этих обоих случаях явно имелась в сильной степени ослабленная нервная система и ослабленные специально большие полушария, которые сильными внешними раздражениями быстро переводились в сплошное тормозное состояние, в сон. На той же собаке мы выше познакомились и с другим патологическим симптомом нервной деятельности, который также, по нашему мнению, нередко воспроизводится невропатологической казуистикой людей. У нее было хроническое узко локализированное функциональное поражение слухового анализатора в коре, прикосновение к которому соответственным раздражением производило последовательно тормозное состояние всей массы полушарий. Сколько есть разнообразных случаев болезненного нервного состояния людей, когда у них нормальная деятельность поддерживается более или менее только до тех пор, пока их не коснутся компоненты, хотя бы и очень незначительные, даже в виде словесных намеков, тех сильных и сложных раздражителей, которые первоначально обусловили нервное заболевание. Наконец здесь же надо напомнить приведенный в девятнадатой лекции случай периодического зрительноео иллюзионирования одной из наших собак. Оно имело, по всей вероятности, свое основание в осложнении внешнего раздражения коры виутренним, исходящим из действия разрастающегося рубца. Так же могут пониматься и некоторые случаи иллюзий у людей при тех или других внутренних раздражениях коры. Это из области патологии. То же подобие между нашими экспериментальными животными и нами наблюдается и в отношении лечения нервных расстройств помимо тождественного действия фармацевтических средств. Как уже и раньше сообщалось, отдых, вообще перерыв опытов, часто помогал возврату к норме. При этом наблюдаются некоторые подробности, одну из которых считаю нелишним привести здесь. Одна из наших собак приемом столкновения тормозного и раздражительного процесса была приведена в чрезвычайно возбужденное состояние (опыты Петровой). Все виды внутренего торможения были нарушены, т. е. все ее отрицательные условные рефлексы превратились в положительные. При всех условных раздражителях, как прежних положительных, так и при прежних отрицательных, начиналась одышка как обыкновенный симптом сильного возбуждения. Отмена отрицательных условных рефлексов положение дела не изменила. Продолжалась одышка, и положительные рефлексы оставались очень увеличенными сравнительно с нормой. Тогда было решено из условных положительных раздражителей применять только физически слабые, т. е. световое и кожно-механические, отменив звуковые как обыкновенно в наших опытах физически более сильные. Благоприятный результат обозначился сейчас же. Животное стало спокойным. Одышка исчезла. Величина слюноотделительного эффекта вернулась к норме. Некоторое время спустя можно было постепенно ввести и сильные положительные раздражители без того, чтобы нарушить результат лечения. Больше того. Еще позднее на несколько дней и дифференцировка кожного раздражения по месту как более легкий вид внутреннего торможения оказалась налицо, не вызвав возбуждения животного. К сожалению, опыт на этом был прерван за неимением времени у экспериментатора. Интересный случай, показывающий, как уменьшение количества внешней энергии, поступающей в большие полушария в виде условных раздражений, повело к падению болезненно повышенного положительного тонусаполушарий. Конечно, людская нервная терапия широко пользуется в виде разнообразных жизненных рецептов ограничением внешних раздражений, падающих на болезненно раздраженные полушария. Позволяю себе привести здесь подробно еще один наш случай, представляющийся мне очень поучительным с точки зрения терапии. Этот случай относится к собаке с совершенно необычной, очевидно ненормальной, реакцией на механическое кожное раздражение, реакцией, носящей характер какого-то сильного возбуждения больших полушарий (наблюдения и опыты Пророкова). При обыкновенном нашем механическом раздражении кожи на бедре собака сейчас же начинает вертеть задом, переступает всеми конечностями, закидывает как-то странно голову кверху, при этом подвизгивает и иногда зевает. При подаче кормушки во время еды реакция прекращается. Сверх ожидания своеобразная реакция нисколько не помешала образованию условного рефлекса на раздражение кожи, что обыкновенно делают местные двигательные рефлексы (отдергивание соответственной конечности, местная