Генеалогия нейронов [Дмитрий Антонович Сахаров] (fb2) читать онлайн
- Генеалогия нейронов 1.65 Мб, 253с. скачать: (fb2) - (исправленную) читать: (полностью) - (постранично) - Дмитрий Антонович Сахаров
[Настройки текста] [Cбросить фильтры]
[Оглавление]
1. ВВЕДЕНИЕ
Современной физиологией прочно установлено, что клеточная популяция нашей нервной системы имеет весьма пёстрый состав. То, что на периферии существуют разные медиаторные механизмы, было известно давно; вслед за тем обнаружилось, что разнообразие химических синапсов и, следовательно, качественное разнообразие самих нейронов ещё более характерно для головного мозга. Задача этой книги — показать, что причину медиаторных особенностей того или иного нейрона следует искать в его родословной. Здесь будет рассмотрена гипотеза, состоящая в том, что наша нервная система представляет собой сборное клеточное образование, что в ней взаимодействуют нервные клетки, которые не идут из одного корня. Сам по себе генеалогический подход, связывающий специфические особенности клеток и тканей с историей их развития, не нов. Большая заслуга в теоретическом обосновании такого подхода принадлежит Н. Г. Хлопину, который, однако, считал его не приложимым к нервной ткани [66]. Развиваемое в этой книге представление о генетической разнородности нервных клеток можно рассматривать как дальнейшую конкретизацию нейронной теории. Субстрат нервных процессов не только прерывен (в чём уже нет сомнений), но и представлен качественно разными клеточными звеньями. Множественное происхождение нейронов, которое для удобства будет далее называться полигенезом, мыслится как главная, но не единственная причина разнообразия медиаторных механизмов. Предполагается, что на протяжении процесса эволюции нейроны, сохраняя исходный тип химизма, не оставались неизменными; внутри каждой клеточной линии могла иметь место дивергентная дифференциация, которая ещё больше увеличивала названное разнообразие. В последней главе читатель найдёт обсуждение этого, а также некоторых других проявлений эволюции медиаторов. И всё же, отдавая должное эволюции нервной системы, предлагаемое представление относит возникновение клеточной разнородности к исходному пункту этой эволюции. Тем самым точка зрения автора отличается от распространённого мнения, что химические синапсы становились разными по мере того, как всё более усложнялись функции нервной системы и всё более специализировались составляющие её нейроны. Нужно, однако, отметить, что такое мнение никем детально не рассматривалось и не аргументировалось — имелось, по-видимому, в виду, что это само собой разумеется. Каких-нибудь полтора-два десятилетия назад проблемы не существовало — никто не задавался вопросом о причине множественности медиаторных механизмов, поскольку не был ещё известен сам феномен. Существование химических синапсов в центральной нервной системе оставалось недоказанным, а наличие двух медиаторных механизмов — холинергического и адренергического в периферических окончаниях объясняли тем, что один антагонистичен другому. Развитие микроэлектродных и электронно-микроскопических исследований убедило физиологов, что концепция химических посредников приложима к центральным синапсам. Но почти одновременно с этой победой химическая теория синаптической передачи стала терять свою логическую стройность: не оправдались ожидания тех, кто надеялся найти некий «центральный передатчик», а также и тех, кто думал, что в мозге, как на периферии, работают два антагонистических медиатора. Медиаторов оказалось много. Ещё не будучи осознанными теоретически, факты о множественности медиаторных веществ привлекли пристальное внимание медиков, которые обоснованно усмотрели в этом новые возможности понять природу ряда нервных и психических заболеваний и найти способы их лечения. Можно даже говорить о первых успехах в этом направлении. Тем настоятельнее встаёт задача разобраться в нейробиологической стороне вопроса — оценить медиаторное разнообразие, его природу и смысл. Читателю уже в общих чертах известно решение, предлагаемое автором этой книги. Наверное, к такому решению можно было прийти разными путями. Путь, которым шёл автор, определён его личными пристрастиями и некоторыми обстоятельствами необязательного характера. Хотелось бы назвать два момента, наложивших на книгу несомненный отпечаток. Во-первых, полное предпочтение отдаётся здесь сравнительному методу исследования. Такой подход, вообще, не очень распространен, но он вполне традиционен для отечественной физиологии, в которой прочно утвердилась идея, что для понимания функции важно знать историю её развития (Л. А. Орбели, X. С. Коштоянц, А. Г. Гинецинский и многие другие). Хачатур Сергеевич Коштоянц (1900-1960), под руководством которого автор начинал работать в нейробиологии, особенно настойчиво подчёркивал важность исторического подхода при анализе химической основы нервной деятельности и много сделал для развития сравнительных исследований в этой области. Накопленный эволюционной физиологией опыт приложим к разным конкретным проблемам, в том числе и к нашей: ведь только в общем виде ясно, что разнообразие медиаторов является результатом исторического развития нервной системы, но без специального разбора не понять закономерностей этого процесса. Второй субъективный момент — место, занимаемое в книге брюхоногими моллюсками. Бесспорно, у гастропод, имеющих особенно крупные нейроны, состав нейронных популяций и клеточные механизмы передачи изучены полнее, чем у многих других животных. Но в действительности имели значение не эти объективные, а личные причины, — то, что к постановке названных вопросов и попыткам понять природу химической мозаики, наблюдающейся в нервной системе млекопитающих, автор пришел в ходе исследования нейронов брюхоногих моллюсков. Что касается мотивов, побудивших заняться изучением нервных клеток гастропод, то их прекрасно сформулировал много десятилетий назад другой исследователь — Н. П. Вагнер. Трудно удержаться и не процитировать нескольких фраз: «При первом взгляде на узлы нервной системы клиона каждый наблюдатель наверное будет поражён громадной величиной их клеток… При взгляде на эту громадную величину… мне пришло на мысль исполнить давнишнее желание и разобрать хоть у одного беспозвоночного типа вполне весь комплекс нервной системы. Такой разбор, по всей вероятности, повёл бы к объяснению, хотя гадательному, многих функций нервной системы у большей части, если не у всех, беспозвоночных животных. Правда, мне хотелось сделать эту работу без особого труда, и прозрачность, или, так сказать, откровенность нервной системы клиона давала мне в этом случае надежду на успех [7, стр. 122]. Цитата взята из монографии Н. П. Вагнера «Беспозвоночные Белого Моря», напечатанной в 1885 г. Профессор Петербургского университета Николай Петрович Вагнер создал при Соловецком монастыре первую в России морскую биологическую станцию, где и выполнил великолепное для своего времени нейробиологическое исследование клиона — крылоногого моллюска, называемого также морским ангелочком (Clione limacina). Прошло три четверти столетия, и в роли такого наблюдателя, бросившего «первый взгляд на узлы нервной системы клиона», вдруг довелось оказаться мне. Наверное, эта история повторится ещё не раз, и каждого нового наблюдателя заново поразит сверкание окологлоточного ожерелья и нагая зримость нейронов, которую Вагнер весьма точно назвал «откровенностью». Кстати, и слово ожерелье, которое непосвящённому покажется художественной вольностью, во времена Вагнера служило рутинным анатомическим термином. Книгу Вагнера, впрочем, я нашёл и прочитал лишь через несколько лет после встречи с ганглиями клиона. Назвать эту встречу случайной было бы неверно; к поискам в этом направлении побуждали блестящие успехи французских авторов — А. Арванитаки, Н. Халазонитиса, Л. Тауца, которые в середине 50-х годов обогатили клеточную физиологию новым замечательным объектом — гигантскими нейронами аплизии. Клион вселял надежду на то, что объекты, сравнимые по своим достоинствам с недоступной аплизией, могут быть найдены в нашей фауне. В самом деле, за работой, выполненной на ганглиях клиона [49], последовало обнаружение гигантских нейронов у беломорских голожаберных [50], затем — ещё более крупных — у дальневосточной тритонии [8], и в результате работа на нейронах гастропод стала для автора главным занятием. Нужно признаться, что в течение нескольких лет изучение нервных клеток моллюсков не было связано с какой-то определённой задачей или проблемой. Интерес к объекту носил общий характер, и я стремился узнать об этих клетках как можно больше. В ряду других исследований проводилось картирование нейронов, различающихся по физиологическим или химическим характеристикам. В ходе работы было замечено, что клетки с одинаковыми наборами свойств можно найти у разных видов гастропод — даже у видов, весьма далёких друг от друга во всех отношениях. Это наблюдение вызвало специальный интерес и повлекло вывод о существовании гомологичных нейронов и о консервативности их специфического химизма [52, 53]. Отсюда было уже недалеко до гипотезы полигенеза [54, 279]. Эта гипотеза рассматривается в книге двояко: и «в чистом виде» и в связи с тем конкретным моллюсковым материалом, который привел к постановке связанных с ней вопросов и подсказал решение. Читатель, которого не интересует эта вторая сторона, может пренебречь главами 4 и 5, ограничившись «Заключением» к каждой из них. В собственные результаты, на которые я опираюсь, вложены опыт, знания и труд многих людей. В первую очередь я должен здесь назвать коллектив лаборатории физиологии им. X. С. Коштоянца Института биологии развития АН СССР, возглавляемый членом-корреспондентом АН СССР Т. М. Турпаевым. Я особенно благодарен за ценное сотрудничество Г. Н. Коробцову, С. Н. Нистратовой и Н. К. Остроумовой (Чернопятовой). Ряд работ выполнен совместно с коллегами из Института биологической физики АН СССР (В. Л. Боровягин, Б. Н. Вепринцев, И. В. Крастс), Научно-исследовательского института неврологии АМН СССР (А. В. Сахарова) и Биологического института АН Венгрии (Я. Шаланки, И. Ж.-Надь) — всем им выражаю самую сердечную признательность. Свою благодарность я хотел бы выразить и тем, от кого получал критику и советы в период работы над рукописью: Г. А. Бузникову, Н. Н Демину, П. Г. Костюку, Л. Г. Магазанику, М. Я. Михельсону, Н. А. Смиттен, Т. А. Сперанской и в особенности Т. М. Турпаеву, взявшему на себя редактирование книги.2. НЕОДНОРОДНОСТЬ КЛЕТОЧНОГО СОСТАВА НЕРВНОЙ СИСТЕМЫ МЛЕКОПИТАЮЩИХ
2. 1. Терминология
Прежде всего следует определить некоторые основные понятия и исходные позиции. Даже если читатель не разделяет этих определений, он по крайней мере будет знать язык, которым пользуется автор. Речь будет идти о химических различиях между нейронами, но интересовать нас будут не любые особенности химизма, а лишь те, которые имеют отношение к продукции физиологически-активного начала, выделяемого из аксонных окончаний. Очевидно, что нейрон, секретирующий глутамат, должен отличаться от, скажем, дофаминергического нейрона. Эту сторону химизма физиологи нередко называют «эргичностью», но многими такой термин воспринимается как жаргонизм, хотя окончание «ергический», введенное в 1933 г. Дейлом [142], вошло в литературу прочно. Взамен «эргичности» мы будем пользоваться термином медиаторная специфичность. Нужно, однако, учитывать, что и этот термин не вполне удовлетворителен, поскольку рассматриваемая сторона химизма обеспечивает продукцию не только медиаторов, но и нейрогормонов. Следовательно, говоря о медиаторной специфичности, мы вкладываем в понятие «медиатор» расширительный смысл. В точном смысле под медиатором (синонимы: синаптический передатчик, нейтротрансмиттер) понимается, как это принято в литературе, физиологически-активное вещество, которое секретируется из возбужденного эфферентного (эффекторного) нервного окончания и диффундирует к мишени. Предполагается, следовательно, что мишень находится где-то рядом и что она обладает чувствительностью к передатчику, т. е. соответствующими рецепторами. Бывает, однако, и так, что снабжённая рецепторами клеточная мишень находится на значительном удалении от аксонных терминалей. В этом случае их активный агент доносится до мишени кровью (или гемолимфой), и его удобно называть нейрогормоном. Итак, действующее начало аксонных терминалей можно классифицировать в зависимости от его химической природы, и тогда мы говорим о медиаторной специфичности нейрона. (Подразумевается, что все аксонные терминали одного нейрона секретируют одно и то же активное начало, — в этом состоит известный принцип Дейла, который за несколько десятилетий своего существования не встретил сколько-нибудь обоснованных возражений). С другой стороны, секретируемые активные агенты можно группировать в зависимости от способа их доставки к месту назначения, и тогда мы делим их на медиаторы и нейрогормоны. Естественно, что в роли медиатора и нейрогормона (и даже просто гормона) может выступать одно и то же вещество, а клетки одной и той же медиаторной специфичности могут оказаться в роли как «обычных» нейронов, образующих синаптические контакты, так и нейросекреторных клеток, т. е. нейронов, выделяющих активное начало в жидкую среду организма. Такое содержание рассмотренных понятий кажется нам более рациональным, чем встречающееся нередко в литературе, когда понятия нейросекреции и нейрогормона жестко связывают с определённой химической природой продукта секреции или когда нейросекреторным клеткам отказывают в праве называться нейронами только на том основании, что рядом с их аксонными терминалями не оказалось соответствующей мишени.
2. 2. Медиаторная специфичность нейрона и идентификация медиатора
Медиаторная специфичность нейрона выражается комплексом признаков, определяемых природой самого медиатора. В клетках разной специфичности неизбежно различны ферментные системы, ведущие синтез медиатора. Так, в нейронах, секретирующих ацетилхолин, высока активность холинацетилазы, чего нет, допустим, в нейронах, секретирующих норадреналин и дофамин. Клетки двух последних типов, хотя и очень близки в ряде отношений, различаются между собой тем, что в первых (норадренергических) имеется фермент дофамин-бета-оксидаза, не нужный вторым (дофаминергическим). Различны, в зависимости от природы медиатора, и вещества, из которых построены секреторные органеллы цитоплазмы нейронов. Так, в состав секреторных гранул в нервных клетках, выделяющих катехоламины, входят белки хромогранины, а в нейронах, выделяющих физиологически активные октапептиды, — белки нейрофизины. Различия в химическом составе медиатора и сопутствующих ему макромолекул находят проявление в строении некоторых структур цитоплазмы, в частности, секреторных пузырьков и гранул, которые неодинаковы по своим морфологическим характеристикам в разных нейронах. Наконец, неодинаковы системы избирательного накопления медиаторных веществ из внеклеточной среды. Этими системами обеспечивается реутилизация выделившегося из клетки медиатора, в связи с чем они работают с высокой специфичностью. Благодаря тому что каждый тип медиаторной специфичности выражается совокупностью характерных свойств химизма и, отчасти, строения, работа по установлению типа специфичности у исследуемых нейронов и синапсов в значительной степени облегчается. Её, в принципе, можно вести, используя широкий арсенал методов, которые всё больше становятся реально доступными; в их числе цитохимическое обнаружение самих медиаторных веществ, определение активности ферментов синтеза медиаторов, иммунохимическая локализация этих ферментов или специфических структурных белков, ультрамикроскопическое исследование секреторных органелл, изучение систем захвата экзогенных медиаторов или их аналогов, и т. д. Применение каждого из этих приёмов связано, конечно, со своими трудностями, но всё же на этой основе можно идентифицировать тип нейрона или синапса. Значительно труднее работа по химической идентификации неизвестных медиаторов. Вопрос о критериях, которые при этом должны выполняться, неоднократно обсуждался в литературе, и разными авторами публиковались разные списки таких критериев, Р. Верман справедливо заметил недавно, что для отождествления передатчика достаточно, чтобы были удовлетворены всего лишь два критерия: критерий накопляемости и критерий идентичности действия. Смысл первого заключается в том, что вещество, предполагаемое медиатором, должно при физиологических для данного синапса условиях выделяться из пресинаптической структуры в ответ на пресинаптическое раздражение в количестве, соответствующем количеству стимулов. По второму критерию, предполагаемый медиатор должен действовать на постсинаптическую структуру, используя те же молекулярные механизмы, которые обеспечивают эффект самого естественного передатчика [340]. Длинные списки критериев возникли, по мнению Вермана, потому, что достаточность двух указанных критериев не всегда осознается, и главным образом потому, что применить эти два критерия совсем не просто. В последние годы выяснилось, что некоторые критерии, которые до середины 60-х годов считались чуть ли ни главными, нужно признать ошибочными. Исторически случилось так, что в первые десятилетия изучения медиаторного процесса объектом исследований служили периферические холинергические соединения позвоночных (окончания блуждающего нерва на сердце и моторных волокон на скелетных мышцах). В ходе этих исследований развились общие представления о химическом синапсе, в том числе убеждение, что химический синапс должен быть обеспечен ферментом, инактивирующим передатчик. Из «критерия инактивирующего энзима» вытекал как следствие ещё один критерий: эффект вещества, предполагаемого медиатором, как и эффект раздражения пресинаптических волокон, должен усиливаться при применении фармакологических агентов, ингибирующих указанный фермент. Эти критерии следует признать неудовлетворительными. Ярче всего их ограниченность выявилась при изучении синапсов, использующих в качестве медиатора иные, чем ацетилхолин, вещества. Как теперь считают, синаптическое действие катехоламинов прекращается главным образом благодаря обратному захвату их нервным окончанием из синаптической щели. Сходный механизм предполагается для синапсов, передачу в которых осуществляет глицин, гамма-аминомасляная кислота и некоторые другие медиаторы [см. 194]. Прекращение действия синаптического серотонина некоторые авторы связывают с десенситизацией рецепторов к передатчику [168], однако на беспозвоночных и для серотонинергических волокон показан избирательный захват медиатора [266]. Сравнительное изучение холинергических соединений также привело к пересмотру существовавшего ранее представления об обязательном участии синаптической холинэстеразы в медиаторном процессе, осуществляемом ацетилхолином. Синаптическое действие этого передатчика может быть остановлено, по-видимому, по крайней мере тремя разными способами: диффузией из синаптической щели, инактивацией холинорецептора и, наконец, энзиматической инактивацией самого ацетилхолина. Это разнообразие механизмов, с одной стороны, выражает собой процесс эволюционного совершенствования синапсов, а с другой, способ, посредством которого определяются функциональные параметры передачи. Подробнее этот вопрос рассмотрен нами, совместно с Турпаевым, в специальных статьях [65, 285, 317]; отчасти, об этом будет идти речь ниже в связи с обсуждением вопроса об эволюции нервных клеток (7.3.). С учетом данных, не укладывающихся в существовавшую ранее схему медиаторного процесса, вопрос о критериях идентификации медиатора рассмотрен в нескольких работах [337, 6, 242, 337, 339, 340]. В общем, мнения их авторов во многом совпадают. Признается ценным для периферических окончаний один из старых критериев: при раздражении пресинаптических волокон предполагаемый передатчик должен обнаруживаться во внеклеточной среде в области синапса. Хотя практические трудности ограничивают приложимость этого критерия к синапсам ЦНС, иногда удается их преодолевать, применяя высокочувствительные методы обнаружения малых количеств медиатора. Несколько критериев выдвигаются в качестве более универсальных. Во-первых, вещество, предполагаемое медиатором, должно в достаточных количествах иметься в пресинаптическом нейроне, с преимущественной локализацией в окончаниях аксона. Во-вторых, здесь же должна присутствовать энзиматическая система для синтеза этого вещества или для обеспечения окончаний медиатором каким-то иным способом. В-третьих, постсинаптическая мембрана должна обладать специфической чувствительностью к предполагаемому медиатору. В-четвёртых, при нанесении предполагаемого медиатора на постсинаптическую структуру он должен воспроизвести все эффекты синаптическо-го действия. В-пятых, фармакологические агенты, влияющие на постсинаптические эффекты естественного передатчика, должны сходным образом влиять на эффекты предполагаемого медиатора при его искусственном нанесении на постсинаптическую структуру. Почти все эти критерии могут приниматься с известными оговорками, и к ним нельзя относиться некритически. Так, нет полной уверенности в том, что специфические рецепторы, на которые действует медиатор, должны обязательно находиться на постсинаптической мембране: между медиаторами и гормонами нет строгой границы, а среди гормонов известны такие, которые взаимодействуют с внутриклеточными рецепторными структурами. Больше всего оговорок высказано по поводу четвёртого и пятого из упомянутых критериев. Здесь исследователь встречается с некоторыми осложнениями принципиального характера. Техника многоканальных микроэлектродов, используемая в таких экспериментах, позволяет апплицировать испытуемое вещество на тело нейрона, внутрь которого введен отводящий микроэлектрод. Синаптические же окончания и сами рецепторы к медиатору нередко располагаются на значительном расстоянии от тела нейрона — например, на удалённых частях дендритов. В таких ситуациях испытуемое вещество, в точности идентичное медиатору, не будет давать эффекта, вполне идентичного эффекту раздражения пресинаптических волокон: например, неодинаковыми окажутся потенциалы равновесия. Специальные исследования этих трудностей показывают, что они преодолимы, но важно не упускать их из вида. Мак-Леннан в последнем издании своей книги «Синаптическая передача» [242] предлагает включить в число критериев развитие денервационного повышения чувствительности к веществу — кандидату в передатчики. В самом деле, в некоторых синаптических структурах чувствительность к медиатору повышается после денервации на несколько порядков. Но нельзя забывать о том, что известны и противоположные ситуации. Так, клетки членистоногих вообще теряют чувствительность к гамма-аминомасляной кислоте после перерезки тормозных волокон, выделяющих на них этот медиатор; рецепторы к медиатору снова восстанавливаются после регенерации иннервирующих волокон [см. ссылки в 279]. Применение указанных критериев должно быть комплексным, только в этом случае можно с уверенностью решать вопрос о медиаторной роли того или иного вещества. Поучительна в этом отношении история изучения передачи в окончаниях симпатических нервных волокон. Сейчас общепризнанно, что передачу в них осуществляет, по крайней мере у теплокровных, норадреналин. Но ещё в начале 60-х годов, несмотря на наличие убедительных фактов, продолжались споры о месте, из которого высвобождается норадреналин при возбуждении симпатических нервов: нельзя было доказательно утверждать, что амин изливается из самих нервных окончаний, а не из внешней по отношению к ним структуры, например, из хромаффинных клеток. Как подчеркнул известный шведский фармаколог А. Карлссон, недостающий критерий удалось выполнить лишь благодаря появлению гистохимического метода Фалька и Хилларпа [111]. Вообще, нужно отметить, что если ещё не так давно вопросом об идентификации медиаторов и установлении типов медиаторной специфичности интересовались только физиологи, то сейчас в решении этих вопросов очень велика стала роль информации, получаемой с помощью морфологических [см. 180] и микрохимических методов исследования [162, 169, 240 - 241а, 259, 263, 330, 331 и др.], и эти методические подходы к медиаторным проблемам представляются сегодня наиболее перспективными.
2. 3. Типы нейронов у млекопитающих
Если теперь с точки зрения медиаторной специфичности рассмотреть клеточную популяцию нервной системы млекопитающих, то она представится в виде мозаики, своеобразный рисунок которой может показаться прихотливым. С одной стороны, не видно закономерной связи между этой мозаикой и функциональной организацией нервной системы. С другой, распределение химически специфичных нейронов не следует за анатомическим членением нервной системы: в одном и том же её отделе нейронный состав, как правило, разнороден. Нужно добавить, что многие группы нейронов остаются ещё неизученными, принадлежность других к тому или иному химическому типу остается предположительной. Но эти белые пятна довольно быстро заполняются. Ниже перечислены выявленные к настоящему времени типы нейронов, различающиеся характером секретируемого физиологически активного продукта, и указано, где они располагаются. Из большой литературы вопроса выбраны для ссылок обзорные или наиболее важные работы.