игра под местом раздражения platysma myoides), встречающиеся у других собак. Здесь, наоборот, рефлекс образовался скоро, и, что совершенно исключительно, этот кожно-механический условный рефлекс по его слюноотделительному эффекту был в громадном большинстве случаев больше, чем при самых сильных звуковых раздражениях. Точно так же и пищевая двигательная реакция, обыкновенно сменявшая своеобразную вышеописанную реакцию в половине срока каждого изолированного условного кожно-механического раздражения, была резко усилена сравнительно с тем, какой она была при других раздражителях. Обыкновенно продолжающееся некоторое время по прекращении подкармливания пищевое возбуждение животного тоже при этом было и продолжительнее и сильнее. Кроме того, при применении в опытах кожного раздражения собака сделалась вообще очень возбудимой. На малейший звук, исходивший из-за двери, за которой находился экспериментатор, собака реагировала комплексом тех же особенных движений. Все это вело к заключению, что кожное раздражение у этой собаки вызывало сильное разлитое возбуждение больших полушарий. Что это за возбуждение - осталось для нас невыясненным. Против полового характера говорило отсутствие эрекции. Мы остановились на предположении, не аналогично ли это эффекту щекотания? Во всяком случае это было необычное, ненормальное нервное явление, и мы поставили себе задачей устранить его. Для этого мы применили процедуру развития внутреннего торможения в виде дифференцировки кожного раздражения по месту. Раздражение на плече сначала дало и особенную реакцию и условную в силу первоначальной генерализации условного раздражения. Но при повторении его без сопровождения подкармливанием скоро исчезла основная пищевая реакция, как двигательная, так и секреторная (к восьмому разу), a затем (к сороковому разу) и особенная двигательная. На бедре же раздражение давало прежнюю картину и особенной и пищевой двигательной реакций, сменявших одна другую. Затем было прибавлено кожное раздражение ближе к бедру, на боку, тоже дифференцируемое. Повторилось совершенно то же самое, что было при раздражении на плече, а особенная реакция на бедре все оставалась, не уменьшаясь. Наконец была выработана дифференцировка на задней лапе. На этот раз особенная двигательная реакция при раздражении бедра начинает ослабевать и, наконец, исчезает совершенно. Таким образом развитие обширного торможения в корковом конце кожного анализатора устранило своеобразный посторонний кожный рефлекс, сохранив и сделав нормальным (из увеличенного) условный кожный пищевой рефлекс. Этот случай, как и некоторые другие наблюдения, подал нам мысль применять прием постепенного развития торможения в полушариях для восстановления вообще нарушенного в них равновесия. Мы пробуем это на собаке, у которой имелся описанный в восемнадцатой лекции узко локализированный болезненный пункт в звуковом анализаторе. Так как этот пункт был специально связан с ударами метронома, то мы при других звуковых раздражениях, действующих на здоровые пункты анализатора, развиваем дифференцировочное торможение в расчете, что иррадиирование его на больной метрономный пункт, может быть, подействует на него благоприятно, вернув его к нормальной возбудимости деятельности. Опыт ведется. Я не знаю, может быть, что-нибудь подобное применяется и в людской нервной терапии, если не считать разные успокаивающие процедуры виде тепловатых вани и т. д. Теперь я займу ваше внимание такими состояниями нервной деятельности наших собак, частью нормального, частью патологического характера, которые, если бы их переносить на человека, пришлось бы называть психическими. Это у собак - гипнотические фазы, переходные между бодрым состоянием и сном, и пассивно-оборонительный рефлекс. Мы видели в шестнадцатой лекции, что переход животного из бодрого состояния в сон имеет свое основание в развитии в головном мозгу тормозного процесса, начинающегося в полушариях под влиянием определенных раздражений и представляющего разные степени экстенсивности и интенсивности в различных фазах наступающего сонного состояния. В настоящее время едва ли можно сомневаться в том, что этих фактов на животном более или менее достаточно для физиологического понимания основных явлений человеческого гипнотизма. Прежде всего идет вопрос об условиях, вызывающих гипнотические состояния. У