2. 3. 1. Холинергические нейроны
Холинергическими называют нервные клетки, эффекторными окончаниями которых секретируется ацетилхолин. Впервые медиаторная функция была доказана именно для этого вещества [235]. Для нейронов, аксонные окончания которых находятся вне ЦНС, удаётся с достаточной полнотой выполнить систему критериев, позволяющих идентифицировать синаптический передатчик с ацетилхолином. Это позволило ещё в 30-х годах отнести к холинергическим следующие типы нейронов: 1) мотонейроны, дающие окончания на скелетных мышцах; 2) симпатические и парасимпатические преганглионарные нейроны и 3) парасимпатические постганглионарные нейроны. Проведённые в последующие годы исследования медиаторной функции ацетилхолина в окончаниях этих нейронов отличаются большой полнотой и тщательностью [см., например, 15, 62, 70, 191]. Гораздо больше трудностей встретилось на пути изучения холинергических интернейронов. Хотя несомненно, что в ЦНС представлены, наряду с другими, и холинергические окончания (достаточно напомнить о существовании синапсов между аксонными коллатералями спинальных мотонейронов и клетками Реншоу), до сих пор сохраняется значительная неясность относительно удельного веса и локализации внутрицентральных холинергических систем. Несмотря на усилия многих исследователей, применение микроэлектродных методов принесло довольно бедные результаты. С достаточной обоснованностью можно сейчас говорить о наличии холинергических нейронов в составе ретикулярной формации (в частности, в стволовой части среднего мозга), откуда начинаются волокна, идущие к холинореактивным структурам коры большого мозга, боковых коленчатых тел и некоторых других участков мозга, а также о холинергической природе некоторых таламических проекций в первичную сензорную кору. Помимо гистохимических данных и демонстрации идентичности эффектов ацетилхолина и пресинаптического возбуждения, удалось показать, что при возбуждении этих структур ацетилхолин выходит во внеклеточное пространство [114, 244, 202, 126]. Мнение некоторых авторов о холинергической природе нейронов нижней оливы, дающих начало лиановидным волокнам мозжечка, оспаривается другими авторами, которые, в свою очередь, считают холинергическими некоторые клетки моста среднего мозга, дающие начало части мшистых волокон мозжечка. Данные о холинореактивности многих клеток ЦНС не подкреплены какими-либо сведениями об источниках волокон, возможно, дающих на этих клетках холинергические окончания. Подробнее см. об этом у Мак-Леннана [242] и Филлиса [268]. Крайне медленный прогресс в работе по обнаружению холинергических клеток головного мозга объясняется методическими трудностями. В течение многих лет идентификацию холинергических структур связывали с гистохимическим выявлением активности ацетилхолинэстеразы. Простота этого подхода оказалась кажущейся. Только в редких и специальных случаях высокая активность ацетилхолинэстеразы наблюдается в нейронах, секретирующих ацетилхолин, — значительно чаще, но тоже не всегда, фермент располагается в структуре, на которую действует медиатор, т. е. за синаптической щелью. Далее, имеется огромная литература о выявлении ацетилхолинэстеразы в таких соединениях, где ни пре-, ни постсинаптическая клетки не являются холинергическими. Наконец, накопилось немало сведений о том, что присутствие этого фермента в нейронах вообще может не иметь отношения к синапсам и синаптической передаче. Всё это обязываеткрайне критически относиться к обширной литературе о «холинергических нейронах», идентифицированных столь ненадёжным способом. Несравненно ценнее для идентификации сведения об активности ключевого фермента синтеза ацетилхолина — холинацетилазы (ацетилтрансферазы холина). Сделаны первые шаги в разработке методов гистохимического обнаружения этого фермента. Они ещё несовершенны, но во всяком случае специфичны: так, у крысы продукт гистохимической реакции выявляется в холинергических спинальных мотонейронах, но его нет в нехолинергических клетках спинальных ганглиев [см. 180]. Можно рассчитывать, что дальнейшее развитие гистохимии холинацетилазы, а также использование и совершенствование методов микрохимического определения активности этого фермента [169, 241] и содержания ацетилхолина в отдельных нейронах [241а] позволят уверенно локализовать холинергические нейроны и их связи. В синапсах, для которых установлена медиаторная функция ацетилхолина (моторные окончания на скелетных мышцах позвоночных, парасимпатические окончания в сердце и др.), секреторные органеллы относятся к одной и той же категории: это не имеющие плотного содержимого округлые пузырьки диаметром около 450-500 Å. Для дифференцирования между пузырьками, содержащими ацетилхолин, и внешне похожими на них пузырьками в окончаниях другой специфичности (в частности, в секретирующих глутамат) важно было бы обладать методом выявления самого ацетилхолина. Однако попытки, предпринимаемые в этом направлении, пока не увенчались успехом. Акерт и Сандри в 1968 г. дали повод надеяться на то, что такой метод найден, когда им удалось импрегнировать содержимое прозрачных пузырьков в нервно-мышечных и центральных синапсах смесью иодистого цинка с четырёхокисью осмия [75], но последующие исследования показали, что реакция выявляет не ацетилхолин и природа её избирательности неясна [см. 74]. Между тем из ацетилхолин-содержащих синаптических пузырьков, полученных из электрических органов рыб, выделили особый кислый белок, везикулин, в связи с чем было высказано предположение, что именно этот белок связывает цинк, обеспечивая электронно-микроскопическую импрегнацию полости синаптических пузырьков [345]. Если в самом деле метод выявляет кислые белки, то специфичность его не может быть очень высока. Внутри синаптических пузырьков ацетилхолин вряд ли находится в растворенном виде, — скорее, он связан отрицательно заряженными противоионами, роль которых, возможно, играют упомянутый везикулин и АТФ [344, 345]. В холинергических окончаниях, наряду с прозрачными секреторными пузырьками, в гораздо меньшем числе встречаются гранулы диаметром около 800 Å, имеющие плотное содержимое. Они как будто не принимают участия в процессе секреции. Эти более крупные гранулы, в отличие от секреторных пузырьков, обнаруживаются не только в пресинаптическом окончании, но и в теле нейрона, а также по ходу аксона. Существует предположение, что в таких гранулах, образующихся в элементах аппарата Гольджи в теле холинергического нейрона, в аксонные окончания доставляются какие-то макромолекулы, необходимые для производства секреторных пузырьков [см. 295]. Итак, для обнаружения холинергических нейронов и синапсов применяют комплекс методов. В микроэлектродных исследованиях широко используются фармакологические агенты, в частности, вещества, влияющие на синтез, секрецию и рецепцию ацетилхолина. Гистохимики возлагают сейчас свои надежды на холинацетилазу. Можно думать, что этот фермент, как и другие специфические белки холинергических нейронов, в частности, везикулин, скоро будут обнаруживать и иммуногистохимическими методами. Электронная микроскопия пока позволяет идентифицировать холинергические окончания лишь приблизительно: холинергические пузырьки трудно спутать с секреторными органеллами моноаминергических или пептидергических окончаний, но трудно и дифференцировать от прозрачных пузырьков какой-то другой медиаторной специфичности. Наконец, существуют основанные на разных принципах методы обнаружения рецепторов ацетилхолина [см. 39], что позволяет косвенно судить о холинергической природе волокон, иннервирующих такие холинореактивные структуры. всё это хорошо, но остро чувствуется потребность в более простых, прямых и надежных способах обнаружения холинергических структур. Ещё одно замечание. Начиная список медиаторных систем с холинергической, мы следовали традиции, которая всегда ставит ацетилхолин на первое место в ряду медиаторов. Это справедливо, если учитывать, что первые представления о механизме передачи в химических синапсах были получены благодаря открытию медиаторной функции ацетилхолина. Нужно, однако, не упускать из вида, что в реальных нервных системах ацетилхолин просто является одним из многих медиаторов и не имеет тех привилегий, которыми его наделила научная традиция. Об этом необходимо сказать потому, что особенное отношение к ацетилхолину явилось и продолжает являться психологической предпосылкой ошибочных генерализаций, которые лишь затрудняют изучение медиаторных систем. Одним из таких обобщений был уже упомянутый «критерий инактивирующего энзима». Немало сил было впустую потрачено на суету вокруг другой ложной идеи — а именно, что ацетилхолин должен обязательно принимать участие в работе нехолинергического нервного окончания, в частности, адренергического («концепция холинергического звена» Бэрна и Ранда [см. 39, 103]). Также и электронные микроскописты, увидев пустые пузырьки в нервном окончании, дружно вспоминают ацетилхолин — а почему бы, скажем, не глутамат, который тоже хранится в таких пузырьках? Наконец, холинорецептор почти монопольно владеет вниманием исследователей, занимающихся рецепцией медиаторных веществ. Важно не забывать, что этот крен в пользу ацетилхолина не имеет объективных оснований.
2. 3. 2. Моноаминергические нейроны
Этим названием объединяют нервные клетки, которые осуществляют свои синаптические эффекты при посредстве того или иного биогенного моноамина. Границы понятия «биогенные моноамины» (или «биогенные амины») в последние годы несколько сузились. Раньше сюда включали различные низкомолекулярные физиологически активные вещества, обладающие нейротропным действием и имеющие в своём составе аминогруппу, — в том числе и такие вещества, как гистамин, некоторые аминокислоты и т. д. После 1962 г., когда в гистохимическую практику вошел люминесцентный метод Фалька и Хилларпа, стало более принятым называть биогенными аминами те тканевые амины, которые выявляются этим методом, т. е. катехоламины и индолилалкиламины. Интерес к этим веществам как медиаторам нервных влияний существует давно и восходит к классическим опытам Отто Леви, показавшего на сердце лягушки, что симпатические нервные влияния, подобно парасимпатическим, осуществляются при помощи химического посредника [236]. Довольно долго, однако, этот интерес ограничивался симпатическими окончаниями и адреналином, который считался передатчиком в этих «адренергических» окончаниях. Заметным шагом вперед явились исследования Эйлера, который пришёл к заключению, что медиатором симпатических эффектов у млекопитающих является не адреналин, а норадреналин [151]. Следующей важной вехой явился 1954 год, когда в английском физиологическом журнале появились две работы, положившие начало изучению медиаторной роли биогенных аминов в мозге млекопитающих. В одной из них М. Фогт показала, что неравномерность распределения норадреналина в разных отделах мозга не позволяет согласиться с существовавшим мнением о функциональной связи этого амина с сосудами: Фогт обсудила предположение, что норадреналин может быть одним из синаптических передатчиков в ЦНС [322]. Другая работа носила сходный характер и описывала распределение в мозге серотонина [76]. Период медленного накопления фактов затянулся на несколько десятилетий, но ситуация резко изменилась в 60-х годах, когда биогенные амины оказались в центре внимания исследователей, занимающихся проблемой синаптической передачи. Несомненно, главной причиной, вызвавшей эту концентрацию усилий, явилось внедрение уже упомянутого гистохимического метода, который впервые сделал возможным прямое наблюдение медиаторных веществ [см. 129]. Применение метода Фалька и Хилларпа и основанных на нём других специальных методов (снятие спектральных характеристик свечения, сочетание люминесцентной гистохимии с радиоавтографией, с фармакологическими воздействиями и т. д.) позволило за короткий срок получить ответ на ряд важнейших вопросов. Были выяснены места расположения моноаминергических нейронов в центральной и периферической нервной системе и для каждой группы таких клеток определены области иннервации [см. 16, 77, 161, 319 и библиографию к этим работам; специально для мозга человека — 253]. Одновременно физиологами было показано, что возбуждение гистохимически идентифицированных моноаминергических систем ЦНС сопровождается выделением медиаторных аминов [93, 272, 293] и что аппликация этих аминов воспроизводит эффекты раздражения пресинаптических моноаминергических элементов [127, 351 и др.]. Люминесцентный метод стал основой больших успехов в создании фармакологических средств, избирательно действующих на моноаминергические нейроны, что уже нашло применение в клинике нервных болезней, в частности, в лечении паркинсонизма. Все большее развитие получает иммуногистохимия моноаминергических нейронов. В качестве антигенов, специфических для этих нейронов, используются ферменты, принимающие участие в синтезе медиаторов, хромогранины и другие специфические белки. Различия в наборе ферментов в клетках, секретирующих разные катехоламины, позволяют дифференцировать эти клетки с помощью иммунолюминесцентного метода [183, 184, 189]. Хорошие знания о рассматриваемых нейронах накоплены электронной микроскопией. Для клеток, секретирующих катехоламиновые медиаторы, характерно присутствие двух популяций пузырьков: мелких, диаметром около 400-500 Å, и более крупных. Первые наблюдаются только в области секреции, вторые — по всей длине аксона и в теле нейрона. И те и другие имеют электронноплотное центральное ядро, но при обычных условиях фиксации оно обычно сохраняется только в крупных пузырьках. Имеются хорошие основания считать, что плотное центральное ядро дают сами катехоламины в результате цепи реакций, которая начинается реакцией конденсации между амином и фиксирующим альдегидом. В самом деле, если фиксация проводится при условиях, уменьшающих возможность диффузии катехоламина из везикул, плотное ядро можно наблюдать и в больших, и в маленьких везикулах. Надёжное выявление плотного зерна в катехоламиновых везикулах обеспечивается фиксацией перманганатом [181, 182]. Недавнее исследование, проведенное на весьма чистой фракции крупных пузырьков (средний диаметр 763 Å), полученной из симпатического нерва, показало, что видимая в микроскоп плотность содержимого пузырьков коррелирует с содержанием в них норадреналина; авторы пришли также к заключению, что содержимое пузырьков сжимается в виде центрального или эксцентрично расположенного зерна лишь в результате фиксации, и нашли такие условия обработки, при которых содержимое остается равномерно распределенным в полости пузырька [314]. Дифференцирование между первичными катехоламинами, с одной стороны, и другими медиаторными биогенными аминами, с другой, основано на использовании разных фиксирующих альдегидов (глутаровый, муравьиный, акриловый), которые применяются в сочетании с тем или иным металлическим окислителем (перманганат, бихромат, четырёхокись осмия) [182, 349]. Электронная микроскопия широко использует также способность аминсодержащих нервных окончаний и секреторных везикул накапливать соответствующий амин, поглощая его из внеклеточной среды. Хотя специфичность таких систем захвата не абсолютна, указанное свойство позволяет локализовать пузырьки, содержащие амин, авторадиографическим методом [97]. Наконец, следует упомянуть о фармакологических агентах, которые, обладая химическим сходством с медиаторными аминами, способны лишать пузырьки их плотного содержимого (как, например, альфа-метилметатирозин) либо, наоборот, значительно повышать электронную плотность, накапливаясь в пузырьках в большом количестве (как, например, 5-оксидофамин). Вещества такого рода облегчают и делают более уверенной идентификацию моноаминергических клеток и волокон. Вместе с тем, обладая способностью разрушать нервные элементы, в которых они накапливаются, некоторые из таких веществ оказались полезным инструментом экспериментального изучения функций моноаминергических нейронов. Здесь в первую очередь нужно назвать 6-оксидофамин, поражающий норадренергические и дофаминергические нейроны [313], а также 5,6-диокситриптамин, оказывающий аналогичное действие на серотонинергические элементы [95]. Все упомянутые методы ультраструктурного анализа позволяют сделать вывод, что медиаторный амин в том или ином моноаминергическом нейроне содержится в обеих популяциях пузырьков. Выдвинута гипотеза, что пузырьки более крупного класса трансформируются в мелкие втерминальном участке аксона [218]. Лучше всего на ультраструктурном уровне изучены норадренергические нейроны. Средние диаметры пузырьков в симпатических терминалях крысы определены в 495 (малые) и 967 (крупные) Å [181]. Многие авторы указывают, однако, для крупных пузырьков цифру 750 Å. Дофаминергические нейроны мозга очень сходны с норадренергическими в ультраструктурном отношении. Слабее изучены серотонинергические элементы. В супрахиазмальном ядре гипоталамуса крысы, богатом терминалями серотонинергических волокон, найдены, помимо окончаний указанного типа, другие окончания. В них тоже имеются мелкие и крупные пузырьки, причём диаметр крупных около 1400 Å, т. е. почти вдвое больше, чем в катехоламиновых окончаниях [303]. Медиаторной функции катехоламинов и индоловых аминов посвящено много современных обзорных и обобщающих работ [например, 2, 16, 91, 98, 117, 186]. В отношении млекопитающих с уверенностью можно говорить о двух катехоламиновых медиаторах (норадреналин и дофамин) и одном индоловом (серотонин). Данных в пользу участия адреналина в синаптической передаче не было до последнего времени, когда свидетельства в пользу медиаторной функции адреналина были получены для двух групп нейронов мозга крысы [183]. Начнём с нейронов, секретирующих дофамин, — иначе говоря, с дофаминергических клеток (рис. 1). Они у млекопитающих лежат почти исключительно в среднем мозге. Одна большая группа расположена в substantia nigra и частично в вентролатеральной части ретикулярной формации среднего мозга; другая, самая мощная, занимает медиальный отдел среднего мозга. Волокна от этих двух групп следуют в краниальном направлении и заканчиваются частично в nucleus caudatus и putamen, частично в tuberculum olfactorium и nucleus accumbeus. Это — нигро-неостриарная система дофаминергических клеток. Она хорошо изучена в количественном отношении. Подсчитано, что в substantia nigra крысы насчитывается около 3500 нейронов, содержащих дофамин; каждый из них образует сеть терминальных волокон общей протяженностью 55-77 см, на этом протяжении имеется около 500 000 варикозных расширений (т. е. пресинаптических участков, из которых секретируется медиатор). В neostriatum крысы, где расположено около 4 000 000 клеток, не содержащих дофамина, этот медиатор находится в примерно 1 000 000 000 пресинаптических варикозных расширений дофаминергических аксонов, занимающих около 0,3% объема этой части ЦНС. Концентрация дофамина в пресинаптическом варикозном расширении составляет до 3000 мкг/г, тогда как в теле такого нейрона содержание амина в 30-150 раз ниже. Кроме нигро-неостриарной системы, дофаминергические нейроны имеются в составе сетчатки (их волокна не выходят за её пределы) и в гипоталамической области (в частности, клетки арочного ядра, отсылающие аксоны к срединному возвышению и в гипофиз). В периферической нервной системе дофаминв высокой концентрации обнаруживается в особых мелких клетках симпатических ганглиев [94], однако мнения о природе этих клеток расходятся: одни исследователи считают их особыми дофаминергическими интернейронами [233], другие полагают, что это островки железистой хромаффинной ткани, которая всегда богата катехоламинами. Нейроны, содержащие норадреналин, имеются в ЦНС в составе среднего мозга, варолиева моста, продолговатого и промежуточного мозга. От них восходят волокна, оканчивающиеся в гипоталамусе, таламусе, лимбических отделах переднего мозга, в коре мозга и мозжечка, и нисходят волокна в спинной мозг. Наконец, норадренергическими являются многие нейроны, расположенные вне ЦНС, в частности нейроны симпатической цепочки и некоторых периферических узлов.
 Рис. 1. Моноаминергические системы нейронов в ЦНС млекопитающих.
5-ОТ — серотонин; НА — норадреналин; ДА — дофамин. А — схема связей между отделами ЦНС, осуществляемых моноаминергическими нейронами: 1 — новая кора; 2 — лимбические отделы переднего мозга; 3 — неостриатум; 4 — палеостриатум; 5 — таламус; 6 — гипоталамус; 7 — медиальный пучок переднего мозга; 8 — средний мозг; 9 — варолиев мост; 10 — продолговатый мозг; 11 — спинной мозг; 12 — сетчатка [из 77]. Б — топография моноаминергических нейронов в мозге крысы (сагиттальный разрез, схема). Индексами указано принятое шведскими авторами обозначение групп моноаминергических клеток [из 161а].
Рис. 1. Моноаминергические системы нейронов в ЦНС млекопитающих.
5-ОТ — серотонин; НА — норадреналин; ДА — дофамин. А — схема связей между отделами ЦНС, осуществляемых моноаминергическими нейронами: 1 — новая кора; 2 — лимбические отделы переднего мозга; 3 — неостриатум; 4 — палеостриатум; 5 — таламус; 6 — гипоталамус; 7 — медиальный пучок переднего мозга; 8 — средний мозг; 9 — варолиев мост; 10 — продолговатый мозг; 11 — спинной мозг; 12 — сетчатка [из 77]. Б — топография моноаминергических нейронов в мозге крысы (сагиттальный разрез, схема). Индексами указано принятое шведскими авторами обозначение групп моноаминергических клеток [из 161а].
В продолговатом мозге крупное скопление норадренергических нейронов расположено в вентролатеральном отделе ретикулярной формации, другая небольшая группа лежит в комиссуральном ядре. В варолиевом мосте самая значительная группа находится в locus caeruleus, кроме неё имеются ещё две небольшие группы. В среднем мозге встречаются лишь отдельные клетки этого химического типа. Восходящие пучки норадренергических волокон имеют очень широкие области иннервации. Густотой терминальной сети выделяются лимбические отделы мозга; очень плотные скопления окончаний этого типа найдены в нижних отделах ствола мозга, в чувствительных и двигательных ядрах черепномозговых нервов, в ретикулярной субстанции и griseum centralis среднего мозга. Нисходящие пути прослежены от варолиева моста и продолговатого мозга до крестцового отдела спинного мозга. Один пучок проходит в боковых канатиках и даёт окончания в боковых и задних рогах; другой спускается на границе передних и боковых канатиков, заканчиваясь в передних рогах на альфа-мотонейронах, а также на преганглионарных симпатических нейронах. В спинном мозге проходит ещё один нисходящий путь норадренергических волокон, который не заканчивается в нём, а выходит в составе передних корешков. Клеточных тел, содержащих норадреналин или другие биогенные амины, в спинном мозге млекопитающих нет, но такие нейроны найдены в спинном мозге рыб [88]. Нейроны, содержащие норадреналин, наряду с дофаминергическими, имеются в составе гипоталамо-гипофизарной системы: окончания в срединном возвышении и в промежуточной доле гипофиза образованы волокнами, приходящими из медиобазальных отделов гипоталамуса. Почти все серотонинергические нейроны расположены в срединной области мозгового ствола. Из нейронов этого типа состоят, в частности, дорзальное и медиальное ядра шва продолговатого, среднего мозга и варолиева моста. Кроме того, небольшие скопления таких клеток найдены в каудальной части среднего мозга. Серотонинергические нейроны иннервируют обширные области ЦНС, включающие новую кору, гиппокамп, бледное тело, миндалину, подбугровую область, ствол головного мозга, спинной мозг. В коре наиболее плотно серотонинергические окончания представлены в её фронтальных отделах. В стволе мозга такие окончания имеются в ряде ядер черепномозговых нервов, в частности, в моторных ядрах блуждающего и тройничного нервов. Множество терминалей этого типа расположено в поверхностной части subst. grisea periventricularis, в area pretectalis и вблизи nuc. interpeduncularis. В спинном мозге серотонинергические окончания распределены примерно так же, как норадренергические.
2. 3. 3. Нейроны, имеющие медиаторами аминокислоты
Рассмотренные только что медиаторы, объединяемые названием «биогенные моноамины», — катехоламины и серотонин представляют собой продукты декарбоксилирования замещённых производных двух аминокислот, фенилаланина и триптофана. В других нейронах сами аминокислоты, по всей вероятности, берут на себя функцию медиаторов. В список таких аминокислот обычно включают гамма-аминомасляную, глутаминовую и глицин [90, 138, 139, 339]. В качестве возможных кандидатов в медиаторы называют также аспарагиновую кислоту, таурин и некоторые другие аминокислоты, но мы здесь ограничимся рассмотрением первых трёх веществ. Нужно признать, что наиболее полные доказательства медиаторной роли гамма-аминомасляной кислоты и глутамата имеются для нейронов беспозвоночных [см. обзоры — 61, 270, 279, 320, а также 237]. Напротив, предположение о медиаторной роли глицина возникло и затем получило подтверждения в процессе изучения синаптической передачи у позвоночных [см. историю вопроса в 339], а у беспозвочных пока неизвестны синапсы, где медиатором был бы глицин. Вывод о том, что глицин является передатчиком в тормозных окончаниях на спинальных мотонейронах, как будто мало кем оспаривается, так как он подтвержден убедительными экспериментальными данными. При аппликации на мотонейроны глицин полностью воспроизводит эффекты, вызываемые естественным медиатором тормозных окончаний. Действие этого медиатора, как и действие глицина, избирательно блокируется стрихнином. Наконец, распределение глицина в спинном мозге проявляет сходство с распределением интернейронов и, возможно, проприоспинальных волокон [см. 71, 99, 234, 339, а также библиографию к этим работам]. Робертс и Митчелл показали, что если изолированный препарат спинного мозга амфибий проинкубировать с меченым глицином, то можно наблюдать выход глицина в раствор в ответ на раздражение ростральной части препарата. Из ряда испытанных веществ способность высвобождаться при раздражении проявили, кроме глицина, только гамма-аминомасляная, глутаминовая и аспарагиновая кислоты, причём во всех этих случаях высвобождение происходило только в присутствии ионов кальция, т. е. носило характер секреции. Авторы ссылаются ещё на два наблюдения секреции глицина из нервных окончаний спинного мозга, одно из которых сделано на млекопитающих [274]. Добавим, что ткань спинного, а также продолговатого мозга обладает специфичной системой накопления глицина, т. е. извлечения его из внеклеточной среды и переноса через клеточную мембрану против градиента концентрации [234]. Сейчас общепризнанно, что такие системы накопления, или обратного захвата, во многих синапсах служат для освобождения синаптической щели от подействовавшей порции передатчика и для реутилизации его пресинаптическим аксоном [194]. На клетки продолговатого мозга глицин оказывает такое же действие, как на спинальные мотонейроны [190, 312]. Данные в пользу синаптической функции глицина в других отделах головного мозга менее определенны [см., однако, 99] Напротив, для головного мозга имеется хорошая система доказательств медиаторной роли гамма-аминомасляной кислоты в различных тормозных синапсах. Среди нейронов, для которых с разной степенью убедительности показана секреция гаммааминомасляной кислоты, можно назвать ряд мозжечковых нейронов (корзинчатые, звёздчатые клетки, клетки Пуркинье), зернистые клетки обонятельных луковиц, корзинчатые нейроны гиппокампа. Насколько известно, все названные клетки образуют только тормозные окончания на постсинаптических структурах [см., однако, 173]. Вывод о том, что синаптические эффекты этих клеток медиируются гамма-аминомасляной кислотой основан на следующих наблюдениях. Во-первых, апплицируемая гамма-аминомасляная кислота воспроизводит эффекты тормозных нервных окончаний. Во-вторых, в обоих случаях, т. е. при действии как естественного передатчика, так и гамма-аминомасляной кислоты, постсинаптические эффекты блокируются одними и теми же алкалоидами — пикротоксином и бикукуллином. В-третьих, гамма-аминомасляная кислота концентрируется именно в тех участках нервной ткани, где имеются соответствующие тормозные окончания. Так же локализован фермент, производящий эту кислоту, — декарбоксилаза глутаминовой кислоты, причём активность этого фермента резко падает после разрушения аксонов, дающих тормозные окончания. В-четвёртых, в области таких тормозных окончаний локализована система избирательного и противоградиентного накопления гамма-аминомасляной кислоты. В-пятых, при синаптическом торможении отмечается выход гамма-аминомасляной кислоты в экстраклеточную среду. Эти результаты, убедительно подтверждающие медиаторную функцию гамма-аминомасляной кислоты, рассматриваются в ряде работ [см. 61, 139, 159, 245, 250, 291, 339 и библиографию к ним]. Значительно менее ясным пока остаётся вопрос об участии этого медиатора в синаптической передаче в спинном мозге и в периферической нервной системе. Локализовать нейроны и синапсы этого химического типа будет гораздо легче, когда будет доработан и усовершенствован гистохимический метод выявления гамма-аминомасляной кислоты на срезах ткани [348]. В отличие от глицина и гамма-аминомасляной кислоты, за которыми закрепилось название «тормозные медиаторы», глутамат оказывает на нейроны млекопитающих, как правило, деполяризующее действие, вследствие чего его часто называют «возбуждающим медиатором». Хотя возбуждающие эффекты глутамата в самом деле отмечаются у нейронов, расположенных в самых разных отделах нервной системы млекопитающих, встречаются также клетки, в частности в коре головного мозга, которые глутаматом тормозятся [см. 43]. На пути доказательства медиаторной функции глутамата встретились серьёзные трудности. Так, действие глутамата оказалось мало специфичным и характерным для дикарбоновых аминокислот. Далее, отсутствие вещества-антагониста не давало возможности проверить, параллельно ли угнетаются эффекты глутамата и натурального передатчика возбуждающих окончаний. Эти и другие трудности, подробно рассмотренные МакЛеннаном в последнем издании его книги «Синаптическая передача», привели этого автора к заключению, что «вопрос о роли глутамата как возбуждающего передатчика в ЦНС млекопитающих должен оставаться открытым» [242, стр. 121 — 122]. Вскоре, однако, сам же Мак-Леннан с сотрудниками нашли среди производных глутаминовой кислоты два вещества с искомыми свойствами фармакологических антагонистов глутамата — альфа-метилглутамат и диэтиловый эфир глутаминовой кислоты [174, 174а]. Также и другие факты, которые накапливаются в последнее время, укрепляют уверенность в том, что глутамат служит медиатором у млекопитающих. Глутамат высвобождается из мозга, причём имеется чёткая зависимость между скоростью его выхода и уровнем активности нейронов [197]. Далее, в нервной ткани имеется высокоэффективная система избирательного захвата глутамата, подобная другим таким системам инактивации и реутилизации медиаторов. Выраженность эффектов глутамата на нервные клетки и богатство нервной ткани этой аминокислотой открывают очевидную и заманчивую перспективу установить природу синаптической передачи в огромной массе возбуждающих синапсов мозга, в частности, его коры. Мнение о том, что глутамат выполняет медиаторную функцию в ЦНС млекопитающих, имеет приверженцев среди видных специалистов [197, 199, 223, 339]. На ультраструктурном уровне аминокислотные медиаторы изучены не очень хорошо. Учизоно нашел, что в мозжечке кошки пузырьки в тормозных окончаниях уплощены, т. е. имеют на срезе форму овала, тогда как в возбуждающих окончаниях они округлые [318]. Последующие наблюдения других авторов, выполненные не только на позвоночных, подтвердили, что в секреторных пузырьках уплощённой формы хранится гамма-аминомасляная кислота. Возможно, сходное строение имеют секреторные пузырьки в глицинергических окончаниях; во всяком случае, экзогенный меченый глицин аккумулируется в спинном мозге пузырьками такой формы [234]. Напротив, глутамат хранится в округлых пузырьках. Все названные секреторные органеллы не содержат электронно-плотного материала. Для членистоногих, у которых легко идентифицировать возбуждающие и тормозные окончания на соматических мышцах, определены следующие диаметры пузырьков: для возбуждающих (содержащих глутамат) — 448±20Å, для тормозных (содержащих гамма-аминомасляную кислоту) — 460±24 (большой диаметр) и 362±18 Å (малый диаметр) [294].
2. 3. 4. Пептидергические нейроны
В 1967 г. Баргманн и соавторы при электронно-микроскопическом исследовании промежуточной доли гипофиза кошки нашли на железистых клетках пресинаптические окончания, образованные аксонами нейросекреторных клеток гипоталамуса [85]. Авторы резонно рассудили, что это наблюдение не позволяет с прежней уверенностью называть во всех случаях такие клетки нейросекреторными и в качестве альтернативы предложили новый термин — пептидергические нейроны. В их оригинальной трактовке этот термин относился только к гипоталамическим нейронам, вырабатывающим октапептиды, и, соответственно, пептидергическими синапсами назывались контакты терминалей этих клеток с клетками эндокринных органов. В последующие годы, однако, новый термин наполнился более широким содержанием. Практика пользования им в литературе показывает, что пептидергическими называют нейроны не только позвоночных, но и беспозвоночных, и что характеру структур, с которыми контактируют аксонные терминали, не придают значения. В принципе сознаётся, что важно было бы в каждом случае располагать данными о действительно пептидной природе активного агента (который вовсе не обязательно должен быть октапептидом), но в реальной действительности природа секретируемого вещества часто остается неизвестной и пептидергическими нейроны называют, опираясь на то, что ультраструктурные или гистохимические характеристики секреции в них такие же, как в нейронах с установленной пептидной природой секрета. Из сказанного видно, что понятие «пептидергические нейроны» должно отличаться от более старого понятия «нейросекреторные клетки» не только по содержанию, но и по объему. Хотя нейросекреция, по-видимому, чаще всего осуществляется пептидергическими нейронами, известны и исключения. Так, у млекопитающих туберо-инфундибулярная система катехоламиновых нейронов, вероятно, функционирует как нейросекреторная: дофамин, выделяемый окончаниями этих нейронов в срединном возвышении, как полагают, попадает в портальную систему сосудов, чтобы таким путем принять участие в регуляции секреции гонадотропина клетками гипофиза. Наличие нейросекреции непептидной природы предполагают и у насекомых. С другой стороны, пептидергические клетки во многих случаях функционируют как «обычные», а не нейросекреторные, нейроны, выделяя свои активные агенты непосредственно вблизи соответствующей клеточной мишени. Тому известно немало примеров, кроме приведённого в начале этого раздела. Так, для рыб известна прямая иннервация железистых клеток аденогипофиза гипоталамическими пептидергическими нейронами [220, 221]. У насекомых имеется целый ряд эффекторных органов, получающих прямую иннервацию пептидергического типа [163]. Иногда в этих случаях пресинаптические окончания называют «нейросекретомоторными», но упоминание о нейросекреции, которой на самом деле здесь нет, без нужды запутывает дело. Физиология синаптической передачи в пептидергических окончаниях совершенно не исследована, и эксперименты в этом направлении представили бы немалый интерес, учитывая вероятную устойчивость пептидной медиаторной молекулы. В ЦНС млекопитающих пептидергические нервные клетки сосредоточены в гипоталамической области головного мозга. Два парных клеточных ядра гипоталамуса — супраоптическое и паравентрикулярное — дают начало волокнам, образующим гипоталамо-гипофизарный тракт, который следует в заднюю долю гипофиза. Это — детально исследованная нейросекреторная система, в которой задняя доля гипофиза выступает в роли гемального органа, т. е. депо секрета, где наполненные им аксонные терминали образуют функциональный комплекс с собирающими секрет сосудами. Окончания нервных волокон содержат здесь два нейрогормона — вазопрессин (антидиуретический гормон) и окситоцин, которые при секреции попадают в общую систему циркуляции. Строение того и другого гормона известно, каждый из них представляет собой октапептид с молекулярным весом около 1000, построенный из девяти аминокислот. Гипоталамо-гипофизарной нейросекреторной системе посвящено немало сводок и монографий [см., например, 48, 143]. Клетки паравентрикулярного и супраоптического ядер обладают всеми морфологическими и физиологическими чертами нейронов. Об этом приходится сказать в связи с продолжающимися, несмотря на их нелогичность, попытками противопоставить пептидергические нейроны всем прочим типам, рассматриваемым вкупе. У пептидергических нейронов имеются дендриты и аксон, в их цитоплазме наблюдаются характерные для нервных клеток структуры. Физиология этих клеток типична для нейронов: они генерируют потенциалы действия, распространяющиеся по аксону; в аксонном окончании такой потенциал вызывает акт секреции, при этом действует типичный для нейронов механизм экзоцитоза, посредством которого секретируются и медиаторы; сами пептидергические нейроны получают аффрентацию от других клеток мозга [145, 146, 246, 251 и др.]. Идентификация нейронов супраоптического и паравентрикулярного ядер облегчается их сродством к некоторым, так называемым «нейросекреторным», красителям. Чаще всего в качестве таковых применяют хромовый гематоксилин по Баргману, паральдегид-фуксин по Габе и псевдоизоцианин по Стербе. Все три красителя, по-видимому, реализуют одну и ту же особенность химической организации этих клеток — наличие особого белка, богатого цистином. Предполагают, что окрашиваемым компонентом клеток является полипептид нейрофизин, который выделяется при секреторном акте вместе с активным началом [254], но сам активностью не обладает. Нейрофизин считают белком-носителем физиологически активного пептида. В самом деле, он обладает способностью абсорбировать вазопрессин и окситоцин, образуя комплексы с ними [см. литературу в 143]. Обладая в некоторых случаях антигенными свойствами, нейрофизин, вернее, группа нейрофизинов даёт возможность локализовать пептидергические нейроны с помощью иммунолюминесценции [см. 199а]. На ультраструктурном уровне клетки, о которых идет речь, отличаются от других типов нейронов наличием крупных, в среднем около 1900 Å, электронноплотных секреторных гранул, окруженных мембраной [201]. Органеллы этого рода нередко называют элементарными нейросекреторными гранулами, но лучше этим термином не пользоваться, так как те же гранулы могут применяться и для синаптической функции, о чём говорилось выше. Гранулы прослеживаются от тела клетки до аксонных терминалей, где они особенно многочисленны. В процессе секреции в области секреторной мембраны аксона накапливаются мелкие пузырьки, лишенные плотного содержимого, которые, по всей вероятности, представляют собой остатки секреторных гранул, изливших свое содержимое во внеклеточную среду, или продукт пиноцитоза [323, 296]. Имеются серьёзные основания считать, что вазопрессин и окситоцин вырабатываются разными группами нейронов. Содержащие эти нейрогормоны гранулы можно разделить дифференциальным центрифугированием, хотя они не различаются внешне по диаметру и электронной плотности [201]. Локальным электрическим раздражением удается вызвать преимущественную секрецию одного из двух веществ. Итогом разных экспериментов по раздельному разрушению гипоталамических ядер, по анализу содержания нейрогормонов в них и т. д. [см. ссылки в 201] является вывод, что в супраоптическом ядре находятся клетки, вырабатывающие вазопрессин, а в паравентрикуляр ном — окситоцин. Этими двумя ядрами не ограничивается система пептидергических нейронов гипоталамуса. В неё следует включить также группу мелкоклеточных ядер вентральной области (инфундибулярное, передние перивентрикулярные и некоторые другие). Считается, что в состав этих ядер входят клетки, вырабатывающие нейрогормоны, которые регулируют секреторную активность тропных желез аденогипофиза. Указанные клетки вентрального гипоталамуса связаны с аденогипофизом специальной портальной системой кровеносных сосудов. Не будучи ещё общепринятым [48], такое представление о механизме регуляторных влияний гипоталамуса на аденогипофиз получило широкое распространение [60, 143]. Нейроны мелкоклеточных ядер вентрального гипоталамуса не обладают тинкториальными свойствами пептидергических клеток супраоптического и паравентрикулярного ядер, поэтому обычные «нейросекреторные красители» при их исследовании неприменимы. Это затрудняет локализацию клеток, синтезирующих физиологически активные агенты вентрального гипоталамуса. Различают семь эффектов, оказываемых этими агентами на железистые клетки аденогипофиза. Соответственно, можно говорить о семи гипоталамических факторах, имеющих следующие обозначения: CRF — фактор, вызывающий выделение кортикотропного гормона, LRF — фактор, вызывающий выделение лютеинизирующего гормона, FSHRF — фактор, вызывающий выделение фолликулостимулирующего гормона, TRF — фактор, вызывающий выделение тиреотропного гормона, GRF или SRF — фактор, вызывающий выделение соматотропного гормона, PIF — фактор, тормозящий выделение пролактина, MIF — фактор, тормозящий выделение меланоцит-стимулирующего гормона. Пептидная природа этих факторов предполагалась давно, но только в недавнее время эти предположения стали получать прямое подтверждение. TRF идентифицирован как (пиро)глутамил-гистидил-пролинамид, т. е. как пептид, построенный всего из трёх аминокислот [101, 102б]. LRF оказался идентичным FSHRF, это — пептид, в составе которого девять аминокислот [102а, 238а]; он обладает антигенными свойствами, что позволило использовать иммунолюминесцентный метод для локализации соответствующих нейронов в мозге млекопитающих [87а]. Имеются данные, что и CRF представляет собой пептид молекулярным весом около 2500 [см. 143].
2. 3. 5. Пуринергические нейроны
Термин «пуринергические» был предложен в 1971 г. Бурнстоком для обозначения клеток, которые, как было показано этим автором и его сотрудниками, применяют в качестве медиатора АТФ или его производное [105, 106]. При изучении нервных влияний на мышцы желудочно-кишечного тракта накапливалось всё больше свидетельств тому, что, помимо адренергических тормозных и холинергических возбуждающих нервных элементов, в регуляции моторики желудочно-кишечного тракта принимают участие нервные клетки какой-то иной химической природы. Бурнстоком и сотрудниками был использован широкий арсенал методов для выяснения природы медиатора, осуществляющего неадренергические тормозные эффекты. Удалось с необходимой строгостью удовлетворить как критерий накопляемости (АТФ и продукты его гидролиза появлялись в перфузате при раздражении соответствующих нервов), так и критерий идентичности действия (АТФ и АДФ являются наиболее мощными ингибиторами моторики кишечной мышцы из большого ряда испытанных пуриновых и пиримидиновых соединений). В той же лаборатории было показано, что нервные окончания, о которых идет речь, отличаются от адренергических и холинергических окончаний, присутствующих в кишечной мышце, по ультраструктурным характеристикам: секреторные органеллы пуринергических окончаний представляют собой достаточно крупные гранулы (диаметр 1000-2000 Å), умеренно плотное содержимое которых заполняет всю гранулу, вплоть до её наружной мембраны. Фармакологические воздействия, которые резко сказывались на внешнем виде гранул адренергических окончаний, не влияли на пуринергические окончания [108]. По данным лаборатории Бурнстока, пуринергические нервные элементы представлены в разных классах позвоночных, в частности, у амфибий, птиц, млекопитающих, и принимают участие в иннервации мышц не только желудка и кишечника, но и дериватов кишечной трубки — легкого, мочевого пузыря. Не только тормозные, но и некоторые формы возбуждающих нервных влияний на эти мышцы тоже, по-видимому, осуществляются пуринергическими нейронами [107, 297].
2. 4. Заключение
Несмотря на работу большого числа исследователей, выясняющих химическую специфичность различных нервных клеток у млекопитающих животных, полученная к настоящему времени картина далека от полноты. Имеется ещё много нервных клеток, которые скорее всего не относятся ни к одному из перечисленных выше типов. Неясна медиаторная специфичность системы волокон, содержащих особые крупные (750-1600 Å) везикулы, — такие волокна представлены в дорзальном сером веществе спинного мозга [147]. Почти неизвестны синаптические механизмы, действующие в сетчатке: за исключением незначительной группы дофаминергических клеток, сложная нейронная популяция сетчатки остаётся лишь приблизительно охарактеризованной в химическом плане. Если взять такой хорошо изученный нервный центр, как мозжечок, то и здесь обнаруживаются зияющие пустоты: неизвестны медиаторы возбуждающих входов, нет данных о специфическом химизме зернистых клеток, не до конца ясно, все ли тормозные нейроны используют в качестве медиатора гамма-аминомасляную кислоту. Такие белые пятна остаются во всех отделах нервной системы. В некоторых, пока немногих, случаях кандидатами в медиаторы предложены определённые вещества, и эти предложения экспериментально проверяются. Одним из старейших кандидатов является гистамин. Некоторое время назад казалось, что эта кандидатура окончательно отпала, так как чувствительный гистохимический метод обнаружения гистамина дал возможность убедиться, что гистамин не содержится в какой-либо категории нервных элементов и локализуется только в ненервных клетках. Однако в последнее время появляются данные, говорящие в пользу кандидатуры гистамина [например, 311]. Гораздо больше интереса вызывает «вещество Р» — группа близких соединений пептидной природы. Удалось изолировать несколько форм «вещества Р» и установить строение одной из них, выделенной из гипоталамуса крупного рогатого скота,- это пептид, построенный из 11 аминокислот. «Вещество Р» обладает сильным деполяризующим действием на корковые нейроны — клетки Беца [269] и на спинальные мотонейроны [222а]. Этот факт интересен не только как свидетельство в пользу старых предположений о медиаторной роли «вещества Р» в первичных сензорных нейронах позвоночных [см., например, 242], но и как доказательство того, что крупная пептидная молекула способна выполнять роль синаптического передатчика. Предложенные кандидаты в медиаторы, однако, слишком немногочисленны, чтобы залатать бреши в знании механизмов передачи. Тем не менее уже сейчас ясно, что таких механизмов много. Мы насчитали около десятка типов нервных клеток, продуцирующих разные медиаторы (ацетилхолин, четыре биогенных амина, три аминокислоты, АТФ или его дериват), ещё столько же типов нейронов занято продукцией гипоталамических нейрогормонов. Этого достаточно, чтобы задаться вопросом о природе и причине клеточного разнообразия. Задача состоит в том, как сформулировать этот вопрос.
3. ПОСТАНОВКА ВОПРОСА О ПРИРОДЕ МЕДИАТОРНОИ СПЕЦИФИЧНОСТИ
3. 1. Аналогия или гомология?
Если медиаторное разнообразие нейронов млекопитающих имеет какую-то причину, то эта причина должна быть действительной не только для млекопитающих, но и для других организмов, имеющих нервную систему. Вряд ли могут быть сомнения в том, что составом нейронных популяций у всех животных управляют одни и те же законы. В самом деле, при сравнении организмов, относящихся к разным зоологическим типам, бросается в глаза, что их нейронные популяции не просто разнородны, но разнородны закономерно: у очень разных животных наборы медиаторов включают одни и те же вещества. Так, у ракообразных и насекомых в списке установленных или предполагаемых медиаторов мы находим глутамат, гамма-аминомасляную кислоту, дофамин, серотонин, ацетилхолин, пептиды. Почти все названные вещества, вероятно, выполняют медиаторную функцию в нервной системе моллюсков. Короче говоря, несомненно, что нейроны, относящиеся к одному и тому же типу медиаторной специфичности, могут встречаться в таких неродственных нервных образованиях, как ганглиозные тяжи турбеллярий, цепочечная нервная система аннелид и артропод, окологлоточное ожерелье гастропод и т. д. [см. обзоры 132, 153-155, 166, 209, 212, 247, 270, 279, 334]. Теорию, объясняющую разнородность клеточного состава нервной системы млекопитающих, нельзя было бы признать удовлетворительной, если бы она оказалась неспособной объяснить эту универсальную распространённость нейронов, проявляющих одинаковую медиаторную специфичность. Несомненно, что теория должна отвечать сразу на два вопроса: во-первых, почему разные медиаторы сосуществуют в одной нервной системе, и, во-вторых, почему одинаковые медиаторы представлены в разных, в том числе негомологичных нервных органах. Поставив вопрос в такой форме, мы тем самым сразу же ограничим количество мыслимых ответов. Эволюционная биология знает немного теоретически возможных причин сходства биологических структур. Такую экзотическую причину, как мимикрия, можно сразу отбросить. Вряд ли заслуживает внимания и другая нетривиальная причина — случайное совпадение: в нашем случае совпадений слишком много, чтобы считать их случайными. Остаётся классическая дилемма — аналогия или гомология. Напомним, что гомологичными называются структуры, связанные общностью происхождения, т. е. восходящие к одной и той же предковой структуре. Как видно из определения, гомологичные структуры могут и не проявлять черт сходства; сходство, однако, часто имеет место и используется при установлении гомологии структур. Подробнее с современным состоянием концепции гомологии можно ознакомиться по специальным статьям [13, 109], из которых вторая посвящена гомологиям в нервной системе; в ней же рассматриваются примеры ошибочного применения термина «гомология». Напротив, в случае аналогии сходные признаки развиваются структурами независимо (конвергентно) и адаптивно под воздействием сходных условий функционирования или общности выполняемых задач. Сходство аналогичных структур поверхностно, так как оно может появляться у совершенно неродственных образований. Но даже в случае так называемых параллелизмов, когда аналогично сходные структуры родственны, рассматриваемые черты сходства возникают не в результате родства, а независимо, под влиянием сходных условий развития.
3. 2. Гипотеза полигенеза
Предложенная мной в 1970 г. гипотеза полигенеза нейронов [53, 279], о которой уже было сказано несколько слов, реализует одну из двух альтернативных возможностей, — а именно ту, что объясняет медиаторное сходство нейронов общностью их происхождения. Я говорил выше и о том, что гипотеза имела фактические основания. Сейчас, однако, было бы удобнее не касаться этой стороны дела и представить оба возможных объяснения в априорной форме, чтобы затем сопоставить имеющиеся факты с каждым из них. Гипотеза полигенеза рассматривает множественность медиаторов как следствие гистогенетической разнородности нейронных популяций. Мы не знаем, как в действительности формировалась нейронная популяция позвоночных или членистоногих или моллюсков, но постулируем, что в этом принимали участие нейроны, относящиеся к разным клеточным линиям, причём каждая такая линия возникла независимо от других. Если разные линии нейронов существовали в диффузной нервной системе примитивных многоклеточных, то легко себе представить, что нейроны, относящиеся к одной и той же линии, могли оказаться в составе любых формаций, образовавшихся при ганглионизации. Гипотезой полигенеза постулируется также, что медиаторный химизм консервативен и что изменения химизма, которые происходили в процессе эволюции нервной системы, были изменениями в пределах исходного типа, специфического для каждой клеточной линии. По смыслу своему предлагаемая гипотеза носит генетический характер: она связывает специфический медиаторный химизм с особенным происхождением. Химическое сходство, с этой точки зрения, должны проявлять клетки, имещие общее происхождение, т. е. гомологичные; при этом не имеет значения, какую функцию они выполняют.
3. 3. Альтернатива: функциональная специализация
Гипотеза полигенеза противостоит распространённому убеждению в том, что между функцией нервного окончания или самого нейрона, с одной стороны, и типом медиаторного химизма, с другой, имеется причинная связь. Нельзя сказать, что это убеждение лишено оснований. Те, кто называют гамма-аминомасляную кислоту и глицин тормозными медиаторами, а глутамат — возбуждающим медиатором, знают, что такая терминология подкреплена фактическими данными, а ведь в ней как раз закрепляется за медиатором определённая синаптическая функция. Между тем в этих и других подобных случаях отношения причинности далеко не самоочевидны. Так, можно думать, как это делают некоторые авторы, что клетка выделяет пептидный нейрогормон, потому что она нейросекреторная; но можно рассуждать и обратным образом, а именно, что нейрон становится нейросекреторным вследствие своей пептидергичности, поскольку с таким типом химизма ему трудно обеспечить лабильную синаптическую передачу. Ниже у нас будет возможность вернуться к вопросу о характере взаимоотношений между химизмом и функцией нейрона, а пока лишь констатируем, что на этой основе можно, в принципе, представить себе конвергентное появление одного и того же типа медиаторного химизма в разных нейронах. Для этого нужно предположить, что на ранних этапах развития нервной системы все нейроны обладали одинаковым малоспециализированным химизмом и что затем, по мере углубления функциональной специализации нервных клеток, их медиаторный обмен становился всё более специализированным. Из такой схемы логически вытекает, что химическая разнородность нейронов лучше всего должна быть выражена у высших животных, тогда как у животных, стоящих на низших ступенях эволюции нервной системы, нейроны должны быть более или менее одинаковыми. В разных линиях эволюции нервной системы нейроны, сталкивающиеся со сходными функциональными задачами, должны независимо друг от друга приобретать сходный медиаторный химизм. Пожалуй, последовательнее других такое понимание природы разнообразия нейронов выразил Т. Лентц в своей книге «Примитивные нервные системы» [231]. «Я считаю, — пишет Лентц, — что нервная система примитивного предкового организма состояла из нервных клеток единственного базового типа… Примитивная нервная клетка, видимо, была способна продуцировать нейрогуморы и нейросекрет и, следовательно, могла дать начало как обычным нейронам, так и нейросекреторным клеткам. В начале оба класса веществ, видимо, действовали гормонально. Обычные нейроны приобрели специализацию в основном для образования нейрогуморов и выделения веществ в синаптических контактах на эффекторы, где эти агенты действуют как нервные передатчики. Путём специализации в основном для секреции полипептидных гормональных веществ и выведения этих веществ во внеклеточные пространства или возле сосудистых систем примитивная нервная клетка могла дать начало нейросекреторным нейронам. У высших животных случаи, когда нейрогуморы действуют на большом расстоянии и, следовательно, выполняют гуморальные функции, или когда в одной и той же клетке имеется комбинация нейрогуморов и нейросекреторного вещества, видимо, свидетельствуют, что такие клетки сохранили или обрели способность вернуться к примитивному состоянию» [231, стр. 117 - 118]. Лентц считает, что специализация нейронов в ходе эволюции выражалась в уменьшении числа медиаторов, секретируемых в аксонных окончаниях (например, о специализации вставочных и моторных нейронов он пишет: «Можно думать, что… их агенты-передатчики были ограничены до небольшого числа нейрогуморов» (стр. 117). В приведенных словах речь не идет о том, каким способом у разных животных появляются нейроны с одинаковым типом химизма, но это легко себе представить, если предположить, что схема Лентца работает параллельно в разных линиях эволюции многоклеточных животных. Вообще, явления параллелизма и, шире, аналогичного развития нервных структур хорошо известны, и вполне естественно, что этот теоретический опыт переносится некоторыми авторами на анализ медиаторного химизма. Как известно, крупнейший гистолог-эволюционист академик А. А. Заварзин проявлял особый интерес к явлениям параллельного развития нервных структур [18]. Не случайно в работах близких к Заварзину авторов развивается мысль, что сходные по своему назначению нервные клетки должны обладать сходным медиаторным химизмом. Мы имеем в виду, в частности, попытки доказать, что так называемая симпатическая нервная система первичноротых имеет адренергическую природу [324, 271]. Профессор М. Я. Михельсон, признанный авторитет в области сравнительной фармакологии, неоднократно говорил в своих работах о независимом приобретении нейронами холинергического типа специфичности у первичноротых и вторичноротых животных [39, 243]. Процитируем одну из работ: «Я, — пишет М. Я. Михельсон, — хотел бы привлечь внимание к факту, что у кишечнополостных холинергическая передача не доказана и сомнительно, что имеет место химическая передача нервных импульсов [см., однако, 38, 341 - 343! — Д. С.]. Тем не менее, природа нашла ацетилхолин в качестве одного из химических передатчиков в обеих главных ветвях филогенетического древа животных — в ветви первичноротых и в ветви вторичноротых. Я хотел бы подчеркнуть, что ацетилхолин найден на роль передатчика в филетических линиях, которые развивались независимо одна от другой в течение сотен миллионов лет. Можно думать, что молекула ацетилхолина — очень удобный кандидат на роль медиатора» и т. д. [243, стр. 129]. Число таких примеров легко расширить. Все они свидетельствуют о широкой распространенности представлений, которым противостоит предложенная нами гипотеза. Эти представления, если попытаться выразить их в логически последовательной форме, должны звучать примерно так: 1. Медиаторная специфичность появляется у нейронов в процессе их эволюции как результат специализации химизма; иными словами, все специфические типы нейронов являются продуктами дивергентной эволюции примитивного плюрихимического нейрона, способного синтезировать и секретировать смесь медиаторных веществ. 2. Специализация химизма имела место в процессе эволюции в связи с функциональной специализацией нейронов; иными словами, у нейронов, которые сталкивались со сходными функциональными задачами, независимо развивался сходный тип медиаторного химизма.
3. 4. Что важно знать для проверки гипотез
Априорно, каждая из двух рассмотренных гипотез удовлетворяет условиям задачи, объясняя и различия, и сходство. Их исходные постулаты при этомвзаимно противоположны:
| Гипотеза полигенеза | Альтернатива | |
|---|---|---|
| Един ли источник происхождения нейронов? | Нет | Да |
| Адаптивен ли тип медиаторного химизма? | Нет | Да |
| Приобретается ли нейронами свойство медиаторной специфичности в процессе эволюции нервной системы? | Нет | Да |
| В чём причина существования нейронов с одинаковым медиаторным химизмом у самых разных животных? | Общность происхождения таких нейронов (гомология) | Общность условий функционирования таких нейронов (аналогия) |
Очевидно, что два объяснения не сходятся ни в едином пункте. Это естественно, так как только в такой форме они не обременены внутренними противоречиями. Обращаясь теперь от этих априорных схем к реальным фактам, мы сознаем, что лишены прямой возможности изучить, одинаковыми или разными были нейроны в момент их возникновения в филогенезе животных, сохраняли изначальный тип химизма или приобрели специфичность в ходе эволюции. Однако каждая из двух гипотез жёстко связана с некоторыми следствиями, подлежащими экспериментальной проверке. Сравнительное изучение нервных клеток должно дать разные результаты в зависимости от того, какая из гипотез верна. Если правильна функциональная гипотеза, то должно существовать соответствие между типом медиаторного химизма и функцией нейрона или физиологическими свойствами его аксонных окончаний. Напротив, генетическая гипотеза допускает, что при одном и том же медиаторе клетки и синапсы могут обладать совершенно разными свойствами и выполнять разные функции. Далее, функциональная гипотеза предсказывает, что качественная неоднородность нейронного состава будет тем невнятнее выражена, чем проще нервная система. Если же верна генетическая гипотеза, то в простых нервных системах может оказаться так же много типов химизма, как в сложных. Наконец, если сравнивать нейроны, относящиеся к одному и тому же химическому типу, но к разным нервным системам, то и здесь ожидаются разные результаты. При правильности функциональной гипотезы, чем примитивнее нервная система, тем менее выраженной будет химическая специфичность, так что, например, холинергический нейрон должен быть отчасти нехолинергическим, зато дофаминергический должен быть «немного холинергическим», «немного пептидергическим» и т. д. Генетическая же гипотеза не ожидает от примитивных нейронов плюрихимизма и предсказывает, что степень специфичности нейронов должна быть равной на всех уровнях эволюции нервной организации.