животных, как мы это уже знаем, они наступают более или менее медленно при слабых и средних однообразно и продолжительно повторяющихся раздражениях (обыкновенный случай при наших опытах), быстро - при сильных раздражениях (случай давнего гипноза животных). При этом непосредственно действующие раздражители, как слабые, так и сильные, могут быть сигналызируемы другими раздражениями, являющимися условными по отношению к первым. Прошу припомнить особенный способ образования условных отрицательных рефлексов, описанных в конце шестой лекции (ольборта), когда индифферентные раздражители, повторенные несколько раз одновременно с ранее выработанными тормозными, тоже делались тормозными. Процедура гипнотизирования людей вполне воспроизводит описанные условия у животных. Ранний классический способ гипнотизирования -- это так называемые пассы, т. е. слабые, однообразно повторяющиеся раздражения кожи, как в наших опытах. Теперь постоянно применяющийся способ - повторяющиеся слова (к тому же произносимые в минорном однообразном тоне), описывающие физиологические акты сонного состояния. Эти слова суть, конечно, условные раздражители, у всех нас прочно связанные с сонным состоянием и потому его вызывающие. На этом основании может гипнотизировать и гипнотизирует все, что в прошлом совпадало несколько раз с сонным состоянием. Это все аналоги цепных отрицательных рефлексов (Фольборта), подобных цепным условным положительным рефлексам, т. е. рефлексам разных порядков, описанным в третьей лекции. Наконец гипнотизирование истеричных, по Шарко, достигается сильными неожиданными раздражителями, как в старом способе гипнотизирования животных. Конечно, при этом могут действовать также и физически слабые раздражители, сигнализирующие сильные, т. е. сделавшиеся в силу совпадения во времени условными по отношению к сильным. Как у животных, так и у людей большинство гипнотизирующих приемов тем скорее и вернее приводят к цели, чем они чаще применяются. Одно из первых проявлений гипноза - потеря человеком произвольных движений и каталепсия, т. е.ерживание частями тела положения, приданного им внешней силой. Конечно, это есть результат изолированного торможения двигательного анализатора (двигательной области коры), не спустившегося на двигательные центры, лежащие ниже полушарий. При этом другие отделы полушарий могут правильно функционировать. Загипнотизированный может понимать, что мы ему говорим, может знать от нас, какую мы ему придали уродливую форму, желать ее изменить и все же не иметь возможности это сделать. Все это наблюдается при гипнотическом состоянии и у животных. Мы уже упоминали в лекции о гипнотических состояниях, как некоторые собаки совершенно сохраняют активную позу, но теряют все условные рефлексы полностью. Случай заторможенности всей массы полушарий без перехода торможения ниже полушарий. Другие собаки реагируют на все условные раздражители деятельностью слюнных желез, а еды не берут. Случай торможения только двигательного анализатора. Наконец у животных, загипнотизированных по давнему способу, оставались неподвижными туловище и конечности, а глаза часто следили за тем, что происходило около них, и они даже иногда могли есть предлагаемую им пищу. Случай еще более дробного торможения, так что, кроме всей остальной массы полушарий, даже двигательный анализатор не был заторможен полностью. Конечно, совершенно понятны при полном торможении этого анализатора как животных, так и у человека тонические местные рефлексы при соответственных внешних раздражениях. Когда дело касается более сложных форм гипнотического состояния, понятно, что провести полную параллель между животным и человеком становится трудным или даже сейчас и невозможным по нескольким причинам. Может быть, мы еще не уловили всех фаз гипнотического состояния, специально что касается степени его интенсивности, а наверное точно не знаем их порядка и преемственности, как этозано раньше. Мы, вероятно, незнакомы со всеми формами его проявления у животных, не наблюдая животных среди их обычной жизни индивидуальной и социальной, а только в узкой обстановке лабораторного опыта, т. е. как бы отвлеченно ох всего поведения. Значит, мы или не умеем еще ставить всех нужных вариаций опытов, а, может быть, иногда не догадываемся правильно отмечать и понимать все сюда относящиеся явления. А у человека мы знакомы с ними при более разнообразных условиях жизни, воспроизводим их и исследуем их, пользуясь грандиозной сигналыстикой речи. Конечно, надо иметь также в виду и то, что при чрезвычайной разнице в сложности поведения человека и животных у последних, может быть, и вовсе нет некоторых форм проявлений гипнотического состояния. Поэтому дальше приходится пользоваться элементарными данными, полученными на животных, только для некоторого пробного физиологического понимания разных обнаружений гипнотического состояния у человека. Возьмем автоматизм загипнотизированных, когда они стереотипно воспроизводят то, что проделывает перед ними гипнотизер, или когда правильно исполняют движения (ходят) по сложному, запутанному и трудному пути. Очевидно, перед нами известная заторможенность некоторых отделов полушарий, которая исключает нормальную более или менее сложную деятельность, направляемую новыми или хотя бы и старыми, но вновь постоянно комбинирующимися раздражениями данного момента. Эта заторможенность, однако, допускает или даже улучшает, вне влияния сложных воздействий, старую, прочно и давно практикованную связь известных раздражений с определенной деятельностью, с определенными движениями. Таким образов в яркой форме при гипнозе воспроизводится подражательный рефлекс, при помощи которого у всех нас в детстве складывается и вырабатывается сложное индивидуальное и социальное поведение. Точно так же смена предметов с их особенностями, вызывавшая раньше и многократно соответственные движения и действия, последовательно раздражая известные анализаторы находящегося в определенной стадии гипноза человека, безошибочно стереотипно направляет его среди этой смены. Разве это не обычная вещь, что мы, занятые главным образом одним делом, одной мыслью, можем одновременно исполнять другое дело, очень привычное для нас, т. е. работать теми частями полушарий, которые находятся в известной степени торможения по механизму внешнего торможения, так как пункт полушария, связанный с нашим главным делом, конечно, является тогда сильно возбужденным? В, что это понимание дела отвечает действительности, я убеждаюсь теперь постоянно на себе при стариковском падении реактивности мозга (ухудшающаяся память событий текущего времени). Чем дальше, тем больше я лишаюсь способности, занятый одним делом, вести исправно другое. Очевидно, сосредоточенное раздражение определенного пункта при общем уменьшении возбудимости полушарий индуцирует такое торможение остальных частей полушарий, что условные раздражители старых, прочно зафиксированных рефлексов оказываются теперь ниже порога возбудимости. Описанное состояние загипнотизированных субъектов, может быть, можно было бы аналогировать с той гипнотической стадией собак, которую мы называем наркотической, когда сильные и старые рефлексы продолжают существовать, а слабые и молодве исчезают. Среди гипнотических явлений у человека привлекает к себе и законно - особенное внимание так называемое внушение. Как понимать его физиологически? Конечно, слово для человека есть такой же реальный условный раздражитель, как и все остальные общие у него с животными, но вместе с тем и такой многообъемлющий, как никакие другие, не идущий в этом отношении ни в какое количественное и качественное сравнение с условными раздражителями животных. Слово, благодаря всей предшествующей жизни взрослого человека, связано со всеми внешними и внутренними раздражениями, приходящими в большие полушария, все их сигнализирует, все их заменяет и потому может вызвать все те действия, реакции организма, которые обусловливают те раздражения. Таким образом внушение есть наиболее упрощенный типичнейший условный рефлекс человека. Слово того, кто начинает гипнотизировать данного субъекта, при известной степени развивающегося в коре полушарий торможения, концентрируя по общему закону раздражение в определенном узком районе, вызывает вместе с тем естественно глубокое внешнее торможение (как в только что указанном моем собственном случае) во всей остальной массе полушарий и тем самым исключает какое-либо конкурирующее воздействие всех других наличных и старых следов раздражений. Отсюда большая, почти неодолимая сила внушения как раздражителя во время гипноза и даже после него. Слово и потом, после гипноза, удерживает свое действие, оставаясь независимым от других раздражителей, неприкосновенное