4. НЕОДНОРОДНОСТЬ КЛЕТОЧНОГО СОСТАВА НЕРВНОЙ СИСТЕМЫ ВИНОГРАДНОЙ УЛИТКИ
4. 1. Виноградная улитка
Из сказанного в предыдущем разделе следует, что решение проблемы во многом зависит от результатов изучения простых нервных систем. Чтобы выяснить, зависит ли в действительности химическая (медиаторная) разнородность нейронной популяции от её функциональной дифференцированности, нужно располагать достаточно детальными знаниями о клеточном составе какой-нибудь нервной системы, проявляющей существенно иной уровень сложности по сравнению с нервной системой млекопитающих. Большой интерес в этом смысле представили бы организмы с наиболее примитивной нервной системой — такие, как кишечнополостные, гребневики, сколециды, иглокожие. Но нужно признать, что о клеточном составе их нервных систем известно относительно немного. Хотя эти немногие факты очень важны (см. об этом ниже, в разделах 6.4 и 6.5), они не могут удовлетворить потребность в систематических сведениях об относительно простой нейронной популяции. Гораздо лучше изучены некоторые другие беспозвоночные. Их нервная система, конечно, намного сложнее, чем у кишечнополостных, но всё же она намного проще, чем у млекопитающих. К таким «оптимальным» объектам можно отнести медицинскую пиявку, представляющую тип аннелид, некоторых ракообразных, насекомых и гастропод. Последние из названных животных располагают важным преимуществом перед другими — гигантизмом нейронов. Благодаря наличию гигантских и крупных нейронов брюхоногие оказались хорошо изученными — и не только в отношении медиаторов и нейрогормонов, но и в отношении назначения отдельных нервных клеток, а также в отношении физиологических эффектов медиаторных веществ. Поэтому результаты, полученные на гастроподах, дают возможность одновременно обсуждать другой важный вопрос: имеется ли зависимость между функцией нейрона (синапса) и типом медиаторного химизма. Гастропод, которые стали объектами систематических нейробиологических исследований, довольно немного. Это — аплизия и тритония, представляющие два разных отряда подкласса заднежаберных, затем прудовик и катушка, представители сидячеглазых пульмонат, наконец, виноградная улитка — тоже из подкласса пульмонат (лёгочных), но из отряда стебельчатоглазых.
 Рис. 2. Строение центральной нервной системы виноградной улитки.
А — позиция центральных ганглиев в головной области моллюска (на схеме изображены только правые ганглии и коннективы). Б — относительные позиции отдельных центральных ганглиев и связи между ними: 1 — церебральный ганглий; 2 — педальный ганглий; 3 — комплекс ганглиев висцеральной дуги; 4 — буккальный ганглий; 5 — церебропедальный коннектив; 6 — цереброплевральный коннектив; 7 — церебробуккальный коннектив; 8 — глотка; 9 — пищевод; 10 — ретрактор глотки; 11 — нога.
Рис. 2. Строение центральной нервной системы виноградной улитки.
А — позиция центральных ганглиев в головной области моллюска (на схеме изображены только правые ганглии и коннективы). Б — относительные позиции отдельных центральных ганглиев и связи между ними: 1 — церебральный ганглий; 2 — педальный ганглий; 3 — комплекс ганглиев висцеральной дуги; 4 — буккальный ганглий; 5 — церебропедальный коннектив; 6 — цереброплевральный коннектив; 7 — церебробуккальный коннектив; 8 — глотка; 9 — пищевод; 10 — ретрактор глотки; 11 — нога.
Несколько подробнее о системе гастропод и объектах нейробиологических исследований будет говориться в разделе 5.2., а сейчас перейдём к виноградной улитке, выбранной из этого небольшого списка в качестве «опорного» объекта, поставляющего материалы для данной главы. Этот выбор объясняется не только тем, что виноградная улитка больше, чем другие названные моллюски, известна автору. Немаловажное значение имеет и тот факт, что существует давняя традиция использования этой улитки в сравнительно-физиологических исследованиях. В частности, X. С. Коштоянц, который ещё в 30-х годах проявил интерес к физиологически активным веществам нервной системы виноградной улитки [29, см. также 30, 32], утвердил эту традицию в отечественной физиологии, восприняв её от утрехтской школы Германа Иордана. Будучи традиционным объектом, виноградная улитка изучена в нейробиологическом отношении гораздо лучше, чем формы, появившиеся в поле зрения физиологов только в связи с интересом к гигантским нейронам (в несколько меньшей степени сказанное относится и к аплизии, которую также изучали давно и много). Это касается таких вопросов, как взаимоотношения между ганглиями, нервная регуляция движений, сердцебиений и т. д. [см. основные результаты и библиографию в 32, 102 и 305]. Наряду со старыми физиологическими работами, обстоятельные исследования по микроскопической анатомии нервной системы виноградной улитки [например, 176, 226, 226а] обеспечивают современных авторов необходимыми исходными данными.
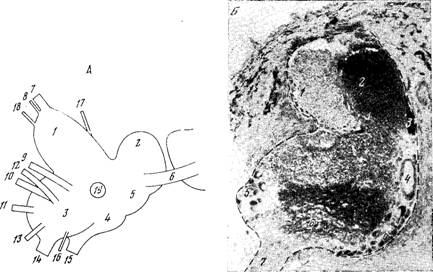 Рис. 3. Церебральные гланглии виноградной улитки.
А — общий вид правого ганглия с вентральной стороны: 1 — процеребрум; 2 — мезоцеребрум; 3 - педальная доля метацеребрума; 4 — плевральная доля метацеребрума; 5 — комиссуральная доля метацеребрума; 6 — церебральная комиссура; 7 — тентакулярный нерв; 8 — зрительный нерв; 9, 10, 11 — внутренний, средний, наружный губные нервы; 12 — нерв пениса (непарный, в отличие от остальных церебральных нервов); 13 — церебробуккальный коннектив; 14 — церебропедальный коннектив; 15 — цереброплевральный коннектив; 16 — нерв статоциста; 17 и 18 — внутренний и наружный перитентакулярные нервы; 19 — гигантская метацеребральная клетка ПЦ1. Б — вентральная область ганглия на срезе, прошедшем через процеребрум и педальную долю метацеребрума. Реакция на дегидрогеназу янтарной кислоты (высокая активность фермента в синаптическом нейропиле и цитоплазме некоторых нейронов). Граница между нервной тканью и оболочкой ганглия обозначена пунктиром: 1 — клеточная масса процеребрума (скопление мельчайших нейронов); 2 — терминальный нейропиль процеребрума; 3 — внутренний нейропиль процеребрума; 4 — гигантская метацеребральная клетка ПЦ1; 5 — латеральная клеточная кора педальной доли; 6 — нейропиль педальной доли; 7 — церебропедальный коннектив.
Рис. 3. Церебральные гланглии виноградной улитки.
А — общий вид правого ганглия с вентральной стороны: 1 — процеребрум; 2 — мезоцеребрум; 3 - педальная доля метацеребрума; 4 — плевральная доля метацеребрума; 5 — комиссуральная доля метацеребрума; 6 — церебральная комиссура; 7 — тентакулярный нерв; 8 — зрительный нерв; 9, 10, 11 — внутренний, средний, наружный губные нервы; 12 — нерв пениса (непарный, в отличие от остальных церебральных нервов); 13 — церебробуккальный коннектив; 14 — церебропедальный коннектив; 15 — цереброплевральный коннектив; 16 — нерв статоциста; 17 и 18 — внутренний и наружный перитентакулярные нервы; 19 — гигантская метацеребральная клетка ПЦ1. Б — вентральная область ганглия на срезе, прошедшем через процеребрум и педальную долю метацеребрума. Реакция на дегидрогеназу янтарной кислоты (высокая активность фермента в синаптическом нейропиле и цитоплазме некоторых нейронов). Граница между нервной тканью и оболочкой ганглия обозначена пунктиром: 1 — клеточная масса процеребрума (скопление мельчайших нейронов); 2 — терминальный нейропиль процеребрума; 3 — внутренний нейропиль процеребрума; 4 — гигантская метацеребральная клетка ПЦ1; 5 — латеральная клеточная кора педальной доли; 6 — нейропиль педальной доли; 7 — церебропедальный коннектив.
Систематическое положение виноградной улитки следующее: тип моллюски (Mollusca), класс лёгочные (Pulmonata), отряд стебельчатоглазые (Stylommatophora), семейство хелициды (Неlicidae). Хелициды — европейское, средиземноморское семейство, но некоторые его представители занесены человеком в новые районы и значительно расширили свой ареал. Так, обычная для Западной Европы садовая улитка Helix aspersa (синоним — Cryptomphallus aspersa) теперь обычна в Австралии, Южной Америке. В нейробиологической литературе уже на протяжении ряда лет данные, полученные на «аргентинской улитке» С. aspersa, рассматриваются как не имеющие отношения к данным, полученным на нейронах европейской садовой улитки Н. aspersa; это, конечно, недоразумение.
 Рис. 4. Подглоточные ганглии виноградной улитки.
А — ганглии висцеральной дуги с дорзальной (верхний рисунок) и вентральной (нижний рисунок) стороны: 1 — правый плевральный ганглий; 2 — правый париетальный ганглий; 3 — висцеральный (абдоминальный) ганглий; 4 — левый париетальный ганглий; 5 — левый плевральный ганглий; 6 — цереброплевральный коннектив; 7 — плевропедальный коннектив; 8 — наружный правый мантийный нерв; 9 — внутренний правый мантийный нерв; 10 — аортальный нерв; 11 — интестинальный нерв; 12 — анальный нерв; 13 — кожный мантийный нерв; 14 — левый мантийный нерв; 15 — нерв колумеллярной мышцы. Б — педальные ганглии с дорзальной (верхний рисунок) и вентральной (нижний рисунок) стороны: 1 — правый педальный ганглий; 2 — левый педальный ганглий; 3 — передняя педальная комиссура; 4 — задняя педальная комиссура; 5 — церебропедальный коннектив; б — плевропедальный коннектив; 7-16 — нервы ноги; 17 — статоцист. Кожные педальные нервы и нервы педальной артерии на схеме не изображены.
Рис. 4. Подглоточные ганглии виноградной улитки.
А — ганглии висцеральной дуги с дорзальной (верхний рисунок) и вентральной (нижний рисунок) стороны: 1 — правый плевральный ганглий; 2 — правый париетальный ганглий; 3 — висцеральный (абдоминальный) ганглий; 4 — левый париетальный ганглий; 5 — левый плевральный ганглий; 6 — цереброплевральный коннектив; 7 — плевропедальный коннектив; 8 — наружный правый мантийный нерв; 9 — внутренний правый мантийный нерв; 10 — аортальный нерв; 11 — интестинальный нерв; 12 — анальный нерв; 13 — кожный мантийный нерв; 14 — левый мантийный нерв; 15 — нерв колумеллярной мышцы. Б — педальные ганглии с дорзальной (верхний рисунок) и вентральной (нижний рисунок) стороны: 1 — правый педальный ганглий; 2 — левый педальный ганглий; 3 — передняя педальная комиссура; 4 — задняя педальная комиссура; 5 — церебропедальный коннектив; б — плевропедальный коннектив; 7-16 — нервы ноги; 17 — статоцист. Кожные педальные нервы и нервы педальной артерии на схеме не изображены.
Виноградной улиткой, строго говоря, называется вид Helix pomatia. Этот вид тоже несколько расселился из Западной и Центральной Европы, — так, в Советской Прибалтике, под Курском и в некоторых других местах он обитает потому, что улиток некогда завезли туда в гастрономических целях. Именно на этом виде проведены многие нейробиологические исследования европейских авторов. Что касается работ, выполненных в нашей стране, то в них виноградными улитками нередко называются улитки, собранные в Крыму или на Кавказе, где Н. pomatia не обитает. В этих случаях объектом исследования чаще всего служит Н. lucorum — вид, очень близкий к Н. pomatia. У этих двух видов сходны не только анатомические черты строения нервной системы, но даже клеточные карты, что позволяет объединить данные, полученные на той и другой форме и касающиеся клеточного состава нейронной популяции. Как и у других гастропод, у виноградной улитки значительное число нервных клеток расположено вне ганглиев, в составе эффекторных органов. Хорошо известным примером является нервное сплетение ноги. В отношении таких периферически расположенных нервных клеток часто применяют термин «нервная сеть», но это не совсем верно, так как на деле это обычно не сеть отдельных нейронов, а сеть, построенная из небольших ганглиев, в состав которых помимо клеточных тел нейронов входят участки синаптического нейропиля. У виноградной улитки такие периферические нейроны почти не изучались, объектом цитофизиологических исследований служили почти без исключения клетки ганглиев центральной нервной системы. Эти ганглии собраны у виноградной улитки в компактную массу, состоящую из двух частей, надглоточной и подглоточной. Надглоточную часть, или мозг, составляет пара церебральных ганглиев. Она связана двумя парами коннективов с подглоточным комплексом, в составе которого семь ганглиев: дорзальную часть комплекса образуют ганглии висцеральной дуги (два плевральных, два париетальных и непарный абдоминальный), а вентральную — парные педальные ганглии. Вместе надглоточные и подглоточные ганглии образуют окологлоточное кольцо, с которым тесно связаны лежащие здесь же, у основания пищевода, парные буккальные ганглии. Одиннадцать названных ганглиев дают начало многочисленным нервам, идущим к разным частям тела (рис. 2 — 4). Гигантские нейроны в том или ином числе представлены в каждом из одиннадцати ганглиев [226, 226а]. В головном отделе виноградной улитки имеются также дополнительные сенсорные ганглии, в частности, у концов каждого из четырёх щупалец.
4. 2. Результаты электрофизиологических исследований
4. 2. 1. Развитие исследований на идентифицируемых нейронах гастропод
Физиологам, начинавшим в 50-60-х гг. работу с отдельными нейронами гастропод, пришлось столкнуться с неожиданной трудностью, заключавшейся в том, что наблюдения, которые делались во время эксперимента, зачастую не соответствовали представлениям о нервной клетке, почерпнутым из физиологических руководств. В разных клетках одного ганглия регистрировались совершенно разные величины мембранного потенциала; потенциалы действия то оказывались равными мембранному потенциалу, то превышали его более чем вдвое, то были размером всего в несколько милливольт. Этого мало. Иногда наблюдатель мог видеть, как длительность потенциала действия растёт в течение залпа и снова уменьшается после паузы. В других случаях клетка генерировала потенциалы действия нескольких разных амплитуд, причем каждый из них разряжался со своим собственным ритмом. Автор этих строк хорошо помнит то не очень давнее время, когда даже авторитетные физиологи отвергали некоторые из полученных результатов, полагая их артефактами, да и сами экспериментаторы охотно делили нейроны на «хорошие» и «плохие», выбирая те, которые ведут себя согласно учебнику. Эти трудности были скорее субъективными, чем объективными. На смену нашим предвзятым представлениям о нейроне постепенно приходило понимание того, что реальность сложнее схемы. Именно в процессе исследования «плохих» клеток и отступлений от правил были добыты знания, обогатившие физиологию. Так, изучение упомянутого выше аномального поведения потенциалов действия позволило открыть новое явление — независимую активность разных аксонных ветвей одного нейрона [305, 310]. Напомним также, какую огромную литературу породило наблюдение киевских авторов, что в одном и том же ганглии нейроны различаются по своей способности генерировать потенциалы действия в безнатриевых растворах [11]. Число таких примеров можно умножить. Успешному разрешению проблемы «плохих нейронов» способствовала работа на идентифицируемых клетках. Лишь при многократном изучении одной и той же клетки в разных препаратах исследователь может установить, что та или иная аномалия не является результатом повреждения или ошибки опыта, а выражает собой особенность данного нейрона. Нам представляется, что в настоящее время, когда большинство исследователей работает на идентифицируемых нейронах и когда свойства ряда нейронов описаны достаточно полно, уже невозможно дать описание физиологии абстрактного «нейрона моллюсков», настолько различны физиологические свойства разных нейронов. Внутриклеточную регистрацию потенциалов из крупных нейронов гастропод впервые осуществили в 1955 г. во Франции А. Арванитаки и Н. Халазонитис; не без их влияния в другой французской лаборатории примерно в то же время такую же работу начал Л. Тауц. Объектом первых исследований были морские заднежаберные моллюски рода аплизия (Aplysia), однако вскоре обе лаборатории стали также публиковать результаты, полученные на нейронах виноградной улитки. Существенная разница заключалась, однако, в том, что, работая на аплизии, исследователи могли визуально идентифицировать некоторые нейроны, т. е. имели возможность изучать индивидуальные характеристики определенных клеток, тогда как на виноградной улитке работа проводилась на случайных клетках. Единственным исключением была гигантская метацеребральная клетка, выделяющаяся по своим размерам [204, 205]. Ярко и по-разному пигментированные нейроны аплизии, покрытые прозрачным периневрием, делали лёгкой идентификацию, которая казалась невозможной при работе с одинаково, на первый взгляд, слабо пигментированными нейронами виноградной улитки, покрытыми толстой и непрозрачной соединительнотканной оболочкой ганглия. Уже в первые годы работы на нейронах аплизии обе французские лаборатории пришли к выводу, что определённые, узнаваемые по внешнему виду нейроны всегда, от препарата к препарату, обладают определённым характером электрической активности. В начале 60-х годов микроэлектродные исследования на нейронах гастропод начались в нашей стране, США и Англии. В США, где эти исследования быстро приобрели широкий размах, пионером их явился Ф. Струмвассер, с которым Арванитаки и Халазонитис поделились своим опытом работы на аплизии [см. историю вопроса в 302]. Может быть, по этой причине аплизия надолго стала основным объектом американских исследователей, несмотря на наличие достаточно удобных местных пресноводных и наземных объектов. В Англии Г. Керкут и его сотрудники убедительно показали, что исследования, в том числе на идентифицируемых нейронах, можно с успехом вести на местном виде — садовой улитке, Helix (Cryptomphallus) aspersa, которая не обладает ни крупными размерами, ни ярко пигментированными нейронами. На этом же виде выполнена серия исследований в Южной Америке. В Японии несколько интересных исследований было выполнено на гигантских нейронах безраковинного лёгочного моллюска Onchidium, обитателя опреснённых мелководий; но эта работа не приобрела широкого размаха. В нашей стране исследователи также пошли по пути освоения объектов, представляющих местную фауну гастропод. Коллектив сотрудников П. Г. Костюка на протяжении ряда лет использует для микроэлектродных экспериментов нейроны катушки (Planorbarius corneus), прудовика (Lymnaea stagnalis) и виноградной улитки. Нужно заметить, что катушка и прудовик, как и другие сидячеглазые лёгочные моллюски, обладают, подобно аплизии, ярко пигментированными нервными клетками, многие из которых имеют крупные размеры и легко поддаются визуальной идентификации [например, 290]. Картирование идентифицируемых нейронов, т. е. описание свойств клеток, которые могут быть опознаны визуально или найдены в определенной области на основе этого описания, было успешно осуществлено группой Канделя на абдоминальном ганглии аплизии [160] и Уиллоусом на ЦНС тритонии [347]. Работа, проведённая мной совместно с Я. Шаланки, явилась как будто первым опытом картирования нейронов виноградной улитки [283]. Исследования в этом направлении я продолжил в сотрудничестве с Г. Н. Коробцовым. В литературе последних лет — как отечественной, так и зарубежной, уже имеется ряд исследований, выполненных на идентифицируемых клетках виноградной улитки.
4. 2. 2. Обозначение нейронов и их визуальная идентификация
Как уже было отмечено выше, первым идентифицируемым объектом при работе на нейронах виноградной улитки явилась гигантская клетка, расположенная в каждом из церебральных ганглиев на вентральной поверхности вблизи корешков губных нервов. Эта клетка резко выделяется своими размерами среди окружающих нейронов, что позволило авторам исследования, Канделю и Тауцу, легко обнаруживать её в разных препаратах и провести ценное исследование. К этим парным метацеребральным нейронам мы вернёмся позже (4.2.5). В целом же нейроны церебральных ганглиев, не отличающиеся значительными размерами, не привлекали исследователей, что отчасти, по нашему мнению, объясняется и их физиологическими свойствами: клетки церебральных ганглиев зачастую ведут себя как «плохие нейроны», т. е. генерируют псевдоспайки, а не полные потенциалы действия, мало чувствительны к поляризующему току, и т. п. Все отечественные и зарубежные лаборатории, работающие на хелицидах, словно сговорившись, избрали в качестве основного или даже единственного препарата дорзальную поверхность подглоточного комплекса ганглиев. Подглоточный комплекс составляют семь ганглиев, из которых дорзально расположены пять: 1) левый плевральный (ЛПлГ); 2) левый париетальный (ЛПаГ); 3) абдоминальный, или висцеральный (ВГ); 4) правый париетальный (ППаГ); 5) правый плевральный ганглий (ППлГ). Среди клеток, расположенных на дорзальной поверхности этих ганглиев, многие имеют крупные размеры и обладают активностью, представленной высокоамплитудными потенциалами действия. Эти качества, наряду с лёгкостью препаровки, делают такой препарат привлекательным для физиолога. Однако само большое количество крупных клеток и опредёленная изменчивость анатомической картины сильно затрудняют визуальную идентификацию индивидуальных нейронов. В своей первой работе, посвящённой картированию идентифицируемых клеток в этом препарате [283], мы пришли к выводу, что уверенную идентификацию клеток может обеспечить только сочетание визуальной оценки с физиологическим исследованием. В этой статье на карту ганглия было нанесено 10 индивидуальных клеток и 6 групп, каждую из которых составляют клетки, обладающие сходными свойствами. Для обозначения нейронов мы использовали способ, близкий к тому, который введён группой Канделя при картировании нейронов абдоминального ганглия аплизии [160]. Каждая клетка обозначена сокращённым названием ганглия и порядковым номером, например, ППа1, ППа2, и т. д.
 Рис. 5. Типичные позиции идентифицируемых индивидуальных клеток (А) и клеточных групп (Б) на дорзальной поверхности ганглиев висцеральной дуги.
Обозначения ганглиев: ППл — правый плевральный, ППа — правый париетальный, В — висцеральный, ЛПа — левый париетальный, ЛПл — левый плевральный. Объяснения в тексте.
Рис. 5. Типичные позиции идентифицируемых индивидуальных клеток (А) и клеточных групп (Б) на дорзальной поверхности ганглиев висцеральной дуги.
Обозначения ганглиев: ППл — правый плевральный, ППа — правый париетальный, В — висцеральный, ЛПа — левый париетальный, ЛПл — левый плевральный. Объяснения в тексте.
Скопления однородных нейронов обозначаются заглавными буквами латинского алфавита: А, В, С и т. д. Такая система оставляет открытой возможность дальнейшего расширения списка изученных нейронов. В самом деле, в процессе последующих исследований на карте появились новые идентифицируемые клетки [24]. Сочетая визуальную и физиологическую оценку клеток, исследователи, работающие во многих лабораториях нашей страны и за рубежом, успешно проводят эксперименты на нейронах дорзальной поверхности подглоточного комплекса ганглиев виноградной улитки [1, 10 - 12, 20, 26, 28, 45, 46, 171, 288, 289, 298а, б, 346 и др.]. Некоторые из этих авторов, применяя нашу систему обозначения клеток, ещё более расширили набор идентифицируемых нейронов, и эта работа, несомненно, будет продолжаться. Вместе с тем, мы провели предварительное обследование нейронов, расположенных на вентральной поверхности упомянутых выше пяти ганглиев, а также на дорзальной и вентральной поверхности педальных ганглиев, занимающих в подглоточном комплексе вентральное положение. Оказалось, что гигантские и крупные клетки, выделяющиеся своими размерами среди прочих нейронов и обладающие характерными физиологическими и фармакологическими свойствами, имеются не только на дорзальной стороне подглоточного комплекса. На рис. 5 показана картина клеточной коры, в препаратах подглоточного комплекса ганглиев. Эти рисунки представляют собой схему, составленную на основании многих десятков рисунков, сверенных с результатами физиологических экспериментов. Каждый конкретный препарат в той или иной степени отличается от этой схемы, поскольку позиции определённых клеток довольно изменчивы. Всё же, имея определенный навык, в большинстве случаев можно без труда найти нужную клетку; ещё легче найти нейрон, представляющий ту или иную однородную группу. При визуальной идентификации учитываются следующие три признака: размеры, цвет и позиция. Каждый из них колеблется лишь в известных пределах. Почти все клетки, обозначенные на карте номерами, имеют гигантские или, по крайней мере, крупные размеры. Этот признак помогает без затруднений идентифицировать, например, клетки ЛПл1 и ППл1, (диаметр — более 150 мк), когда они занимают типичную позицию, так как на дорзальной поверхности плевральных ганглиев больше нет гигантских и даже крупных клеток. Однако иногда позиции гигантских плевральных нейронов изменены, к чему более склонна ППл1. Тело этого нейрона может оказаться лежащим не в самом ППлГ, а в пограничной с ним области ППаГ, на что ещё в 1918 г. обратила внимание Кунце, изучавшая срезы ганглиев [226]. При нетипичном положении этой или иной клетки возрастает значение физиологической идентификации. Колебания размерных характеристик более выражены для одних нейронов (например, ВЗ, В5), чем для других (например, ЛПа2, ППа1). Цвет нейронов улитки только неопытному глазу представляется одинаковым. При внимательном рассмотрении неповрежденных нейронов легко заметить, что цитоплазма одних имеет желто-оранжевый оттенок, других — белесоватый, третьих — молочно-белый; кроме того, часть нейронов содержит коричневатые зерна пигмента. Эти признаки также довольно постоянны и могут служить характеристикой определённых нейронов. Так, нейроны группы F, расположенные более или менее компактно в ВГ, выделяются прозрачной оранжеватой цитоплазмой; они, как и хорошо пигментированные нейроны группы Е, никогда не содержат белого материала. В отличие от этого, клетки группы D иногда бывают совершенно белыми, но и в тех случаях, когда в них мало белого секрета, они выделяются отсутствием пигментации, бледным видом цитоплазмы. Присутствие белого материала характерно также для клеток, относящихся к группам А, В и С. Индивидуальные, отдельно от групп лежащие белые нейроны, имеются в обоих париетальных и висцеральном ганглиях. Клетка ППа1 иногда бывает белесоватой. Об изменчивости позиций уже говорилось выше. Отметим клетку ЛПа1, которая в некоторых случаях лежит не в ЛПаГ, а рядом с ним, в ВГ, т. е. по другую сторону от складки, разделяющей два ганглия. Большинство нейронов, однако, имеет достаточно постоянное расположение. Детальнее см. об этом в упомянутой статье Кунце [226].