для них, как в момент его первоначального приложения к коре не бывшее с нами в связи. Многообъемлемость слова делает понятным то, что внушением можно вызывать в гипнотизируемом человеке так много разнообразных действий, направленных как на внешний, так и внутренний мир человека. Можно бы возразить, откуда идет такая сила внушения сравнительно с сновидениями, которые большей частью позабываются и только редко имеют некоторое жизненное значение. Но сновидение есть следовое и притом большее частью давнее раздражение, а внушение - наличное. Далее, гипноз представляет меньшую степень торможения, чем сон, следовательно, внушение вдвойне по раздражающей силе больше сновидения. И, наконец, внушение как раздражение - коротко, изолированно и цельно, а потому и сильно; сновидение обыкновенно представляет цепь разнообразных и противоположнох следовых раздражений. Факт, что гипнотизируемому можно внушить все противоположное действительности и вызвать реакцию, прямо противоположную действительным раздражениям: сладкий вкус вместо горького, необыкновенное зрительное раздражение вместо самого обыкновенного, и т. д., без натяжки можно бы понять как парадоксальную фазу в состоянии нервной системы, когда слабые раздражения имеют больший раздражающий эффект, чем силь ные. Реальное раздражение, например от сладкого вещества, идущее прямо в соответствующую нервную клетку, надо думать, больше сравнительно с раздражением словом "горький", переходящим из соответствующей звуковой клетки в клетку, отвечающую реальному раздражению горьким, как условный раздражитель первого порядка всегда сильнее условного раздражителя второго порядка. Парадоксальная фаза, может быть, имеет более широкое значение, чем только указанное ранее, и теперь в патологических случаях. Можно представить себе, что она же дает себя знать и в тех нормальных людях, которые больше поддаются влиянию слов, чем фактам окружающей действительности. Возможно, что мы когда-нибудь научимся делать внушение и животным во время гипнотического состояния. Факт, что известные фазы гипнотического состояния у челсвека остаются более или менее стационарными, повторяется и у собак. Точно так же у людей, как и у животных, при известных внешних условиях и в зависимости от индивидуальности нервной системы, гипнотическое состояние более или менее скоро переходит в полный сон. В определенной связи с гипнотическим состоянием стоит пассивно-оборонительный рефлекс. Как уже сказано раньше, давнюю форму гипноза животных можно с основанием рассматривать как пассивно-оборонительный рефлекс, состоящий при встрече с чрезвычайным или сильным внешним раздражением в большем для меньшем обездвижении животного путем задерживания (развивающегося прежде всего в коре полушарий) скелетно-двигательной системы. Этот рефлекс многократно обнаруживался у наших экспериментальных животных в различных степенях интенсивности и в несколько разных формах, неизменно удерживая свой основной задерживающий характер. Вариации его состояли как в большем или меньшем ограничении движений животного, так и в ослаблении и исчезании его условных рефлексов. Пассивнооборонительный рефлекс обыкновенно вызывали чрезвычайные и сильные внешние раздражения. Но чрезвычайность и сила внешних раздражений, конечно, совершенно относительные величины. Чрезвычайность определяется тем, чему животное подвергалось ранее, а сила действия внешнего раздражения зависит от состояния данной нервной системы: от ее врожденных свойств, от здоровья или болезни ее, наконец, от различных стадий здорового существования. Все это мы видели на наших собаках. Собаки, выставляемые ранее много раз перед многолюдной аудиторией, оставались перед ней, наконец, совершенно нормальными; впервые перед ней появляющиеся приходили в сильно тормозное состояние. Ранее описанная исключительная собака Умница относилась к малейшим колебаниям среды как к сильным раздражениям и чрезвычайно ими тормозилась. Некоторые собаки, подвергшиеся сильному влиянию чрезвычайного наводнения и, очевидно, впавшие в хроническое болезненное состояние, тормозились сильными условными раздражителями, которые раньше на них такого действия не имели. Наконец некоторые собаки представляют такое тормозимое состояние только при известных степенях гипнотического состояния. Последний случай производит на наблюдателя чрезвычайное