4. 2. 3. Электрофизиологические критерии идентификации
Даже с помощью одного внутриклеточного электрода можно получить довольно разнообразную информацию о клетке. Важно из всех этих возможностей выбрать такие, которые позволяют, взятые в совокупности, с уверенностью находить нужную клетку. Рассмотрим следующие показатели: спонтанная импульсная активность; мембранный потенциал; потенциалы действия; спонтанный синаптический приток; реакция клетки на поляризацию мембраны; биофизические характеристики клеточной мембраны; фармакологические характеристики клеточной мембраны. Спонтанная активность. В своей первой работе, посвящённой идентификации нейронов улитки, мы исключили этот показатель как сильно изменчивый и потому ненадёжный [283]. Главной причиной такого решения послужили в то время наблюдения, проведённые на многих препаратах на клетке ППа1. Эта клетка, проявляющая уникальный тип залповой активности, у некоторых улиток оказывалась молчащей, у других имела нерегулярную незалповую активность, и нам приходилось прибегать к фармакологическим критериям, чтобы убедиться в том, что исследуемая клетка — действительно ППа1. Ныне, обладая несколько большим опытом, мы вслед за многими авторами, работающими на нейронах гастропод, считаем, что характер спонтанной активности вправе рассматриваться среди свойств, постоянных для каждого нейрона. Наш опыт, как и опыт других исследователей, показывает, что определённые клетки относятся к типу «молчащих», т. е. не проявляют спонтанной активности (например, ЛПа2 и ППа3); другие имеют активность регулярного типа, т. е. с довольно постоянными интервалами между потенциалами действия (например, группа F); третьи проявляют нерегулярную активность — относительно редкую (например, ЛПл1, ППл1) или частую (ППа4); четвёртые склонны к залповым разрядам, которые в разных клетках неодинаковы. Как оказалось, упомянутая выше изменчивость спонтанной активности, особенно сильно выраженная у некоторых нейронов, зависит от некоторых факторов, знание которых помогает прояснить картину. Исследования, проведённые на изолированных идентифицированных нейронах гастропод, подтвердили, что клеткам в самом деле присущи определённые свойства — например, наличие или отсутствие пейсмекерных свойств, способность разряжаться характерными залпами и т. д. [118].Реализация этих свойств в целостном многоклеточном препарате сильно зависит, во-первых, от синаптического притока, который может меняться при незначительных изменениях условий препаровки, при повреждениях окружающих нейронов и проводящих путей и т. д. Имея это в виду, целесообразно, может быть, оценивать характер спонтанной активности дважды: в обычных условиях и в условиях блокирования синаптического притока. По нашим наблюдениям, все постсинаптические потенциалы в нейронах улитки полностью снимаются при добавлении к рингеровскому раствору ионов Со2+ в количестве 20 мМ. Так, упомянутый выше «залповик» ППа1, проявляющий довольно изменчивую спонтанную активность, после снятия синаптического притока ионами кобальта сразу же начинает в чистом виде проявлять присущие ему залпы. Блокирующее действие кобальта на химические синапсы легко отмывается. Кроме синаптического притока, на спонтанную активность клеток могут влиять факторы, меняющие состояние электрогенного насоса — например, температура. Имеются и более сложные обстоятельства, знание которых приходит с опытом работы. Так, при осторожном введении электрода в клетку, относящуюся к группе D, обычно регистрируется редкая спонтанная активность. Пропусканием деполяризующего тока можно вызвать на короткое время частый залп импульсов, вслед за чем в течение нескольких минут клетка развивает глубокую гиперполяризацию, примерно до -70 мВ, и надолго умолкает [280]. Такой же молчащей будет клетка, если частый залп был вызван резким введением в неё электрода. По-видимому, в обоих случаях имеет место стимуляция электрогенного насоса (это предположение экспериментально не проверялось). Приводя этот пример, мы хотели показать, что в данном случае характер спонтанной активности — отсутствие её или нерегулярная импульсация — зависит от незначительных и порой непроизвольных изменений в поведении экспериментатора. Важнее всего, однако, что даже и в этом сказываются характеристические особенности клеток. В отличие от клеток группы D, другие нейроны (например, клетки группы F) никогда не развивают длительную и даже кратковременную гиперполяризацию в ответ на преходящую деполяризацию. Потенциал покоя. Наш собственный опыт в этом отношении не простирается дальше наблюдений, что определенным нейронам постоянно присущи меньшие значения потенциала, чем другим. Сошлёмся на количественные данные авторов, специально занимавшихся этим вопросом при работе на идентифицированных нейронах виноградной улитки. «Молчащие» ростральные нейроны правого и левого париетальных ганглиев изучены в этом отношении независимо в трёх разных лабораториях, при этом всеми авторами даются близкие значения потенциала покоя. По А. В. Карякину [20], потенциал равен -56,6±4,2 мВ (для клетки ЛПа3, которую Карякин обозначает как ЛПа1) и -56±5,1 мВ (для ППа3). Французские авторы [171] указывают общую среднюю цифру для этих двух клеток, а также для ЛПа1, -54±6 мВ. Наконец, Н. Г. Пархоменко [45, 46] выделяет эти нейроны как клетки типа «а», характеризующиеся совокупностью биофизических характеристик, и указывает, что потенциал покоя у них обычно равен (-55) — (-65) мВ. В противоположность клеткам типа «а», клетки типа «б» имеют, по данным Н. Т. Пархоменко, потенциал покоя, равный (-35) — (-40) мВ. Судя по рисунку, приложенному к этой работе, к этому типу относятся клетки группы В. Подтверждая эти результаты Пархоменко собственным опытом, мы можем добавить, что к этому типу можно отнести и другие белые клетки, в частности нейроны групп А и D. Несмотря на ограниченность имеющихся данных, относящихся к идентифицированным клеткам, они однозначно свидетельствуют о закономерном характере различий в величинах потенциала покоя. Оценивая этот показатель, нужно, конечно, иметь в виду возможный вклад электрогенного насоса, о чём говорилось выше. Этот вопрос рассматривается в ряде специальных работ [1, 28, 113, 120 и др.]. Потенциалы действия. Параметры потенциала действия также различны у разных нейронов; кроме того, при идентификации клетки иногда важно рассматривать последовательный ряд потенциалов, так как их параметры могут характерно меняться в течение залпа. С самого начала работы по картированию идентифицированных нейронов мы придавали большое значение этому показателю и предложили разделить полные потенциалы действия, не являющиеся псевдоспайками, на две категории: имеющие задержку на нисходящей фазе потенциала и не имеющие её [283]. В то время нам пришлось выслушать немало возражений, сводящихся в основном к тому, что «задержка» появляется у любой клетки, когда она находится в плохом состоянии. Однако тот факт, что упомянутая задержка во всех без исключения случаях наблюдается у одних идентифицируемых нейронов и отсутствует у других, давал нам уверенность в своей правоте. В частности, среди клеток правого париетального ганглия такую задержку закономерно проявляют нейроны группы D и залповик ППа1. Мы отметили также, что характерной чертой потенциалов действия в этих клетках является прогрессирующее удлинение задержки на нисходящей фазе в ряду импульсов, следующих с достаточно высокой частотой. Потенциалы действия, имеющие такую форму, проявляют большой овершут иблагодаря этому отличаются значительной величиной. Последующее изучение вопроса другими авторами подтвердило эти наблюдения и показало, что различия параметров потенциалов действия тесно связаны с другими различиями между нейронами [327]. Здесь снова уместно сослаться на результаты Н. Т. Пархоменко. По его данным, задержку на нисходящей фазе потенциала действия проявляют нейроны типа «б», т. е. те идентифицируемые клетки, у которых потенциал покоя равен (-35) — (-40) мВ. По подсчётам Н. Т. Пархоменко, потенциал действия в этих клетках имеет следующие параметры: амплитуда 90-105 мВ, овершут до 60 мВ, длительность 15 — 25 мсек. В отличие от этого, в нейронах типа «а» потенциал имеет следующие параметры: амплитуда 80-90 мВ, овершут 20-35 мВ, длительность 4-9 мсек, задержка на нисходящей фазе отсутствует. Клетки двух типов различаются по вольт-амперным характеристикам, по активности внутриклеточного калия и по ряду других показателей. Наконец, что особенно существенно, Н. Т. Пархоменко убедительно показал, что в этих двух типах различны ионные механизмы генерации соматических потенциалов действия. Если нейроны, относящиеся к типу «а», инактивируются и перестают генерировать в безнатриевых растворах, то в нейронах типа «б» способность к генерации в этих условиях сохраняется благодаря входящему току ионов кальция [45, 46]. Продолжая работу в этом направлении, мы нашли, что по типу «а» себя ведут клетки группы F, быстро теряющие возбудимость в безнатриевых растворах. Клетка ППа4, медленно теряющая возбудимость, но не способная генерировать кальциевые потенциалы, представляет особый тип и, возможно, сходна с теми неидентифицированными нейронами, изученными В. Д. Герасимовым [10], которые каким-то образом используют малые количества ионов натрия для создания эффективного градиента. Наконец, изучение клеток, проявляющих кальциевые потенциалы действия, привело нас к предположению, что, возможно, эти потенциалы являются секреторными. Белые клетки, для которых особенно характерна совокупность биофизических свойств, выделенных Н. Т. Пархоменко как тип «б», являются, судя по нашим электронно-микроскопическим данным, нейросекреторными клетками с гранулами пептидергического типа. Они имеют множество соматических отростков, из которых нейросекрет, по-видимому, секретируется в оболочку ганглия или под неё. Если это так, то соматическая мембрана должна обладать свойствами секреторной, пресинаптической мембраны [24]. Любопытно, что через два года точно такой же вывод был сделан французскими авторами, исследующими нейроны аплизии; на основании совсем другой системы экспериментов они также заключили, что соматическая мембрана, судя по поведению ионов кальция, может рассматриваться как модель пресинаптической мембраны [299]. В самом деле, известно, что мембрана пресинаптического окончания в условиях ингибирования натриевой проводимости развивает при деполяризации регенеративный спайкоподобный процесс, связанный с входом в окончание ионов кальция [207]. Обращают на себя внимание черты сходства этого процесса с потенциалом действия в клетках типа «б», в частности в нейронах группы D и ППа1: задержка на нисходящей фазе, её «утомление» в ходе залпа и т. д. Отмеченными выше различиями не исчерпываются все возможные варианты потенциалов действия. Разными авторами неоднократно отмечалось, что для части клеток характерно наличие длительной, около 300 мсек, следовой гиперполяризации, имеющей тенденцию к суммации в ряду следующих друг за другом потенциалов. Этим признаком обладают, например, нейроны ППл1, ЛПл1, В4. В одних клетках (их много) соматическому потенциалу предшествует аксонная компонента, в других (например, ППа1) её нет. Имеются нейроны, в которых, напротив, сома, по-видимому, невозбудима и регистрируется только однокомпонентный потенциал действия аксонной природы. У части таких клеток амплитуда этого потенциала претерпевает резкие изменения под воздействием изменений в спонтанном синаптическом притоке (например, В6). То же, но в усложненном виде, имеет место в клетках с несколькими аксонами, где потенциалы действия генерируются в каждом из аксонов. Скопление таких клеток представлено на вентральной поверхности висцерального ганглия у выхода анального нерва. Спонтанный синаптический приток. Картина постсинаптических потенциалов, регистрируемых внутриклеточным электродом, весьма характерна для каждого нейрона, и эти характеристики закономерно повторяются от препарата к препарату. Мы не знаем в ганглиях виноградной улитки нейронов, полностью лишённых постсинаптической активности, и указания некоторых авторов на наличие таких нейронов полагаем ошибкой. В простейших случаях синаптический приток представлен однородными ВПСП. Примером могут служить нейроны группы F, причём у соседних нейронов, относящихся к этой группе, интенсивность синаптического притока бывает очень разной; обычно у этих клеток ВПСП имеют такую высокую частоту, что они, сливаясь, создают определённый уровень деполяризации, который в свою очередь определяет частоту генерации потенциалов действия. В других нейронах, получающих также лишь ВПСП, они приходят относительно редко и не сливаются; такие клетки склонны молчать или изредка генерируют, в ответ на приходящие ВПСП,нерегулярные спайки. В качестве примеров можно назвать обе гигантских клетки плевральных ганглиев. Далее, картина может осложняться тормозным притоком, который в разных клетках выражен по-разному: нерегулярные отдельные ТПСП относительно небольшой амплитуды (такие, например, характерны для многих нейронов педальных ганглиев); приходящие залпами «гигантские» ТПСП (например, в клетках В6, ППа4); ТПСП обоих названных типов в одном и том же нейроне (например, ППа1). Многими авторами наблюдались и описывались двуфазные ПСП. Что касается клеток висцерального комплекса ганглиев, здесь, по нашему мнению, двуфазными в некоторых препаратах бывают те самые ПСП, которые в других препаратах проявляют себя как «гигантские» ТПСП. По какой причине у некоторых улиток эти потенциалы лишены первой, возбуждающей фазы, нам неясно. Реакция клетки на поляризацию мембраны. Рассмотренный нами способ дифференцировать клетки на основании их ответов на инъекцию тока через внутриклеточный микроэлектрод [283] является развитием идеи, обсуждавшейся рядом авторов. Мы предложили классифицировать нейроны как осциллирующие и неосциллирующие, различая в каждой из этих категорий две подгруппы. Классификация основана прежде всего на выявлении способности или неспособности клетки длительно генерировать потенциалы действия. Инъекция деполяризующего тока является в этом случае универсальным тестом, применимым как к активным, так и к молчащим клеткам. Осциллирующие нейроны после начального частого залпа длительно удерживают активность, частота которой зависит от уровня деполяризации. Неосциллирующие после начального залпа умолкают. Активность осциллирующих нейронов бывает мономодальной и бимодальной. У мономодальных осцилляторов, в отличие от бимодальных, импульсный разряд не прерывается периодическими паузами. Пример мономодальных осцилляторов — нейроны группы F. Бимодальные осцилляторы в свою очередь можно разделить на две категории. У некоторых клеток паузами бывают разделены небольшие группы импульсов, причём в течение паузы клетка продолжает периодически генерировать пейсмекерные потенциалы, не достигающие порогового уровня для генерации потенциалов действия. Инъекция деполяризующего тока почти не влияет на частоту импульсов в группе и на их число, но делает паузы более короткими. Такие клетки генерируют пейсмекерный потенциал мономодально и лишь из-за колебаний возбудимости активность выглядит бимодальной. К этой категории относятся клетки ППа2, В5. Совершенно иной характер имеет активность в «настоящем» бимодальном осцилляторе — нейроне ППа1. Здесь разряд имеет форму периодических залпов, причём число импульсов в залпе, обычно равное при комнатной температуре 10 - 20, может быть и очень большим, до нескольких десятков. В течение залпа клетка всё более деполяризуется, затем наступает волна гиперполяризации, знаменующая собой начало паузы. В ходе паузы постепенно развивается деполяризация, вызывающая начало нового залпа и продолжающаяся до новой волны гиперполяризации.Инъекция деполяризующего тока увеличивает и продолжительность залпа, и частоту импульсов в нем. Такой тип бимодальной активности есть свойство, внутренне присущее клетке и ярко выраженное у нейрона ППа1 [283, 289]. В сглаженной форме, однако, мы изредка наблюдали такое поведение у клеток группы D. Неосциллирующие нейроны также неодинаковы. Одни из них в ответ на инъекцию деполяризующего тока умолкают после начального залпа вследствие аккомодации, т. е. без заметного сдвига мембранного потенциала. Таковы клетки группы Е. В других клетках генерация потенциалов действия инактивируется прогрессирующей деполяризацией; после выключения тока клетки долго не могут вернуться к прежнему уровню мембранного потенциала, что говорит о слабости реполяризующего механизма. Так ведут себя, например, однородно крупные клетки мезоцеребральной области церебральных ганглиев. Карпентер [112] недавно заново проанализировал сравнительные данные о различиях между осциллирующими (пейсмекерными) и неосциллирующими (непейсмекерными) нейронами, а также между разными формами эндогенной активности нейронов. Он, в частности, отмечает, что пейсмекерные нейроны гораздо слабее аккомодируются к приложенному току (о чём уже говорилось выше); мнение Карпентера о том, что у пейсмекерных нейронов потенциалы действия генерируются сомой, в отличие от непейсмекерных, генерирующих аксоном, представляется нам слишком категорическим: скорее, это верно лишь для некоторых специальных случаев. Карпентер приходит также к заключению, что ни одно из предложенных объяснений механизма медленных осцилляции, представленных в залповых нейронах, нельзя признать удовлетворительным. В связи с этим интересны данные о том, что у нейронов, имеющих генератор медленных волн (т. е. у залповиков), постоянная времени мембраны, измеряемая на толчках гиперполяризующего тока, примерно в пять раз больше, чем у незалповых нейронов того же препарата (эксперименты на нейронах гастропод); эти различия исчезают в условиях охлаждения (15° и ниже), когда перестаёт работать и генератор медленных волн [325]. Биофизические характеристики клеточной мембраны. Не рискуя рассуждать о предмете, требующем специальных знаний, ограничимся несколькими замечаниями. Н. Т. Пархоменко [45, 46] обнаружил достоверную связь между входным сопротивлением нейрона и его способностью к генерации потенциалов действия в безнатриевых растворах; эта способность, как отмечено выше, совершенно различна в разных идентифицируемых клетках. Тот же автор отметил, что клетки примерно одинакового размера могут сильно различаться по величине входной ёмкости, и отнёс эти различия за счёт разной площади поверхностной мембраны (вообще, нередко площадь мембраны, а вслед за тем удельное сопротивление вычисляют основываясь на измерении постоянной времени). Здесь, однако, нужна осторожность, учитывая хотя бы только что упомянутые данные о температурной зависимости постоянной времени мембраны у залповых нейронов. Если говорить о конкретных клетках, то, по нашим наблюдениям, постоянная времени велика у клеток пептидергического типа (ППа1, группы А, В, D). Известно, далее, что в ганглиях моллюсков мембраны разных нейронов различаются вольт-амперной характеристикой. Кандель и Тауц [205], описавшие для гигантской метацеребральной клетки особую зависимость ВПСП от мембранного потенциала, связали это свойство с аномальными выпрямляющими свойствами клеточной мембраны. В отличие от «стандартного» поведения, когда амплитуда ВПСП растёт с увеличением мембранного потенциала (таковы, по нашим наблюдениям, клетки ЛПа3, ППа3, В6 и мн. др.), в метацеребральном нейроне величина ВПСП вблизи потенциала покоя увеличивается, когда поляризация клетки уменьшается. Для таких ВПСП характерно также, что их продолжительность уменьшается при гиперполяризации и увеличивается при деполяризации. Поскольку аномальное выпрямление представлено в совершенно определенных клетках и, по-видимому, коррелирует с другими свойствами нейрона, в частности рецепторными [168], по поведению ВПСП при поляризации можно судить, к какой из двух категорий относится исследуемая клетка.
4. 2. 4. Специфичность клеточных рецепторов
В 1961 г. Тауц и Гершенфельд обнаружили, что в ганглии моллюска одни нейроны отвечают на ацетилхолин деполяризацией и возбуждением, а другие — гиперполяризацией и торможением; было предложено различать две категории нейронов, D и Н [308]. Это открытие породило огромную литературу, и по мере детализации знании о реакциях нейронов на медиаторы росло число «фармакологических типов» нейронов. Работа на идентифицированных клетках дала возможность понять, что нейрон обладает определёнными, повторяющимися от препарата к препарату, рецепторными свойствами. По сложившейся традиции, говорят о «фармакологических характеристиках» нейронов, подразумевая, во-первых, знак ответа на тот или иной медиатор; во-вторых, ионный механизм этого ответа; и, в-третьих, отношение соответствующего рецептора к литикам и миметикам. Имеется ряд работ, выполненных на нейронах гастропод, которые дают широкое представление о типах ответов клеточных мембран на апплицируемые медиаторные вещества. Ацетилхолин. До сих пор физиологами найдено пять способов действия ацетилхолина на поверхностную клеточную мембрану: 1) деполяризация повышением проницаемости для натрия; 2) деполяризация понижением проницаемости для калия; 3) гиперполяризация повышением проницаемости для калия; 4) гиперполяризация повышением проницаемости для хлора, при низкой внутриклеточной концентрации этого иона; 5) деполяризация повышением проницаемости для хлора, при высокой его внутриклеточной концентрации. Из пяти способов четыре представлены в нейронах моллюсков [212], исключение составляет лишь второй из перечисленных механизмов, постулированный для клеток мозга и симпатических ганглиев млекопитающих [224, 329]. Высказывается мнение, что в ганглиях моллюсков можно найти ещё два механизма: стимуляция и торможение ацетилхолином электрогенного насоса [212], но в этом отношении пока нет строгих экспериментальных фактов. На виноградной улитке клеточные эффекты ацетилхолина исследовались нами на поверхностных нейронах всех ганглиев подглоточного комплекса. В этих опытах (и в других наших электрофизиологических экспериментах, о которых речь будет идти позже) применялась обычная методика регистрации трансмембранного потенциала капиллярным внутриклеточным микроэлектродом. Регистрирующий электрод мы, как правило, заполняли 2М цитратом калия (в специальных случаях, когда требовалось инъецировать в клетку ионы хлора, отводящий электрод заполняли 2,5М хлористым калием). Изолированное окологлоточное кольцо или часть его помещали в проточную камеру объемом 10 мл. Раствор Рингера для улитки имел следующий состав (в мМ): хлористый натрий — 80, хлористый калий — 4, хлористый кальций — 7, хлористый магний — 5, рН доводили до 7,4 с помощью Трис-хлорида. Трис использовали и для компенсации осмотичности раствора при удалении тех или иных катионов. Поляризацию клеточной мембраны осуществляли через отводящий электрод, используя мостовую схему, или через второй внутриклеточный электрод. Эффекты ацетилхолина только на раннем этапе исследования наблюдали в условиях внесения его в окружающий раствор, в основной части экспериментов применяли стандартную методику ионофоретической аппликации из подведенного к клетке капиллярного электрода. Знак ответа нервной клетки на ацетилхолин (как и на другое медиаторное вещество) может в естественных условиях быть непостоянным вследствие того, что мембранный потенциал нейрона иногда меняется в широком диапазоне. Мы столкнулись с этой трудностью при изучении группы нейросекреторных клеток правого париетального ганглия: этим клеткам свойственно развивать иногда глубокую гиперполяризацию, на фоне которой эффект ацетилхолина, обычно гиперполяризующий и тормозящий активность, становится деполяризующим, хотя и не до уровня генерации [24, 280]. Результаты испытания медиаторных веществ можно стандартизировать при условии, что знак реакции оценивается при значении мембранного потенциала, пороговом для генерации потенциалов действия. Такое условие мы стремились выполнять в данном исследовании. На рис. 6 суммированы результаты, касающиеся знака ответа на ацетилхолин нейронов, находящихся в разных ганглиях подглоточного комплекса. Данные, относящиеся к картированным индивидуальным нейронам, на этих схемах не отражены, они изложены в разделе 4.2.5. Рисунки дают представление о региональных особенностях ответов нейронов на ацетилхолин. Так, на вентральной поверхности педальных ганглиев и в той области правого париетального ганглия, где расположены крупные нейросекреторные клетки (группа D), ацетилхолин почти без исключений вызывает гиперполяризацию. Противоположные, деполяризующие эффекты наблюдаются, как правило, на дорзальной поверхности висцерального ганглия в зоне, занятой преимущественно нейронами группы F. Несколько более пёстрые результаты, полученные в аналогичных условиях голландскими авторами, которые изучали ответы нейронов, расположенных на дорзальной поверхности комплекса [346], объясняются, по всей вероятности, тем, что этими авторами при оценке знака ответа не выдерживалось отмеченное выше стандартизирующее условие.

Рис. 6. Знак ответа на ацетилхолин со стороны нейронов, расположенных в разных участках ЦНС виноградной улитки. А — ганглии висцеральной дуги; Б, В — педальные ганглии с дорзальной и вентральной стороны. На схемах суммированы результаты разных опытов, но данные, полученные на индивидуально идентифицируемых клетках, в рисунок не включены и сообщаются в тексте (то же относится к рис. 7). Ионофоретическая аппликация. Заштрихованный кружок указывает позицию клетки, деполяризуемой медиатором, чёрный кружок — гиперполяризуемой клетки.
Об особом, «двойном» действии ацетилхолина на некоторые нейроны будет сказано ниже, при описании группы G (4.2.5.). Ионные механизмы эффектов ацетилхолина были нами исследованы в нескольких случаях. Деполяризующие эффекты оказались натрийзависимыми и гиперполяризующие — зависимыми от ионов хлора [24]. Пока не обнаруживались другие типы эффектов, найденные на нейронах других моллюсков. В литературе имеются данные о том, что холинорецепторы, ответственные за разные ионные эффекты ацетилхолина на нейроны гастропод, имеют разное строение, что выражается их отношением к холинолитикам и холиномиметикам [208, 232]. Первичные катехоламины. Признано, что дофамин, подобно ацетилхолину, выполняет медиаторные функции в нервной системе гастропод. Как правило, чувствительные к дофамину нейроны моллюсков, в частности аплизии, отвечают на него гиперполяризацией, но бывают и противоположные эффекты [83]. Глайзнер, по-видимому, впервые нашел у садовой улитки клетку, реагирующую на дофамин деполяризацией [170]. У виноградной улитки нам удалось исследовать в этом отношении только поверхностные нейроны задне-медиальной области правого париетального ганглия (дорзальная клеточная кора). В этой области большинство клеток, в частности нейроны ППа1, ППа2 и клетки группы D, гиперполяризуются и тормозятся дофамином, но имеется также несколько небольших клеток, отвечающих на дофамин деполяризацией. Чувствительностью к норадреналину обладают, как правило, те же нейроны садовой улитки, которые реагируют на дофамин [170]; для виноградной улитки собственными данными по действию норадреналина мы не располагаем. Серотонин. В литературе детальные данные о клеточных эффектах серотонина имеются только для садовой улитки Helix (Cryptomphallus) aspersa. Сначала Гершенфельду и Стефани удалось заметить только возбуждающие эффекты, которые развивались с большой латентностью, из чего авторы сделали вывод, что серотонин действует на расстоянии от тела нейрона [168]. Позже Глайзнер сообщил, что у этого вида имеются также нейроны, отвечающие на серотонин торможением [170], а Гершенфельд нашел, что тормозные эффекты имеют разную ионную и рецепторную природу в правом и левом париетальном ганглиях [165]. Все эти исследования проводились на дорзальной поверхности ганглиев подглоточного комплекса. У виноградной улитки мы с Я. Шаланки также описали сначала только возбуждающие эффекты серотонина [283]. В ходе дальнейшей работы мы с Г. Н. Коробцовым нашли в этой и других областях ЦИС виноградной улитки разные типы ответов на ионофоретически апплицируемый серотонин [25]. Отдельные клетки, отвечающие на серотонин торможением, были обнаружены в левом и правом париетальных ганглиях, т. е. там, где у садовой улитки их нашел Гершенфельд.
 Рис. 7. Знак ответа на серотонин со стороны нейронов, расположенных в разных участках ЦНС виноградной улитки.