впечатление. Вот собака, которая в обстановке наших опытов раньше оставалась все время бодрой, быстро и жадно ела подаваемую ей после условных раздражителей пищу. Частым и сплошным применением слабых условных раздражителей мы привели ее в известную степень постоянного гипнотического состояния в опытной обстановке, собака сделалась малоподвижной. Теперь наступает следующее странное положение дела. Мы применяем сильные условные раздражители; собака, не раз повернувшись к месту, где подается еда, затем резко отворачивается в сторону и к пище не прикасается. Всякий, смотря на собаку, непременно скажет, что собака чего-то боится. Пустим в ход слабый условный раздражитель -- собака сейчас же подходит к подаваемой кормушке и спокойно ест. Рассеем, устраним гипнотическое состояние, и все условные раздражители дают нормальный эффект. Ясно, что в особом состоянии животного прежние обычные раздражители действуют каень сильные и производят тормозной рефлекс. И наоборот, на нашей исключительно тормозной собаке Умнице, как только некоторыми приемами нам удавалось несколько поднять раздражительный тонус ее полушарий, сейчас же можно было наблюдать значительное ослабление ее иначе почти беспрерывноео пассивно-оборонытельного рефлекса. Во всех только что приведенных случаях всегда бросаются в глаза характерные пассивно-оборонительные позы животного. Когда перед вами проходят все эти опыты, нельзя не прийти к заключению - и оно должно быть признано правильным, по крайней мере во многих случаях, - что то, что психологически называется страхом, трусостью, боязливостью, имеет своим физиологическим субстратом тормозное состояние больших полушарий, представляет различные степени пассивно-оборонительного рефлекса. Конечно, после этого является вполне оправданным рассматривать бред преследования и фобии как естественный тормозной симптом болезненно ослабленной нервной системы. Есть формы страха и трусости как паническое бегство и специальные рабские позы, которые, повидимому, противоречат высказанному заключению, что в основании их лежит тормозной процесс. Надо принимать, что это безусловные рефлексы, исходящие от центров, расположенных прямо под большими полушариями, и вступающие в деятельность именно при заторможенности больших полушарий. Последнее доказывается исчезанием при этом условных рефлексов. Несколько слов по поводу наших опытов, описанных в конце предшествующей лекции. Если они при повторениях и вариациях найдут полное подтверждение, то ими, может быть, будет брошен некоторый свет на темные явления нашего субъективного мира, касающиеся отношений между сознательным и бессознательным. Эти опыты показывали бы, что такой важный корковый акт, как синтезирование, может совершаться и в частях полушарий, находящихся в известной степени торможения под влиянием преобладающего в коре в данный момент сильного раздражения. Пусть этот акт тогда не сознается, но он произошел и при благоприятных условиях может обнаружиться в сознании готовым и представляться как возникший неизвестно как. В заключение всех этих лекций повторяю, что все наши опыты, как и подобные опыты других авторов, направленные к чисто физиологическому анализу высшей нервной деятельности, я рассматриваю как первую пробу, которая, однако, по моему глубокому убеждению, вполне оправдала себя. Получилось неоспоримое право сказать, что исследование чрезвычайно сложного предмета вышло, таким образом, на настоящую дорогу и ему предстоит, конечно, не близкий, но полный успех. Про себя мы можем сказать, что теперь перед нами гораздо больше вопросов, чем сколько их было раньше. Прежде по необходимости мы искусственно упрощали, схематизировали предмет. Теперь же, при некотором знании общих основ его, нас обступают, вернее сказать - подавляют, массы частностей, требующих детерминизации.Список печатных трудов сотрудников автора [2]


















 [1] 1 Дата указана ошибочно: первое и второе издания вышли в свет в 1927 г. -- Ред. 5 И. П. Павлов, Собр. соч. , т. IV
[2] 2 Ло 1927 г. включительно
[1] 1 Дата указана ошибочно: первое и второе издания вышли в свет в 1927 г. -- Ред. 5 И. П. Павлов, Собр. соч. , т. IV
[2] 2 Ло 1927 г. включительно
Последние комментарии
12 часов 38 минут назад
12 часов 38 минут назад
12 часов 47 минут назад
12 часов 55 минут назад
13 часов 53 минут назад
14 часов 11 минут назад