А, Б — ганглии висцеральной дуги; В, Г — педальные ганглии с дорзальной и вентральной стороны соответственно. Условия те же, что на рис. 6.
Рис. 7. Знак ответа на серотонин со стороны нейронов, расположенных в разных участках ЦНС виноградной улитки.
А, Б — ганглии висцеральной дуги; В, Г — педальные ганглии с дорзальной и вентральной стороны соответственно. Условия те же, что на рис. 6.
 Рис. 8. Противоположные по знаку ответы клеток висцерального ганглия (дорзальная поверхность) на серотонин (ионофоретическая аппликация).
А — клетка, деполяризуемая серотонином; Б — клетка, гиперполяризуемая серотонином (остановка спонтанной активности); В — та же клетка при более негативном значении мембранного потенциала: генерация отсутствует, серотонин гиперполяризует.
Рис. 8. Противоположные по знаку ответы клеток висцерального ганглия (дорзальная поверхность) на серотонин (ионофоретическая аппликация).
А — клетка, деполяризуемая серотонином; Б — клетка, гиперполяризуемая серотонином (остановка спонтанной активности); В — та же клетка при более негативном значении мембранного потенциала: генерация отсутствует, серотонин гиперполяризует.
Кроме того, в висцеральном ганглии найдена большая, компактно расположенная группа клеток, у которых серотонин тоже вызывает гиперполяризацию. Эта группа (группа G) хорошо представлена на вентральной поверхности ганглия, но выходит и на дорзальную поверхность. Результаты ионофоретической аппликации серотонина на нейроны этого ганглия схематически подытожены на рис. 7, а на рис. 8 представлены записи эффектов серотонина на клетки ганглия. В педальном ганглии эффекты серотонина оказались в основном деполяризующими, но и здесь были обнаружены отдельные нейроны, отвечающие гиперполяризацией (рис. 7, Г). Ионный механизм был изучен только на тех клетках, которые отвечают на серотонин деполяризацией: эффект всегда оказывался натрийзависимым. Интересно отметить, что это относится и к гигантским метацеребральным нейронам, которые хорошо отвечают на серотонин несмотря на то, что сами являются серотонинергическими (см. 4.2.5). Подробнее о действии серотонина на идентифицированные клетки см. ниже. Здесь отметим лишь, что стимулирующие эффекты серотонина, вероятно, могут иметь разные механизмы. По нашим наблюдениям, одни нейроны отвечают на серотонин быстро развивающейся деполяризацией, которая прекращается вслед за прекращением аппликации серотонина. Таковы, например, гигантские метацеребральные нейроны (клетки ПЦ1 и ЛЦ1 — см. 4.2.5.). В других случаях стимуляция характеризуется большой латентностью, длительным последействием и как бы избирательно направлена на осциллогенез; такое действие серотонина нельзя имитировать деполяризующим током. Этот тип эффекта наблюдается у нейронов пептидергического типа — группы А, В, D, крупные белые клетки ганглиев висцеральной дуги [26]; он выражен и у некоторых педальных нейронов, которые, по-видимому, не являются пептидергическими.
4. 2. 5. Идентифицированные клетки
ЛПл1 и ППл1. Эти симметрично расположенные гигантские клетки проявляют сходные физиологические свойства. Размеры в фиксированном материале до 150 мк, живые клетки немного крупнее. Чаще всего расположены на дорзальной поверхности ганглиев, но иногда позиция тела клетки смещается. Клетки пигментированные, каждая из них выделяется на фоне окружающих мелких нейронов. Визуальная идентификация проста. Обе клетки склонны молчать или генерируют редкие и нерегулярные потенциалы действия в ответ на спонтанные ВПСП. При пропускании слабого деполяризующего тока легко выявляются осцилляторные свойства: появляется регулярная активность мономодального характера без аккомодации к току. ТПСП в спонтанном синаптическом притоке никогда не наблюдались. Потенциал покоя 45-50, у молчащих нейронов до 60 мВ; потенциал действия 2-компонентный — первый, меньший компонент соответствует, по-видимому, аксонной триггерной зоне, тогда как второй длительностью около 10 мсек и амплитудой около 65-70 мВ, т. е. с небольшим овершутом, является соматическим. Следовая гиперполяризация, как уже отмечалось, бывает значительной величины [283]. Ацетилхолин и серотонин оказывают умеренное деполяризующее действие с выраженной десенситизацией; дофамин как будто неэффективен (эти наблюдения сделаны только на правой клетке). ЛПа1. Единственный гигантский нейрон, расположенный на заднем конце левого париетального ганглия, у выхода левого мантийного нерва. Чаще всего клетка лежит несколько правее нерва, иногда над ним. В этой области граница между левым париетальным и висцеральным ганглиями изменчива, так что в редких случаях клетка оказывается по другую сторону щели, т. е. в висцеральном ганглии. Таким образом, клетку очень просто идентифицировать визуально. Когда позиция этого нейрона нетипична и он переходит в висцеральный ганглий, его легко отличить от «собственных» нейронов висцерального ганглия по отсутствию активности. При прямом возбуждении пропусканием деполяризующего тока клетка генерирует, как правило, псевдоспайки, при этом залп быстро прекращается вследствие аккомодации к току. В спонтанном синаптическом притоке наблюдаются только мелкие ВПСП. Действие медиаторов не изучено. Клетка ЛПа1, вместе с ЛПа3 и ППа3, была объектом исследования французских авторов, применявших, в частности, метод фиксации потенциала. Эти авторы дают следующие, средние для всех трёх нейронов, цифры: потенциал покоя — 54±6 мВ (при 23-25°), потенциал действия 92 мВ, трансмембранное сопротивление 1 — 2,5 МОм, постоянная времени 40-50 мсек, откуда входная ёмкость равна около 25 нФ [171]. ЛПа2. Гигантская клетка, расположенная на переднем конце ганглия. В этой области картина крупных нейронов отличается значительным непостоянством: по данным Кунце, на дорзальной стороне в разных препаратах можно видеть до четырёх и на вентральной до трёх гигантских нейронов. Одна из этих клеток белая, что облегчает её индентификацию; белая клетка чаще всего лежит на вентральной стороне ганглия, но иногда заходит на дорзальную, располагаясь медиально. Остальные, пигментированные, гигантские нейроны приходится различать, учитывая их относительную позицию и физиологические свойства. Клетка ЛПа2 всегда расположена латеральнее «молчащего» нейрона ЛПа3. ЛПа2 проявляет редкую активность или молчит, но, в отличие от ЛПа3, хорошо поддерживает активность при пропускании деполяризующего тока и нередко генерирует в ответ на спонтанные залпы ВПСП. Другое отличие от ЛПа3 — наличие в спонтанном синаптическом притоке как возбуждающих, так и тормозных ПСП. Залпы ТПСП кумулируются в торможение большой длительности. Потенциал покоя около 40- 50 мВ, потенциалы действия с овершутом, без задержки на нисходящей фазе. Ацетилхолин на уровне генерации потенциалов действия гиперполяризует, этот эффект извращается после инъекции в клетку ионов хлора. ЛПа3 и ППа3. А. В. Карякин, изучавший эти клетки [20], считает их парными нейронами, обладающими сходным и симметричным ветвлением аксонов. Приведём сообщённые этим автором характеристики клеток: размеры около 120 мк; потенциал покоя 56,7±4,7 мВ (температура не указана); потенциал действия 94,8±8,3 мВ, длительностью 5 мсек, однокомпонентный при прямом раздражении и двухкомпонентный при антидромном; действие ацетилхолина деполяризующее; синаптический приток только возбуждающий, ВПСП мелкие, амплитудой 2-3 мВ, длительностью 40-50 мсек, амплитуда ВПСП линейно зависит от величины мембранного потенциала, рассчитанный равновесный потенциал для ВПСП равен -30 мВ. Мы можем к этому добавить, что обе клетки легко аккомодируются к деполяризующему току и быстро перестают генерировать, так что называть их нейронами пейсмекерного типа, как это делает А. В. Карякин, на наш взгляд, неверно. Нуждаются в проверке и данные о действии ацетилхолина, полученные А. В. Карякиным только в условиях перфузии всего ганглия. В немногих опытах с ионофоретической аппликацией мы наблюдали, что деполяризация, вызванная ацетилхолином, не достигает уровня генерации потенциалов действия; будучи апплицированным на уровне генерации, ацетилхолин вызывал в наших экспериментах гиперполяризацию мембраны (сделано только на клетке ЛПа3). (См. также об этих клетках в работах 120 и 171]. ППа1. В заднемедиальной доле правого париетального ганглия обычно имеется несколько гигантских нейронов, число и позиции которых варьируют. С полной уверенностью поэтому клетку ППа1 визуально идентифицировать нельзя, но всё же при достаточном опыте её можно узнать, руководствуясь следующими признаками: во-первых, это одна из крупнейших, а иногда крупнейшая клетка доли; во-вторых, она всегда примыкает к правой границе доли, т. е. к складке, отделяющей долю от латеральной области ганглия; в-третьих, клетка обычно выглядит более бледной и прозрачной, чем другие гигантские нейроны доли, хорошо пигментированные; обычно оранжеватая клетка ППа1 бывает белесоватого оттенка. Я остановлюсь на этой клетке несколько подробнее, так как она понадобится нам в следующей главе. Физиологические свойства клетки ППа1 настолько своеобразны, что позволяют не спутать её ни с каким другим нейроном. Она имеет совершенно особый тип залповой активности, подробно описанный выше (4.2.3.).
 Рис. 9. Характеристики эндогенной залповой активности нейрона ППа1 [Из 289].
А - изменение формы потенциалов действия в течение залпа (цифрой указан номер потенциала в залпе). Б — изменение длительности потенциала действия в течение залпа (показателем длительности служит интервал между восходящим и нисходящим фронтом потенциала действия при 50% его амплитуды). На абсциссе — номер потенциала, на ординате — время, мсек. В — изменение межспайкового интервала в течение залпа. На абсциссе — номер интервала, на ординате — время, сек.
Рис. 9. Характеристики эндогенной залповой активности нейрона ППа1 [Из 289].
А - изменение формы потенциалов действия в течение залпа (цифрой указан номер потенциала в залпе). Б — изменение длительности потенциала действия в течение залпа (показателем длительности служит интервал между восходящим и нисходящим фронтом потенциала действия при 50% его амплитуды). На абсциссе — номер потенциала, на ординате — время, мсек. В — изменение межспайкового интервала в течение залпа. На абсциссе — номер интервала, на ординате — время, сек.
Повторим ещё раз, что потенциалы действия крупные, до 100 мВ, с большим овершутом и задержкой на нисходящей фазе, причём длительность задержки меняется в ходе залпа. Потенциалы генерируются сомой и не имеют аксонного компонента, что также довольно редкое свойство [283]. Клетка ППа1 сильно деполяризуется ацетилхолином, серотонин тоже оказывает на неё возбуждающее действие. Оба медиатора становятся неэффективными в безнатриевой среде, тогда как способность генерировать потенциалы действия в этих условиях сохраняется (прослежено в течение не менее получаса после исчезновения реакции на возбуждающие медиаторы). Удаление ионов кальция из безнатриевого раствора вызывает падение амплитуды потенциалов действия и вскоре полное подавление возбудимости [24]. Дофамин гиперполяризует клетку, вызывая торможение активности. Необычный тип эндогенной активности, проявляемый нейроном ППа1, привлекает к этому нейрону интерес многих физиологов. Из публикаций последнего времени отметим работу Христофферсена, изучавшего на этом нейроне (а также на ЛПа3) вклад электрогенного насоса в мембранный потенциал [120], и исследование Шаланки с соавторами, которые детально изучили параметры залповой активности [289]. Эти авторы нашли, что при 22° залп состоит из 10 - 20 спайков, причем межспайковые интервалы находятся в пределах 0,3-1,5 сек., длительность залпов составляет 4,0+0,3 сек., а межзалповых пауз — 5,7± 1,5 сек. Величина мембранного потенциала перед спайком составляет 55±5 мв, сразу после залпа до 64+3 мв, амплитуда потенциала действия около 75 мв (рис. 9). Последняя величина меньше наблюдавшейся нами, что, возможно, обусловлено различиями в составе раствора Рингера. Также, в противоположность нашему опыту, Шаланки с соавторами отмечают, что залповая активность клетки ППа1 обычно представлена в чистом виде, не замаскированная искажениями, которые создаются синаптическим притоком (этими авторами отмечались лишь ВПСП, и то в небольшом количестве и не всегда). Мы, напротив, всегда наблюдаем богатый синаптический приток, включающий, как уже отмечалось выше, не только ВПСП, но и разные ТПСП, в том числе «гигантские», которые в некоторых препаратах бывают двуфазными (первая фаза деполяризующая, вторая гиперполяризующая, причем в ряду таких ПСП кумулируется гиперполяризация). Эта синаптическая бомбардировка в наших экспериментах, как правило, лишает активность того регулярного рисунка, который наблюдается у этой клетки после снятия ПСП. Поскольку несомненно, что Шаланки и соавторы работали с той же клеткой, которую ранее мы с Шаланки картировали, присвоив ей индекс ППа1, причину различий нужно искать в каких-то условиях эксперимента или препаровки. Ещё одно расхождение касается хода отростка клетки ППа1. Основываясь на результатах исследования срезов после инъекции в ППа1 проционового желтого, Шаланки с соавторами заключают, что отросток не выходит за пределы правого париетального ганглия. По моим наблюдениям, аксон ППа1 покидает этот ганглий и переходит по комиссуре в висцеральный ганглий. Более точные сведения, полученные на других хелицидах, показывают, что из висцерального ганглия аксон затем выходит в составе интестинального нерва [162б, 210]. Гайнер, нашедший, что аксон этой клетки доходит до предсердия, приводит веские физиологические и биохимические доводы в пользу предположения, что ППа1 — пептидергическая нейросекреторная клетка, для которой предсердие служит местом выделения особого, синтезируемого только этим нейроном нейрогормона в кровоток [162а, б]. ППа2. Клетка, по размерам сравнимая с ППа1, т. е. гигантская, оранжевато-прозрачная, всегда лишённая белого материала, часто пигментированная. В отличие от ППа1, которая обладает уникальным набором характеристик, ППа2 как будто является представителем группы нейронов, проявляющих сходные свойства и расположенных по обе стороны щели, разделяющей правый париетальный и висцеральный ганглии. Другие, неидентифицированные члены этой группы, имеют меньшие размеры. В висцеральном ганглии к этой группе, возможно, относится клетка В5. Для ППа2 и сходных с ним нейронов характерны потенциал покоя величиной 35-45 мВ, потенциалы действия с аксонным и соматическим компонентами и без задержки на нисходящей фазе (амплитуда соматического потенциала около 70- 80 мВ) отсутствие чувствительности к ацетилхолину. Фоновая активность ППа2 изменчива, на инъекцию деполяризующего тока клетка отвечает активностью бимодального характера, тип которой описан в разделе 4.2.3. Серотонин возбуждает эту клетку, дофамин тормозит её. В спонтанном синаптическом притоке видны отдельные и залповые ВПСП, имеются также ТПСП. ППа3 - описана вместе с ЛПа3. ППа4. Как правило, эта клетка имеет меньшие размеры, чем ППа1, ППа2 и ППа3. Её, однако, обычно легко узнать благодаря тому, что у медиальной границы скопления бледных клеток группы D эта клетка наиболее богата пигментом. Как правило, клетка активна, потенциалы действия генерируются с частотой около 1 в сек с очень нерегулярными интервалами. Они имеют довольно характерный вид: спайк возникает без предшествующей фазы медленной деполяризации, он быстр (не более 10 мсек), иногда генерируются сдвоенные спайки, разделенные короткой паузой. Спайки однокомпонентные, амплитудой до 80 мВ, при гиперполяризации амплитуда уменьшается. По-видимому, генерация имеет место только в аксоне, а сома этой клетки невозбудима; при генерации сдвоенных спайков, вероятно, в активность вовлекается вторая аксонная ветвь. Эти признаки активности, наряду с очень крупными ВПСП, резко отличают клетку ППа4 от лежащих поблизости нейронов группы D и клетки ППа1. Упомянем другие, выявленные нами свойства клетки [24]. Иногда, хотя и редко, в ней наблюдаются залпы гигантских ТПСП, вызывающие остановку активности. При пропускании деполяризующего тока клетка ведёт себя как мономодальный осциллятор. Ацетилхолин вызывает очень сильную деполяризацию, которая сменяется фазой гиперполяризации; интересно отметить, что такой тормозной фазы не бывает после деполяризации, вызванной инъекцией тока, но она бывает после деполяризации, вызванной залпом ВПСП. Возможно, клетка снабжена двумя родами холинорецепторов. Атропин не снимает действия ацетилхолина. Серотонин деполяризует и возбуждает эту клетку. Исключение ионов натрия из омывающей среды приводит к прекращению генерации потенциалов действия, а также к снятию деполяризующих эффектов ацетилхолина и серотонина. Весьма характерно, что такое влияние безнатриевой среды развивается очень медленно, в течение около 1,5 час. протока, а при остановке протока клетка быстро восстанавливает способность генерировать потенциалы действия. Такой тип поведения некоторых неидентифицированных нейронов подробно исследовал на виноградной улитке В. Д. Герасимов [10]. При исключении ионов кальция генерируется учащённая регулярная активность, амплитуда потенциалов действия уменьшается. ППа5. Эта крупная пигментированная клетка представлена не во всех препаратах. Она расположена у медиальной границы ганглия на уровне вершины висцерального ганглия и отличается от остальных идентифицируемых и картированных нейронов ганглия своей тормозной реакцией на ацетилхолин, который гиперполяризует эту клетку. Исследована она слабо. Потенциал покоя до 50 мВ, потенциалы действия более 75 мВ, обычно имеется активность, иногда довольно регулярная, но чаще изменённая синаптическим притоком, в котором различимы ВПСП (они растут при гиперполяризации) и залпы гигантских ТПСП. На фоне торможения, вызванного последними, действие ацетилхолина инвертируется. В1. Очень крупная, прозрачная, умеренно пигментированная клетка, которая хорошо видна, если висцеральный ганглий повернут к наблюдателю задне-правой стороной, где она занимает крайнее угловое положение. В некоторых препаратах это угловое положение занимает крупная белая клетка, которая чаще лежит на вентральной стороне ганглия, и тогда В1 несколько смещена. Клетка не проявляет спонтанной активности, а её реакция на деполяризацию бывает двух форм, что, возможно, связано с разными состояниями клетки. В одних случаях в ответ на инъекцию деполяризующего тока генерируется небольшое число потенциалов действия, их следовые гиперполяризации, суммируясь, гасят генерацию. В других препаратах, встречающихся чаще, в течение и после такого залпа мембранный потенциал непрерывно и медленно уменьшается. На некоторой фазе этого падения пауза, вызванная прекращением генерации потенциалов действия, вдруг сменяется серией быстрых осцилляций, амплитуда которых в разных случаях бывает различной. Эта реакция, найденная нами в клетке В1, не наблюдалась более ни у какого другого нейрона улитки; её подробно исследовал Э. Лабош, посвятивший этому вопросу специальную статью [228]. В остальных отношениях о клетке В1 известно довольно мало. Величина потенциала покоя 45-50 мВ, потенциалы действия около 80 мВ длительностью 15 мсек. В спонтанном синаптическом притоке мы наблюдали ВПСП, амплитуда которых росла при увеличении мембранного потенциала. Ацетилхолин вызывает эффекты, подобные эффектам деполяризующего тока. В2. Прозрачная, непигментированная клетка средних размеров, лежащая поверх корешка интестинальнго нерва и окружённая мелкими пигментированными нейронами. Единственная в своей области, эта клетка проявляет частую спонтанную активность, что позволяет идентифицировать её с большой уверенностью. Активность нерегулярная, с перебоями, с меняющейся в ходе залпа амплитудой потенциалов действия. В3. Крупная белая клетка, представленная не во всех препаратах и, по-видимому, являющаяся одним из нейронов белой группы А. Чаще всего нерегулярно активна, потенциалы действия двухкомпонентные, с большим овершутом, с задержкой нанисходящей фазе соматического компонента. Имеются ВПСП иТПСП. Ацетилхолин гиперполяризует, серотонин стимулирует. В4. Клетка, отличающаяся очень крупными размерами, имеется в указанном на схеме месте у многих, но не у всех особей. Трудно с полной достоверностью утверждать, что это всегда одна и та же клетка, так как её физиологические свойства характерны для целой группы крупных нейронов, имеющихся в этой области (группа F). В5. Очень крупная клетка, встречающаяся во многих препаратах. По своим физиологическим характеристикам соответствует клетке ППа2. В6. Зрительная идентификация этой клетки трудна, хотя в общем можно сказать, что клетка занимает позицию, ближайшую к левому париетальному ганглию, на краю выпуклой, нечётко обозначенной дольки в задней части висцерального ганглия. Физиологически она легко отличается от окружающих нейронов, в большинстве своём относящихся к группе F. Клетку В6 характеризует меняющийся рисунок активности и богатство синаптического притока, в котором выделяются крупные ВПСП и приходящие сериями крупные ТПСП. Иногда возбуждающий приток настолько силен, что он создает постоянную деполяризацию, достаточную для частой и регулярной авторитмичности. Такие периоды могут сменяться периодами полного молчания, вызываемыми залпами ТПСП, или же периодами, когда отдельные нерегулярные спайки генерируются в ответ на отдельные ВПСП. Для этой клетки, как и для В2, к которой она близка физиологически, характерна изменчивость амплитуды потенциалов действия, что, по-видимому, указывает на их аксонное происхождение. Группы А и В. Белые клетки, расположенные в области переднего конца висцерального ганглия так, что часть их (группа А) попадает в состав висцерального, а часть (группа В) — в состав левого париетального ганглия. Это нейроны, имеющие потенциал действия с аксонным и соматическим компонентами, причем у последнего большой овершут и задержка на нисходящей фазе. К ним приложимы характеристики, указанные Пархоменко для нейронов типа «а» (см. 4.2.3.). Размеры клеток изменчивые, но за редким исключением не очень большие (не более 50-60 мк). Судя по описанию, именно с этими клетками работала на садовой улитке Ф. Вальд [325а], нашедшая, что натрий-зависимый аксонный потенциал действия отличается у них от преимущественно кальциевого соматического потенциала,- полное повторение вывода, сделанного до этого Н. Т. Пархоменко [46]. Клетки этих групп обычно имеют нерегулярную активность. При деполяризации ведут себя как мономодальные осцилляторы. Имеют ВПСП и отдельные некрупные ТПСП. Ацетилхолин гиперполяризует, это действие извращается после инъекции ионов хлора. Серотонин, деполяризуя, стимулирует пейсмекерный механизм. Группа С. Небольшое число крупных белых клеток, расположенных вдоль медиальной границы левого париетального ганглия. Исследованы слабо и как будто соответствуют по своим характеристикам клеткам групп А и В. Группа D. Бледные, реже белые клетки размером до 150 мк, образующие выпуклую долю в правом париетальном ганглии. Размеры и позиция доли изменчивы (особенно в передне-заднем направлении), иногда складка делит её на две доли. Но при всей изменчивости позиций эти клетки всегда лежат латеральнее области, занятой клетками ППа1, ППа2 и ППа4. Число клеток этой группы — около 30. Эта группа изучалась нами довольно детально как электрофизиологически, так и электронно-микроскопически. Вкратце, физиологические свойства нейронов следующие. По мембранным характеристикам эти клетки соответствуют типу «а», по Н. Т. Пархоменко (см. 4.2.3.). Потенциал действия двухкомпонентный, причём в активности иногда представлен только аксонный компонент («псевдоспайк»), но чаще имеется и соматический компонент с большим овершутом и задержкой на нисходящей фазе. Длительность этого потенциала изменчива, но как будто не менее 30 мсек. Клетки молчат или имеют редкую активность. После залпа, вызванного деполяризацией мембраны, в течение нескольких минут клетка может развить глубокую, до -70 мВ, гиперполяризацию, на фоне которой извращается действие ацетилхолина (он гиперполяризует и тормозит клетку на уровне генерации активности). Серотонин деполяризует и снимает способность гиперполяризоваться после активности; на фоне серотонина клетка ведёт себя как мономодальный осциллятор. Дофамин гиперполяризует. Клетки группы D обладают отчетливо выраженной способностью генерировать потенциалы действия в безнатриевой среде. Нужно заметить, что в протоке безнатриевого Трис-рингера возбуждающее действие серотонина снимается за несколько минут, т. е. ионы натрия действительно вымываются из околоклеточной среды. Между тем потенциалы действия продолжают генерироваться (проверено в течение более двух часов непрерывного протока, а также в ганглиях, выдержанных в течение 4-6 час. в сменяемом безнатриевом рингере). В этих условиях исключение из раствора ионов кальция или добавление 20 мМ Со2+, блокирующего кальциевые каналы, приводит к быстрому уменьшению потенциалов действия и затем подавлению их генерации. Гиперполяризующее действие ацетилхолина связано с повышением проницаемости для ионов хлора. Группа Е. Некомпактное скопление относительно небольших, пигментированных клеток, лежащее в правом париетальном ганглии позади коннектива, связывающего этот ганглий с висцеральным. Отчасти лежат между крупными и гигантскими нейронами. Характерный признак — быстрая аккомодация к деполяризующему току, благодаря чему у этих клеток нельзя получить длительно поддерживаемой активности. Ацетилхолин их деполяризует. Возможно, клетки с такими же свойствами лежат в примыкающей области висцерального ганглия. Группа F. Крупные пигментированные нейроны, лежащие в заднелатеральной части висцерального ганглия (где имеются и клетки с другими свойствами). Для нейронов этой группы характерна довольно регулярная активность, которая определяется богатым притоком ВПСП, создающим постоянный фон деполяризации. ТПСП нет. Потенциалы действия двухкомпонентные, не очень большие (до 50-70 мВ), с небольшим овершутом. Ацетилхолин деполяризует, учащая разряд. Как генерация потенциалов действия, так и эффект ацетилхолина снимаются в течение нескольких минут в протоке безнатриевого Трис-рингера. Серотонин деполяризует. Группа G. Крупных или средних размеров клетки, часто с многокомпонентной спайковой активностью. Создаётся впечатление, что генерация потенциалов действия имеет место в нескольких аксонных ветвях, тогда как сома неактивна. Характерный признак — тормозное действие серотонина (у зимних улиток). Клетки этой группы на дорзальной поверхности висцерального ганглия не образуют компактной группы и смешаны с другими нейронами, в частности, с относящимися к группе F. Напротив, на вентральной поверхности ганглия в той его дольке, которая находится между корешком анального нерва и левым плевральным ганглием, почти все поверхностные нейроны относятся к этой группе. Расположенные здесь клетки довольно крупные, а одна из них, лежащая у латерального края доли, — гигантская, её легко визуально идентифицировать, и можно зарезервировать за этой клеткой индекс В7. На этой гигантской клетке мы с Г. Н. Коробцовым неоднократно отмечали двойной эффект ацетилхолина — тормозной гилерполяризующий при аппликации на сому и деполяризующий, возбуждающий при аппликации на отросток на расстоянии от сомы. Пока неясно, все ли клетки группы обладают двумя механизмами холинореактивности, как эта гигантская клетка, но по крайней мере некоторые неидентифицируемые нейроны этой области, вероятно, разделяют это свойство. Гигантские метацеребральные нейроны ПЦ1 и ЛЦ1. На вентральной поверхности каждого из церебральных ганглиев имеется по гигантскому нейрону, расположенному возле корешка внутреннего губного нерва. Это пигментированные клетки размером около 140 мк, легко различимые среди окружающих нейронов меньшего размера. Будучи единственными исследованными идентифицированными нейронами церебральных ганглиев, они могут быть обозначены как клетки ПЦ1 и ЛЦ1. Гистохимическое исследование обнаружило в этих клетках серотонин (см. раздел 4.3.); благодаря этому свойству они представляют для нас специальный интерес, о чём речь пойдет в главе 5. Кандель и Тауц [204] нашли, что потенциал покоя равен примерно 50 мВ; потенциалы действия, имеющие аксонный и соматический компоненты, достигают, по нашим данным, 60- 70 мВ. Клетка молчит или проявляет довольно продолжительные периоды активности. При раздражении нервов наблюдаются крупные ВПСП, а в спонтанном притоке более мелкие ВПСП от двух интернейронов [204]. Клетки получают также тормозной приток от тентакулярных нервов [304]. Ацетилхолин деполяризует, при этом клетки проявляют хорошо выраженную десенситизацию к ацетилхолину. Действие ацетилхолина блокируется кураре. Глутамат имитирует синаптическое торможение [304]. Клеточная мембрана обладает аномальными выпрямляющими свойствами [205]. Следует упомянуть замечательную серию исследований Коттрелла и соавторов [см. 131, 266а], которые нашли, что ПЦ1 и ЛЦ1 являются интернейронами, дающими возбуждающие синаптические окончания на некоторых идентифицированных клетках буккальных ганглиев. Особенно интересно, что аксонные ветви каждой из двух клеток приходят в каждый буккальный ганглий с двух сторон — через правый и левый церебробуккальный коннективы. Всего, как это нашли ещё Кандель и Тауц, каждый из двух нейронов даёт начало трём аксонным ветвям: одна следует в ипсилатеральный наружный губной нерв, другая — в ипсилатеральный церебробуккальный коннектив и третья — в церебральную комиссуру; попав в симметричный ганглий, эта ветвь вступает в контралатеральный церебробуккальный коннектив. Хотя клетки содержат серотонин и выделяют его в своих аксонных окончаниях [см. 131], сами они, по нашим наблюдениям, чувствительны к этому медиатору, который их возбуждает. Деполяризующее действие серотонина быстро исчезает в безнатриевой среде, как и способность генерировать потенциалы действия. Этот эффект обратим.
4. 3. Гистохимическая локализация медиаторных биогеннных аминов
4. 3. 1. Метод формальдегидной конденсации и еговарианты
Единственным доступным и вместе с тем надёжным методом прямого выявления медиаторов в структурах нервной ткани пока что остается люминесцентно-гистохимический метод локализации биогенных аминов, посредством которого выявляются четыре медиаторных амина: дофамин, норадреналин, адреналин и серотонин. Метод основан на получении люминесцирующих производных соответствующих аминов в результате их реакций с формальдегидом. История метода восходит к началу 30-х годов [см. 287], но только в 1962 г. группе шведских авторов удалось разработать чувствительную процедуру, которая впервые позволила локализовать биогенные амины в нервных клетках, волокнах и окончаниях. Эта гистохимическая процедура, обычно называемая методом Фалька и Хилларпа, обеспечила быстрый прогресс в изучении биогенных аминов нервной системы ([см. обзор 129], а также раздел 2.3.2). При всех своих замечательных свойствах метод Фалька-Хилларпа имеет и отрицательные стороны: он капризен, медлителен, трудоёмок. В связи с этим после его появления не прекращались попытки разработать более простую методику на той же или иной химической основе. Мной совместно с А. В. Сахаровой была, в частности, осуществлена разработка простого и быстрого «водного» метода, который как гистохимическая процедура существенно отличается от «газового» метода Фалька-Хилларпа [58, 59]. Нашу методику можно рассматривать, пожалуй, как развитие идеи, лежащей в основе старого метода Эренко для выявления норадреналина в замороженных срезах надпочечника. Мы изучили механизм водного метода и нашли, что в основе его лежат те же реакции, которые обеспечивают люминесценцию биогенных аминов в методе Фалька-Хилларпа [287]; это позволяет рассматривать все названные методы (Эренко, Фалька-Хилларпа и нашу водную методику) как варианты некоего единого по своему химическому механизму метода — метода формальдегидной конденсации. Из сказанного вытекает, что результаты, полученные газовым и водным методами, должны быть одинаковыми, если методы обладают равной чувствительностью. Опыт использования того и другого метода показывает, что это в самом деле так. При локализации биогенных аминов в нервной системе прудовика мы применяли метод Фалька-Хилларпа в оригинальной прописи [286]; с тех пор мне неоднократно приходилось работать с тем же объектом водным методом, и я мог убедиться, что при всем различии процедур результаты реакции одинаковы. Простота водного метода позволила мне накопить большой сравнительный материал по топографии моноаминергических нервных элементов у разных гастропод, и этот материал совершенно сопоставим с литературными данными, полученными на тех же или других гастроподах с помощью газового метода (см. 5.). Водным методом получены и приводимые здесь собственные данные о моноаминергических нейронах виноградной улитки, тогда как почерпнутые из литературы дополнительные сведения взяты у авторов, работавших газовым методом. К сожалению (или, может быть, к счастью), не во всех случаях варианты метода формальдегидной конденсации взаимозаменимы. На мозге теплокровных гистохимическая реакция блокируется при проведении её водным методом, а также некоторыми модификациями газового [287], но реакция идет до конца при применении оригинальной прописи метода Фалька и Хилларпа. Судя по некоторым сообщениям, у газового метода имеются свои ограничения. Так, в работе, один из авторов которой — крупнейший специалист по биогенным аминам беспозвоночных, мы читаем: «Метод Фалька и Хилларпа даёт обычно более успеха при изучении тканей наземных и пресноводных животных, чем когда его применяют на тканях морских животных (например, Aplysia)» [195, стр. 55]. О безуспешных попытках локализовать моноаминергические нейроны в ганглиях ап-лизии методом Фалька и Хилларпа мне известно и из личного сообщения американского нейробиолога Р. Мак-Камана. Между тем, работая водным методом на морских гастроподах, в том числе заднежаберных, мы не испытывали никаких трудностей (неизвестно, впрочем, как поведет себя водный метод с нервной тканью аплизии).
4. 3. 2. Моноаминергические нейроны виноградной улитки
Виноградная улитка оказалась в числе первых объектов, к которым шведские гистохимики приложили метод Фалька и Хилларпа вскоре после его разработки. В то время дело ограничилось публикацией кратких тезисов, а полная работа вышла из печати в 1965 г. [141]. Авторы подтвердили известные к тому времени данные о высоком содержании в нервной системе моллюсков серотонина, обнаружили высокое содержание в ней дофамина и показали, что серотонин и дофамин локализованы в разных нейронах. Каждый из аминов обнаруживается в соматической цитоплазме соответствующей клетки, а также в клеточном отростке, причем наиболее высокая концентрация амина наблюдается в варикозных расширениях аксона в синаптическом нейропиле. Эта работа оказалась важным этапом на пути изучения медиаторной функции серотонина и дофамина в нервной системе моллюсков, но в ней совершенно отсутствовали топографические данные о моноаминергических нейронах и их синаптических терминалях. Топографии моноаминергических элементов было посвящено наше исследование, проведённое совместно с А. В. Сахаровой. Ранее была опубликована лишь небольшая часть полученных данных, касающаяся церебрального ганглия [353], или приводились краткие ссылки на некоторые из результатов [54-56, 279]. Основное внимание было уделено ганглиям ЦНС, в которых локализацию клеток и волокон, проявляющих специфическую люминесценцию, проводили на серийных срезах. Таким же способом были исследованы буккальные ганглии и обе пары щупалец, в которых имеются чувствительные ганглии. Из периферических органов были изучены нога, пенис, мышца — ретрактор пениса (несерийные срезы), сердце (плёночные препараты). Мы нашли, что распределение клеток и волокон, содержащих биогенные моноамины, имеет постоянный характер и закономерно сохраняется от особи к особи; не было отмечено каких-либо различий в этом отношении между двумя изученными видами виноградной улитки. В ЦНС и на периферии имеются нервные элементы, проявляющие зелёное свечение, характерное для катехоламинов, и элементы с жёлтым свечением, характерным для серотонина. Условия реакции, при которых получено зелёное свечение, свидетельствуют о том, что оно обусловлено присутствием первичных катехоламинов. В самом деле, литературные данные по количественному определению биогенных аминов у моллюсков указывают на отсутствие адреналина в ЦНС виноградной улитки и других близких форм; первичные катехоламины представлены в основном дофамином, хотя имеются сведения о присутствии также и норадреналина [209, 260, 334]. Из исследованных отделов центральной нервной системы наиболее богаты биогенными аминами педальные ганглии и педальные доли церебральных ганглиев. Здесь расположены группы преимущественно мелких катехоламиновых нейронов; синаптический нейропиль этих областей проявляет исключительно яркое зелёное свечение. Нейроны этого химического типа закономерно встречаются также в букальных ганглиях и в ганглиях нижней пары щупалец. Клетки с жёлтым свечением найдены в церебральных и педальных ганглиях. Наконец, в ганглиях висцеральной дуги, относительно бедных биогенными аминами, имеется группа нейронов, дающих жёлтое свечение; утверждают, что у садовой улитки в соответствующих клетках содержатся два амина — дофамин и серотонин ([213]; см. также 6.7.). Остановимся подробнее на топографии моноаминергических элементов. Как принято в литературе, мы будем называть «зелёными» клетки и волокна, содержащие первичный катехоламин (т. е. дофаминергические, но возможно, и норадренергические), и «жёлтыми» — нейроны, дающие свечение индоловой природы (здесь серотонинергические). В церебральных ганглиях, которые более или менее симметричны, распределение свечения тоже имеет более или менее симметричный характер. Относительно крупные зелёные нейроны имеются только в плевральной доле каждого из ганглиев, в её дорзальной клеточной коре. Таких клеток всего несколько, они расположены более или менее компактной группой, причём одна из зелёных клеток лежит рядом с гигантским дорзальным нейроном. Аксоны этих клеток, войдя в нейропиль, образуют зоны крупных синаптических контактов. Создаётся впечатление, что эти аксоны дают ветви, нисходящие по церебро-плевральным коннективам в подглоточный комплекс ганглиев. В общем крупные зелёные нейроны составляют очень небольшую часть нейронов, покрывающих дорзальную поверхность церебральных ганглиев. Гораздо более многочисленны клетки этого типа на вентральной стороне, где они составляют несколько компактных групп. Все эти группы мелких зелёных нейронов расположены во внутренних слоях клеточной коры, непосредственно у её границы с нейропилем, в который они посылают пучки своих аксонов. Эти клетки преимущественно находятся на границе между долями, в тех местах, где клеточная кора как бы вклинивается в волокнистое вещество ганглия. Одна из таких групп лежит на границе между плевральной долей и процеребрумом; отходящие от неё зелёные волокна проходят по дорзальной поверхности процеребрума и входят в состав тентакулярного нерва. В нерве зелёные волокна собраны в пучок, который, войдя в тентакулярный ганглий, даёт на своём пути зоны яркого синаптического нейропиля. В дистальной части тентакулярного ганглия пучок распадается, участки зелёного нейропиля отмечаются во всех пальчатых выростах ганглия. Зелёные волокна прослеживаются на внутренней поверхности мышечной стенки щупальца, но в мышце — ретракторе щупальца таких волокон нет. Другие группы мелких зелёных нейронов лежат у границы педальной доли с соседними долями церебрального ганглия — с плевральной и коммиссуральной. Крупное скопление таких клеток имеется в вентральной коре ганглия, где эти нейроны образуют свиту у гигантского вентрального нейрона; сам гигантский нейрон всегда проявляет жёлтое свечение. Указанные группы мелких зелёных клеток являются источником исключительно яркого свечения, характерного для задней половины нейропиля педальной доли. В отличие от метацеребрума, в процеребруме очень мало волокон с зелёным свечением и совсем нет клеток этого типа. Группа тонких зелёных волоконец прослеживается на границе между нейропилем и массой мелких, лишённых свечения нейронов, характерных для этого отдела ЦНС. Указанные волоконца посылают веточки в клеточную массу, где контактируют с телами мелких нейронов. Другие веточки дают редкую сеть синаптических варикозных расширений в нейропиле [рис. 14 в работе 353]. Результаты люминесцентно-гистохимического изучения процеребрума были подкреплены электронно-микроскопическими данными. В частности, было показано, что нервные окончания, содержащие характерные для катехоламинов гранулы с плотным ядром, действительно образуют аксо-соматические контакты [352, 353]. В мезоцеребруме также нет клеток со специфическим свечением. К этому отделу церебрального ганглия подходит пучок зелёных волокон, окружающих густым сплетением начальную часть пучка волокон мезоцеребральных клеток. Жёлтое свечение клеток и волокон, как уже сказано, обусловлено присутствием серотонина, который содержится в нервной системе улитки в значительных количествах [см. ссылки в 132, 166]. Клетки с жёлтым свечением, как и зелёные, имеются только в метацеребральной части каждого из церебральных ганглиев. Большое внимание многих исследователей привлекают содержащие серотонин парные гигантские нейроны, лежащие на вентральной поверхности ганглиев, — клетки ПЦ1 и ЛЦ1. Применение микрохимических методов и электронно-микроскопической гистохимии позволило установить, что желтое свечение этих клеток действительно обусловлено присутствием серотонина. Показано, что эти клетки, в отличие от других исследованных нейронов, способны синтезировать серотонин из 5-окситриптофана и что содержание в них серотонина снижается после перерезки тентакулярного нерва — источника тормозного синаптического притока к этим нейронам [137, 259, 263]. Кроме пары гигантских клеток, в церебральных ганглиях имеются скопления жёлтых клеток среднего размера. Они расположены в педальных долях ганглиев и их волокна образуют скопление варикозных пресинаптических расширений в прилегающей области метацеребрального нейропиля. В педальных ганглиях элементы, проявляющие специфическое свечение, расположены также в основном симметрично. Немногие исключения из этого правила будут упомянуты. Остановимся сначала на клетках, содержащих катехоламины. Для вентральной клеточной коры каждого из ганглиев характерно присутствие мелких зелёных нейронов. Они располагаются во внутренней части коры, у границы с нейропилем, и закономерно находятся в тех местах, где из ганглия выходят педальные нервы, иннервирующие ногу улитки. Отдельные мелкие зелёные клетки иногда видны в самих педальных нервах. Нейропиль вентральной части педальных ганглиев по яркости свечения и обилию зелёных волокон сравним с нейропилем педальной доли церебрального ганглия. Дорзальнее этой области нейропиль расслаивается на тяжи волокон, богатые зелёным свечением, и зоны, свободные от него. В частности, на уровне передней (дорзальной) педальной комиссуры нейропиль разделён на медиальную и латеральную области, причем первая гораздо беднее зелёными волокнами, чем вторая, но и в медиальной области, вблизи комиссуры, имеются участки яркого зелёного нейропиля: На этом уровне в каждом ганглии обнаруживается одна относительно крупная зелёная клетка, лежащая рядом с нейропилем немного нейтральнее комиссуры. Пара ещё более крупных одиночных зелёных клеток расположена непосредственно на уровне комиссуры. Кроме того, в каждом ганглии здесь имеется группа мелких зелёных клеток, которые лежат не компактно, а вперемежку с более крупными нейронами, лишёнными специфического свечения. Эта группа простирается от нейропиля до поверхности клеточной коры. В самой дорзальной части педальных ганглиев имеются четко локализованные небольшие островки очень яркого зелёного нейропиля. Здесь в правом ганглии у его переднего конца отмечена одна непарная относительно крупная зелёная клетка. Обращает на себя внимание большое топографическое сходство в распределении нервных элементов, содержащих катехоламины, в педальных и церебральных ганглиях. Для вентральной части тех и других ганглиев характерно присутствие групп мелких зелёных нейронов у внутренней границы клеточной коры, а также сплошной зелёный нейропиль, для дорзальных областей — присутствие разрозненных относительно крупных зелёных нейронов в разных, в том числе и поверхностных участках клеточной коры, и лишь островки зелёного нейропиля. Маловероятно, что это просто случайное совпадение. Жёлтые клетки педального ганглия, как правило, имеют средние размеры. Они собраны в группы, занимающие, во-первых, поверхность вентральной клеточной коры (в её медиальных и задних участках), а также расположенные у выхода церебро-педальных коннективов. Клетки с жёлтым свечением наблюдаются также в самом церебро-педальном коннективе (только в правом). Помимо этого, в педальных ганглиях имеются небольшие группы мелких жёлтых нейронов. В ганглиях висцеральной дуги, как уже было отмечено, имеется группа клеток, проявляющих жёлтое свечение. У изученных нами видов в состав этой группы входят нейроны преимущественно небольших размеров, расположенные в пограничной области висцерального и правого париетального ганглиев. Топография этой группы несколько изменчива, обычно большая часть её расположена в правом париетальном ганглии, где нейронами этого типа окружены пучки волокон, идущие в оба правых мантийных нерва. Гигантские нейроны, которых много в этой области, как будто не входят в состав рассматриваемой группы клеток. Зелёные волокна, наблюдающиеся в нейропиле ганглиев висцеральной дуги, по-видимому, приходят сюда через плевральные ганглии. Хотя в нервах, отходящих от висцеральной дуги, имеется немало зелёных волокон, неясно, можно ли их считать отростками зелёных клеток, находящихся на периферии. Во всяком случае, опыты с наложением лигатуры на висцеральный нерв показали, что материал, проявляющий свечение накапливается проксимальнее лигатуры,- следовательно, в этом нерве волокна идут из центра на периферию, а не наоборот [261], и, следовательно, их источником служат клетки, лежащие за пределами висцеральной дуги. Участки зелёного нейропиля находятся преимущественно в тех местах, откуда начинаются нервы. Особенно много зелёных варикозных волокон у выхода анального нерва и в его начальном участке. Беднее всех специфическим свечением нейропиль плевральных ганглиев, где имеется небольшое число транзитных зелёных волокон. В буккальных ганглиях нами найдена симметричная парная группа зелёных клеток, расположенных в виде свиты рядом с одной из гигантских клеток в латеральной области каждого из ганглиев. В нейропиле много зелёных и жёлтых волокон. Зелёные волокна в нижней паре щупалец в общем напоминают по своей локализации то, что выше описано для верхней пары (омматофоров). Однако в составе чувствительного ганглия нижних щупалец мы нашли компактную группу мелких зелёных нейронов, тогда как в верхних щупальцах зелёных клеток никогда не наблюдали. В сердце чётко выявляются волокна со специфическим свечением, имеющим как будто жёлтый оттенок. Эти волокна, войдя в сердце, ветвятся и ветви их следуют вдоль сократимых элементов миокарда (как в предсердиях, так и в желудочке). Наши наблюдения, полученные на растянутых тотальных препаратах с помощью водного формальдегидного метода, соответствуют данным авторов, которые смотрели срезы и растянутые препараты сердца виноградной улитки, обработанные формальдегид-газом по Фальку и Хилларпу [110, 133]. Очень богато представлены зелёные нервные волокна в ноге и её дериватах — мышечной стенке пениса и в особой мышце, втягивающей этот орган (ретрактор пениса). Все три мышечных органа пронизаны зелёными волокнами; кроме того, имеются небольшие ярко светящиеся островки типа синаптического нейропиля, расположенные рядом с группами мелких нервных клеток, лишённых специфического свечения. Такие синаптические зоны особенно закономерно представлены в ноге и ретракторе пениса. Сравнимые наблюдения сделаны на этих мышечных органах хелицид авторами, применявшими газовый метод Фалька [100, 275а]. Напротив, другая ретракторная мышца головной части улитки — ретрактор омматофора — совершенно лишена нервных элементов, проявляющих специфическое свечение. Как уже было упомянуто, в омматофоре зелёные нервные волокна наблюдались нами только в наружной мышечной стенке, но не в ретракторе.
4. 4. Электронно-микроскопическое исследование
4. 4. 1. История вопроса
Выше уже отмечалось, что на ультраструктурном уровне химическая специфичность нервных клеток наиболее демонстративно выражается различиями в строении и внешнем виде секреторных органелл. Появившаяся в 1963 г. статья Гершенфельда, посвящённая ультраструктуре синаптического нейропиля некоторых наземных пульмонат [164], была первой публикацией, наложившей определённый отпечаток на последующие работы других авторов. Исследуя нейропиль улиток, Гершенфельд впервые столкнулся с трудностью, известной и в нейрогистологии позвоночных: на основании каких критериев считать контакты синаптическими? К началу 60-х годов электронные микроскописты, работавшие на позвоночных, выработали так называемую «классическую триаду» критериев: 1) наличие секреторных везикул в пре-синаптическом окончании; 2) скопление митохондрий там же; 3) уплотнение пре- и постсинаптической мембраны или одной из них в области активного контакта. Оказалось, что работая с гастроподами, опираться на эту триаду признаков весьма непросто. Синаптический нейропиль в ганглиях гастропод представляет собой массу переплетающихся и контактирующих нервных волокон, почти не разделённых глией. Это вслед за Гершенфельдом обнаружили и другие авторы, работавшие с другими видами. Вот цитата из нашего с В. Л. Боровягиным электронно-микроскопического атласа нервной ткани тритонии [4]: «Нейропиль… на гистологических препаратах имеет вид плотного беспорядочного войлока, в который вкраплены отдельные, немногочисленные в сравнении с кортикальной зоной, ядра глии. На первой стадии ознакомления с электронно-микроскопическим материалом впечатление хаоса не ослабевает, а напротив ещё более усиливается. Волокна разного диаметра, идущие пучками и порознь, а иногда одно внутри другого; соприкосновения этих волокон, слишком частые для того, чтобы в каждом случае предполагать синаптические контакты; скопления различных, чаще всего разнородных везикул, нередко лежащие в каждом из двух соприкасающихся волокон — всё это создает впечатление, что сколько-нибудь обоснованная функциональная трактовка наблюдаемых картин невозможна» [стр. 22]. В этих словах определённо выражена растерянность перед трудно трактуемым материалом. Можно думать, что Гершенфельд тоже должен был её испытывать. Характерен пример с уплотнением мембран. Сейчас достаточно хорошо известно, что и у позвоночных далеко не все химические синапсы характеризуются этим признаком [см., например, 91], у беспозвоночных же он представлен лишь в редких случаях. «Уплотнения», продемонстрированные Гершенфельдом, скорее всего — случайные колебания плотности или артефакты, что хорошо видно при внимательном рассмотрении фотографий. С такой же предвзятостью был трактован Гершенфельдом вопрос о секреторных органеллах. Гершенфельд утверждал, что в нервных окончаниях имеется только три вида таких органелл: 1) прозрачные пузырьки диаметром 600 — 800 Å, 2) пузырьки диаметром 800 — 1100 Å, имеющие плотное зерно и 3) структуры типа элементарных нейросекреторных гранул диаметром 1200 — 1400 Å. В одних окончаниях, по словам Гершенфельда, имеются пузырьки только первого типа, в других они смешаны с пузырьками второго или третьего типа. На самом деле, как будет показано в этой главе, секреторные пузырьки лёгочных улиток относятся к большому разнообразию хорошо различимых типов. Классификация, сделанная Гершенфельдом, была навязана знаниями об ультраструктуре нервной системы позвоночных. Нужно признать, что оценка, данная В. Л. Боровягиным и мной нейропилю тритонии, была немногим лучше. Мы писали в своём атласе: «Здесь присутствуют электронноплотные, ограниченные мембраной крупные гранулы диаметром около 1000 — 1500 Å,… прозрачные пузырьки диаметром до 500 — 600 Å и пузырьки с плотным зерном, имеющим варьирующую плотность» [4, стр. 24]. Это утверждалось, несмотря на то, что на опубликованных нами фотографиях чётко видно, что разнообразие органелл необычайно велико и что, в частности, крупные гранулы различны в разных волокнах. Последующие годы позволили во многом разобраться, решить ряд трудных вопросов и в значительной степени освободиться от указанного психологического препятствия — стремления подогнать данные под стандарт. Упомянем наиболее важные вопросы, подвергшиеся анализу. Вопрос о месте синаптических контактов в ЦНС гастропод сначала решался с излишней категоричностью, когда в ряде работ было сделано заключение, что единственным местом контактов является синаптический нейропиль, тогда как аксо-соматических синапсов у гастропод не бывает. Вопрос этот с самого начала получил нездоровое развитие в связи с опубликованием А. Абрахамом статей о «перицеллюлярных синапсах», у аплизии [например, 72]. Профессор Абрахам любезно предоставил мне возможность ознакомиться с некоторыми своими препаратами, и у меня сложилось впечатление, что примененный им метод импрегнации выкрашивает межуточное вещество оболочки ганглия, в связи с которым и находится сеточка, окружающая тела нейронов. Категоричность выводов Абрахама, известного нейрогистолога, вызвала между тем противоположную реакцию у электронных микроскопистов, которые на том же объекте не смогли найти вокруг тел нейронов ничего, кроме сателлитной глии. Точно так же, только сателлитную глию, находили в окружении тел нейронов авторы, исследовавшие лёгочных и голожаберных моллюсков, в том числе тритонию (см. библиографию в [4]). Однако изучение процеребральной области церебральных ганглиев, проведённое нами совместно с И. Ж.-Надем на двух видах пульмонат, показало, что отсутствие аксо-соматических синапсов не является общим правилом: для этой области ЦНС такие синапсы весьма характерны ([352, 353]; см. также раздел 4.4.3.). Аналогичные картины были найдены затем в педальных ганглиях прудовика [35]. Можно считать решённым вопрос об особом способе контакта «волокно в волокне». Предположение некоторых авторов, что такие контакты представляют собой род синапсов, было проверено нами с В. Л. Боровягиным на тритонии и не получило подтверждения: внутриаксонные шнуры оказались выростами глиальных клеток и им была приписана трофическая функция [4]. Большим шагом вперёд явилось электронно-микроскопическое исследование отдельных идентифицируемых клеток с известными электрофизиологическими характеристиками. Такую работу американские авторы осуществили на клетках абдоминального ганглия аплизии [122, 125, 160]. Наиболее важный вывод из неё заключается в том, что каждая клетка или группа однородных клеток обладает характерным для неё набором ультраструктурных признаков. Предпринятое нами (частично в сотрудничестве с И. Ж.-Надем и Н. К. Остроумовой) электронно-микроскопическое изучение нервной системы виноградной улитки имеет такое же направление. Уточним, какие электронно-микроскопические сведения важны для обсуждения вопросов, поставленных в главе 3. Во-первых, как и при изучении нейронов улитки другими методами, желательно получить представление о том, насколько разнороден клеточный состав нервной системы и насколько эта разнородность постоянна. Во-вторых, интересно выяснить, какие типы нейронов представлены особенно богато, а какие — скудно. Такие сведения интересны для сравнения улитки с другими животными. В-третьих, нам важно знать, одинакова ли химическая специфичность окончаний, выполняющих одинаковую функцию. Для этого мы рассмотрим моторные нервные окончания на разных мышцах. Хотя мы располагаем неплохим собственным материалом, для большей полноты картины будут привлекаться литературные данные, что будет оговорено в тексте.
4. 4. 2. Материал
Ввиду большого объёма нервной системы виноградной улитки, мы не могли обследовать её целиком и подвергли изучению восемь произвольно избранных участков. Они показаны на схеме (рис. 10). Участки эти занимают следующее положение: 1. Процеребральный отдел церебрального ганглия. 2. Мезоцеребральный отдел церебрального ганглия. 3. Участок дорзальной поверхности метацеребрального отдела церебрального ганглия. (Нужно добавить, что указанные три участка брались как от правого, так и от левого ганглия; про- и мезоцеребральные отделы фиксировались целиком). 4. Передне-медиальная область левого париетального ганглия. 5. Передне-медиальная область правого париетального ганглия. 6. Задне-латеральная область правого париетального ганглия. 7. Передний конец висцерального ганглия. Участки 4 и 5 фиксировались на всю толщину ганглия, тогда как участки 6 и 7 брались только с дорзальной стороны ганглия. Для участков 2 — 7 трудоёмкая работа по измерению диаметра гранул проведена Н. К. Остроумовой. 8. Тентакулярный ганглий (от правого и левого омматофора), целиком. Нейро-эффекторные окончания изучались на мышечных клетках периневрия (оболочки ганглиев), на мышечных клетках ретрактора глазного щупальца и на железистых клетках («воротничковые клетки») омматофоров. Во всех случаях материал подвергался двойной фиксации (глутаровый альдегид, затем четырёхокись осмия) и контрастировался уранил-ацетатом и (или) лимоннокислым свинцом по общепринятой методике. Данные об иннервации миокарда, мышечных клеток ретрактора пениса и ноги взяты из литературы.
 Рис. 10. Участки церебрального (1, 2, 3), левого париетального (4), правого париетального (5, 6), висцерального (7) и тентакулярного (8) ганглиев, подвергнутые электронно-микроскопическому исследованию (см. текст).
Рис. 10. Участки церебрального (1, 2, 3), левого париетального (4), правого париетального (5, 6), висцерального (7) и тентакулярного (8) ганглиев, подвергнутые электронно-микроскопическому исследованию (см. текст).
4. 4. 3. Клетки и синапсы ЦНС
1. Процеребрум. Процеребральные отделы церебральных ганглиев виноградной улитки давно привлекают внимание исследователей своей необычностью, непохожестью на другие области окологлоточного нервного кольца. Здесь имеются только очень мелкие нервные клетки, что резко отличает процеребрум от всех других ганглиев, где наряду с некоторым числом мелких нейронов представлены клетки средних размеров, крупные и гигантские. Далее, процеребрум отличается тем, что нейропиль в нём занимает боковое положение, тогда как вообще для ганглиев моллюсков характерно центральное положение нейропиля, окружённого со всех сторон телами нейронов. Необычным является также и то, что процеребрум развивается из отдельной закладки и присоединяется к остальной части церебральных ганглиев на относительно поздней стадии онтогенеза. Наконец, нужно отметить, что процеребрум имеется только у стебельчатоглазовых лёгочных улиток; во всех остальных группах класса гастропод такого образования в ЦНС нет, и попытки установить гомологии этого отдела встречаются с известными трудностями. Как показало электронно-микроскопическое исследование, процеребрум необычен и по своей тонкой структуре, что выражается в мощном развитии системы аксо-соматических синапсов. Помимо виноградной улитки, процеребрум был нами изучен у одного из видов слизней (см. 5.3.5.). Результаты этого исследования подробно изложены в статье, хорошо иллюстрированной электронно-микроскопическими фотографиями [353]. Это позволяет нам ограничиться здесь кратким изложением главных результатов. Процеребрум виноградной улитки, как и слизня, состоит из нескольких частей, между которыми проходят чёткие границы (см. рис. 3, Б), что хорошо видно в световой микроскоп. Периферическое положение занимает клеточная масса, состоящая из однородно мелких нейронов. Их отростки, сливаясь в пучки, покидают клеточную массу и входят в нейропиль, расположенный в медиальной части процеребрума. Часть нейропиля, примыкающая к клеточной массе, называется терминальным нейропилем. Четкая граница отделяет его от так называемого внутреннего нейропиля, который простирается медиально к мезоцеребруму. Если в клеточную массу входить со стороны периневрия, то в первую очередь встречаются периферически расположенные в ней крупные «пограничные клетки», не имеющие нейрональных черт и, по-видимому, представляющие собой род питательных клеток. Их многочисленные отростки располагаются между нейронами наружных слоев. Пограничные клетки имеют ряд интересных черт строения (например, крупные митохондрии, содержащие многочисленные электронноплотные гранулы), но здесь нет смысла на этом задерживаться. Нейроны наружных слоев клеточной массы имеют неправильную форму и богаче цитоплазмой, чем лежащие глубже. Мелкие, не более 10 мк в диаметре, нейроны внутренних слоев имеют лишь тонкий ободок цитоплазмы вокруг ядра и упакованы исключительно плотно, так что нередко встречаются прижатые друг к другу клетки, разделённые лишь межклеточной щелью. В других случаях между наружными мембранами двух соседних нейронов приложен узкий однослойный глиальный тяж. Эти прослойки образованы отростками глиальных клеток, весьма немногочисленных в клеточной массе процеребрума и по размерам своего клеточного тела не отличающихся от нейронов. Нейроны униполярны, единственный отросток направляется в сторону нейропиля. Иногда при прохождении отростка около сомы другого нейрона в соме наблюдается скопление пузырьков в области такого контакта. Эти прозрачные пузырьки размером 500 — 800 Å, возможно, свидетельствуют о существовании сомо-аксонных синапсов (таковые описаны у позвоночных). Скопления таких же пузырьков видны в некоторых аксонах, образующих в клеточной массе аксо-аксонные или аксо-соматические контакты. Такие аксоны, как правило, следуют в направлении, поперечном по отношению к пучкам волокон, выходящих из клеточной массы. В пресинаптическом аксоне хорошо различимы узкие участки, в которых имеются только нейротубулы, и расширения, богатые митохондриями и синаптическими пузырьками. Каждое расширение обычно образует контакты гломерулярного типа сразу с несколькими нейронами или их отростками. В пучке тонких, диаметром около 0,5 мк, аксонов, покидающих клеточную массу, обычно можно встретить одно или несколько более толстых волокон, которые содержат везикулы с электронноплотным зерном. Размеры таких везикул — около 800 — 1200 Å. Такие волокна тоже образуют аксо-соматические контакты в клеточной массе. Поскольку в клеточной массе нет клеток, которые могли бы дать начало таким волокнам, мы предположили, что они приходят в процеребрум извне. В самом деле, люминесцентно-гистохимическое исследование показало, что этим аксонам соответствуют волокна, дающие зелёное свечение при выявлении биогенных моноаминов (см. 4.3.2). Как оказалось, небольшой пучок волокон, содержащих первичный катехоламин, вступает в процеребрум со стороны метацеребрума, тянется вдоль внутренней границы клеточной массы и отсылает в неё одиночные волокна, в которых видны варикозные расширения, расположенные между телами нейронов. Эта картина вполне соответствует той, которая наблюдается в электронный микроскоп. Редкая сеть люминесцирующих волокон, имеющих варикозные расширения, видна также в терминальной нейропиле, но во внутреннем нейропиле такие волокна отсутствуют. Соответственно, электронно-микроскопические препараты демонстрируют отсутствие волокон с упомянутыми секреторными везикулами во внутреннем нейропиле, тогда как в терминальном такие волокна встречаются. Вообще картина синаптических контактов терминального нейропиля весьма напоминает то, что было сказано в отношении клеточной массы. Напротив, внутренний нейропиль отличается совершенно иной картиной. Здесь наиболее характерным элементом являются аксоны, которые в своей расширенной части содержат не везикулы, а извитые трубочки, которые лишь немногим шире обычных нейротубул (как известно, нейротубулы извитыми не бывают). Внутренний нейропиль процеребрума отличается от терминального не только в ультраструктурном отношении, но и цитохимически: здесь наблюдается высокая активность дегидрогеназы глюкозо-6-фосфата. 2. Мезоцеребрум. Так называются парные выпуклые доли, расположенные в каждом из церебральных ганглиев впереди от церебральной комиссуры (рис. 3, А). В световой микроскоп видно, что правый мезоцеребрум несколько крупнее левого. Каждый заполнен относительно крупными клетками, лишь в латеральной части имеются нейроны меньшего размера. Ни те, ни другие не дают положительной реакции на биогенные амины, но мелкие нейроны или часть из них проявляют себя как гомори-положительные клетки (собственные данные). Электронно-микроскопически в оболочке, окружающей мезоцеребрум, помимо элементов соединительной ткани, выявляется много мелких клеток. Каждая клетка имеет один длинный отросток, нередко с микротубулами, направленный в сторону ганглия. Они бывают тёмными или светлыми в зависимости от количества рибосом. Клетки и их отростки заполнены множеством секреторных гранул изменчивой электронной плотности. Размеры гранул 500 — 1000 Å, изредка до 1300 Å, средний размер определён как 747 Å. Эти клетки идентифицированы как клетки «дорзальных тел» — эндокринных образований, контролирующих активность гонады у гастропод [см. 200]. Ранее дорзальные тела стебельчатоглазых улиток были известны только по данным световой микроскопии. Наши ультраструктурные данные указывают на возможную нейронную природу указанных клеток. Кроме них, в оболочке встречаются нервные волокна, контактирующие с мышечными клетками периневрия. Эти моторные окончания описаны в следующем разделе. В нервной ткани мезоцеребрума нами отмечены три типа нейронов, различающиеся по характеру секреторных гранул. Для цитоплазмы первого из указанных типов характерно относительно небольшое число гранул, их диаметр достигает 3000 Å (средний 2172 Å), гранулы наполнены гомогенным плотным содержимым почти до своей наружной мембраны. Другой тип характеризуется гранулами диаметром до 2000 Å (среднее для 139 гранул — 1366 Å), мелкозернистое содержимое гранул отделено от мембраны светлой зоной. В третьем случае наружный вид гранул такой же, но их диаметр несколько меньше (максимальный 1700, средний для 165 гранул — 1070 Å). 3. Дорзальный метацеребрум. В оболочке этой зоны церебрального ганглия видны уже упомянутые выше мелкие клетки«дорзальных тел». Внешний вид и кривая распределения ихдиаметров такие же, как в мезоцеребруме, средний для 557 гранул диаметр определён в 710 Å. Секреторные гранулы в нервных клетках характеризуются гомогенным содержимым, заполняющим гранулы практически до самой оболочки. Гранулы в перикарионах немногочисленны. По диаметрам гранул можно различить две категории клеток: максимальные размеры гранул в них составляют 1700 и 2300 Å, средний диаметр соответственно 1058 и 1890 Å. 4. Левый париетальный ганглий. В исследованной области ганглия самой крупной является клетка ЛПа3. В её цитоплазме отсутствуют секреторные гранулы, но характерно присутствие большого числа цитосом. Гранулы, однако, в большом числесодержатся в неидентифицированных нейронах, занимающих визученном участке медиальное и вентральное положение. В небольших нейронах, которых много на вентральной стороне, не густо расположены гранулы умеренной плотности, со светлой наружной зоной. Их диаметр до 2000 Å, в среднем для 210 гранул — 977 Å. Имеется также крупный нейрон, содержащий множество гранул с ровным, не очень плотным содержимым, сплошь заполняющим всю гранулу. Диаметр достигает 3000 Å, среднее для 375 гранул 1460 Å. Ещё в одной крупной клетке также наблюдается множество гранул, они тоже лишены наружной светлой зоны, а содержимое отличается гомогенностью и малой электронной плотностью. Эти гранулы самые крупные, максимальный диаметр 3300, средний для 292 гранул — 1923 Å. 5. Правый париетальный ганглий, передняя часть. Гигантская клетка ППа3, как и ЛПа3, гранул не содержит. Небольшие нейроны содержат довольно много гранул диаметром до 2000 Å. Хотя как максимальный, так и средний размер этих гранул (991 Å) близок к величинам, указанным для мелких нейронов левого париетального ганглия, внешний вид гранул здесь совсем иной: они практически не имеют периферической светлой зоны. 6. Правый париетальный ганглий, латеральная область. Мы изучили область, в которой расположена большая группа крупных клеток, обозначаемых нами как группа D и подробно изученных электрофизиологически. Эти клетки содержат множество гранул, обладающих наиболее крупными размерами — до 4000 Å. Гранулы имеют неравномерное плотное, чётко ограниченное центральное ядро и периферическую светлую зону. Средний для 436 гранул диаметр определен в 2063 Å. Для этих нейронов характерна также концентрическая организация эргастоплазмы, образующей фигуры субстанции Ниссля. В изученной области ганглия имеются клетки с гранулами других типов. Группа довольно крупных нейронов характеризуется тем, что в их цитоплазме обширные области заполнены муциноподобным содержимым; между этими пустотами расположены участки активной цитоплазмы, содержащие митохондрии, богатый эндоплазматический ретикулум. Секреторные гранулы расположеныв этих нейронах скоплениями, обычно вблизи крупных липидных включений. Размеры гранул несколько меньше, чем в клетках групп D, — средний диаметр около 1500 Å; содержимое гранул плотное, светлая зона под ограничивающей мембраной невелика или отсутствует. Далее, имеются относительно крупные нейроны, содержащие необычные каплевидные включения сложной ультраструктуры, а также небольшое число гранул с плотным ядром (максимальный диаметр 2000, средний 1350 Å). Ещё один тип — редкие мелкие нейроны с маленькими, в среднем около 700 Å, редкими гранулами, имеющими очень плотное ядро. 7. Висцеральный ганглий. В исследованной области (верхушка ганглия) наблюдается очень разнородный состав нейронной популяции. Изучение не было исчерпывающим, и указываемые ниже три типа нейронов не выражают всего наблюдающегося здесь разнообразия. Прежде всего отметим клетки с большим числом гранул, в которых плотное содержимое не оставляет наружной светлой зоны. Максимальный диаметр 2000, средний для 560 гранул — 1100 Å. Цитоплазма этих клеток отличается богатством нейрофиламентов, имеются и нейротубулы. Эти структуры (филаменты) отсутствуют в клетках другого типа, в которых гранулы по внешнему виду похожи на описанные выше. Но диаметр гранул здесь иной: максимальный — 3000, средний для 170 гранул — 1380 Å. Гранулы здесь расположены реже. Наконец, имеются богатые рибосомами небольшие нейроны с гранулами, плотное зерно которых часто расположено эксцентрично. Диаметр до 2300 Å, средний для 217 гранул — 1293Å. 8. Тентакулярный ганглий. Расположенный в концевой части омматофора (верхнего щупальца) тентакулярный ганглий состоит из двух частей: округлого проксимального основания и дистальной части, имеющей форму пальцевидных выростов. Эти выросты вытянуты в сторону чувствительного обонятельного эпителия, покрывающего концы щупалец. В каждом таком выросте имеются мелкие нейроны, пучки волокон и участки синаптического нейропиля. Проксимальная часть ганглия представляет собой как бы расширение па конце тентакулярного нерва, приходящего из церебрального ганглия. Центральную зону этой части занимают пучки волокон, продолжающиеся в тентакулярный нерв, а на периферии находятся тела нервных клеток, в основном мелких. Среди волокон бросается в глаза масса очень тонких аксонов, часто не разделённых глией, которые, по-видимому, являются нисходящими отростками клеток тентакулярного ганглия и восходящими отростками мелких нейронов процеребрума [см. об этом 176]. Кроме этих тонкихаксонов, здесь, как и в самом тентакулярном нерве, виден пучок толстых нервных волокон, которые транзитом проходят через тентакулярный ганглий и, выходя из него, образуют короткий нервик, тут же входящий в ретракторную мышцу омматофора. Это — моторные волокна, об их окончаниях на мышце см. в следующем разделе (4.4.4). Клеточные тела этих мотонейронов расположены в метацеребральном отделе церебрального ганглия [176]. Наконец, в тентакулярный ганглий приходят из церебрального ганглия аксоны катехоламиновых нейронов метацеребрума. Как отмечалось в разделе 4.3.2., эти волокна в виде небольшого пучка вступают в ганглий; проходя через проксимальную часть, они дают веточки в синаптический нейропиль; особенно сильное ветвление этих волокон наблюдается в нейропиле пальцевидных выростов. Волокна, идентифицированные в люминесцентный микроскоп как содержащие первичный катехоламин, имеют такую же ультраструктуру, как соответствующие волокна в процеребруме (см. выше). Они здесь также заполнены гранулами с электронноплотным центральным зерном. Помимо этих гранул, местами встречаются скопления мелких пустых пузырьков, что обычно рассматривается как свидетельство активной секреции. Размеры гранул, как и в процеребруме, составляют примерно 800 — 1200 Å, средний диаметр для 120 гранул определен в 971 Å. В составе тентакулярного ганглия имеется много клеток, цитоплазма которых богата гранулами с иными ультраструктурными характеристиками. Поскольку волокна с такими гранулами не наблюдались в тентакулярном нерве, можно было думать, что эти клетки являются нейросекреторными либо принимают участие в местных нервных связях в пределах щупальца. В самом деле, нередко наблюдаются выходы волокон, богатых гранулами, под оболочку ганглия, где бывают видны картины активной секреции. Из литературы хорошо известно, что у гастропод оболочка ганглиев и нервов, имеющая хорошую связь с сосудистой системой, часто выступает в роли ней рогемального органа, т.е. места запасания и секреции нейрогормонов [например, 336]. Очевидно, это явление представлено и в тентакулярном ганглии виноградной улитки. Мы пока не можем оценить всего разнообразия клеток тентакулярного ганглия, богатых секреторными гранулами; впечатление о гетерогенности популяции таких клеток (составляющих вместе с тем лишь небольшую часть общей клеточной популяции) можно иллюстрировать анализом, проведенным для нескольких нейронов. Первый тип. Гранулы с плотным содержимым, иногда структурированным; содержимое заполняет большую часть объема гранулы, но под наружной её мембраной остаётся светлая зона, от которой иногда отпочковываются мелкие пузырьки. Размеры гранул 1000 — 2200 Å, в среднем 1562 Å. Наблюдаются выходы волокон с такими гранулами к оболочке ганглия и секреция в межуточное вещество. Второй тип. Гранулы с содержимым от небольшой до умеренной плотности, заполненные им обычно до лимитирующей мембраны (периферическая светлая зона, если она есть, очень узкая). Размеры гранул сильно варьируют и достигают иногда 3000 Å, средний диаметр 1750 Å. Наблюдается выход небольших пучков таких волокон в оболочку ганглия, место дальнейшего назначения этих нервиков неизвестно. Для таких волокон характерно присутствие большого числа нейротубул. Клетка с такими гранулами, имеющая довольно крупные размеры, проявляет признаки соматической секреции: её короткие соматические отростки образуют множественные выходы к полости (сосуд?), расположенной внутри ганглия. Третий тип. Гранулы не вполне шаровидные, иногда с неровным контуром, их плотное содержимое занимает относительно небольшую часть объема, оставляя значительную светлую зону, и нередко расположено эксцентрично. Клетки этого типа небольшие, их цитоплазма богата рибосомами и элементами аппарата Гольджи. Размеры гранул в этих клетках около 1300 Å. Мелкие нервные клетки, группы которых расположены в пальцевидных выростах ганглия в непосредственной близости от чувствительного эпителия верхушки щупальца, представляют собой биполярные сензорные нейроны [176]. Богатые митохондриями и нейротубулами дистальные отростки этих нейронов ветвятся в слое специализированных эпителиальных клеток, наружная поверхность которых представляет собой сложную сеть микроворсинок. Ни в этих чувствительных отростках, ни в перикарионах сензорных нейронов секреторных органелл я не наблюдал. Детальнее с цитологией этих биполярных клеток можно ознакомиться по работе Роджерса, изучившего их у садовой улитки [277]; по мнению Роджерса, они воспринимают тактильное раздражение, но серьёзными аргументами это мнение пока не подкреплено, и многие считают, что верхушка омматофора специализирована для хеморецепции.
4. 4. 4. Нейро-эффекторные окончания
1. Окончания на мышечных клетках оболочки ганглия. У виноградной улитки, как у многих гастропод, оболочка, окружающая ганглии и нервы, содержит мышечные клетки. Нам известны два предположения о назначении этих клеток: одни авторы предполагают, что с их помощью устанавливается соответствие между длиной растяжимого тела моллюска и длиной нервов; другие считают, что мышечные клетки способствуют движению гемолимфы через сосуды, которыми насыщена оболочка ганглиев и нервов. Легко наблюдать рефлекторные сокращения оболочки при раздражениях, наносимых на отдаленные участки нервной системы. Эти рефлекторные сокращения свидетельствуют о наличии специальных нервных элементов, обеспечивающих возбуждение мышечных клеток. Электронно-микроскопически мышечные клетки обнаруживались нами в оболочке самых разных отделов нервной системы улитки. Специальные поиски нервных окончаний были предприняты нами совместно с Н. К. Остроумовой на срезах, полученных с оболочки церебрального ганглия, в области мезо- и метацеребрума. Во многих случаях были найдены контакты между мышечной клеткой и одиночным нервным волокном, которое чаще всего лежало вдоль мышечного и соприкасалось с ним на большом протяжении. Никогда не отмечалось, чтобы мышечное волокно получало более одного нервного окончания. В ультраструктурном отношении все нервные волокна, иннервирующие эти мышечные клетки, относятся к одному типу: они содержат секреторные гранулы диаметром 1000 — 2000 Å, в среднем 1437 Å (среднее для 367 гранул). Гранулы имеют электронноплотную сердцевину и периферическую светлую полоску. Такие ультраструктурные признаки в общем характерны для аксонных терминалей пептидергических нейронов. Неизвестно, где находятся тела мотонейронов, дающие начало этим волокнам. Нет также сведений о физиологии синаптической передачи в таких моторных окончаниях. Роджерс, исследовавший мышечные клетки оболочки ганглия у садовой улитки, отмечает в своей статье, что эти клетки иннервируются, но не даёт описания везикулярного содержимого моторных аксонов; на одной из его фотографий, однако, видно, что эти нервные волокна содержат большое число гранул диаметром более 1000 Å, имеющих плотное или умеренно плотное содержимое [276]. 2. Окончания на мышечных клетках ретрактора омматофора. Как описано выше, моторные нервные волокна приходят к ретракторной мышце из метацеребрума в составе тентакулярного нерва. Они проходят сквозь проксимальную часть тентакулярного ганглия и на уровне пальцевидных выростов выходят из ганглия в виде короткого пучка, который тут же входит в мышцу. Электронно-микроскопически я нашел, что, войдя в мышцу, толстые моторные волокна многократно ветвятся и далее пучки относительно тонких ветвей, вместе с окружающей глией, следуют между мышечными волокнами. Имеется несколько форм контактов между такими нервными волокнами и мышечными клетками. В одних случаях от мышечной клетки к нервному волокну направляется саркоплазматический вырост; в таком выросте никогда не бывает миофибрилл. Синаптические пузырьки, о которых речь пойдет ниже, имеются в нервном волокне в области контакта с выростом мышечной клетки, но отсутствуют в неконтактной части. Как правило, к такому содержащему пузырьки участку нервного волокна направлены выросты не одной, а нескольких мышечных клеток. В других случаях наблюдается простой продольный контакт между мышечным и нервным волокнами. Наконец, иногда можно видеть, что нервное волокно как бы погружается, вдавливается в мышечную клетку, окружается саркоплазматической складкой. По своим ультраструктурным характеристикам эти нервные волокна не имеют ничего общего с описанными выше волокнами, иннервирующими мышечные клетки оболочки ганглия. Там крупные электронноплотные гранулы содержатся в аксоне на всем его протяжении и, по всей вероятности, транспортируются к окончаниям из тела мотонейрона. Здесь в ретракторе везикулярные органеллы наблюдаются, как уже сказано, только в пресинаптической части нервного волокна. Непосредственно в области контакта наблюдается порой совершенно чистая популяция прозрачных везикул, средний диаметр которых мной определён равным 470 Å. На некотором отдалении от пресинаптической мембраны в нервном волокне бывают видны, но не всегда, более крупные пузырьки, иногда довольно многочисленные. Внутри этих пузырьков наблюдаются в большем или меньшем числе зернистые включения, а их наружный поперечник составляет около 1000 Å, достигая иногда 1200 Å. Для этих моторных окончаний в литературе также имеются данные Роджерса, изучавшего их на садовой улитке и одном из видов слизней [275]. Особенности иннервации ретрактора омматофора у этих форм до деталей совпадают с тем, что описано выше для виноградной улитки. Везикулы относятся к двум размерным категориям: меньшие, диаметром около 500 Å, лежат у пресинаптической мембраны, другие, диаметром 800 — 1000 Å, — не обязательно вблизи области контакта. Те и другие прозрачны, но это отличие от моих результатов, возможно, объясняется тем, что Роджерс не применял глутаральдегидной префиксации, которая, как известно, сильно сказывается на сохранении содержимого секреторных органелл. 3. Окончания на мышечных клетках стенки омматофора. Стенка омматофора, будучи участком стенки тела, по-видимому, имеет сложный мышечный состав и её моторная иннервация не ограничивается одним типом окончаний. Об этом, в частности, свидетельствуют данные Баррантеса, который в мышечной стенке омматофора у одного из видов слизней нашёл два типа окончаний: в одном секреторные гранулы диаметром от 710до 950 Å содержат центральное ядро умеренной плотности, в другом типе гранулы диаметром 1000 — 1550 Å почти целиком заполнены содержимым высокой электронной плотности; в тех и других окончаниях наблюдается примесь прозрачных пузырьков диаметром 520 — 720 Å [87]. Мною у виноградной улитки эта мышца специально не исследовалась, но при изучении ретрактора иногда встречалась возможность рассматривать и фотографировать также область стенки щупальца. При этом были найдены окончания, соответствующие первому типу из описанных Баррантесом. 4. Окончания на мышечных клетках ретрактора пениса. Имеются две хорошие электронно-микроскопические работы, посвящённые иннервации этой мышцы виноградной улитки [100, 158]. Ультраструктурные данные дополняются в них результатами гистохимического и биохимического анализа. Нервы, проходящие внутри этой мышцы, на самом деле представляют собой род периферического сплетения, они содержат в своём составе нервные клетки и участки синаптическо-го нейропиля. Наблюдаются четыре типа аксонных окончаний, если считать и нервно-мышечные и межнейронные контакты, из чего делается вывод об участии в местных нервных регуляциях четырёх медиаторных механизмов. Соответственно, имеются следующие четыре типа секреторных органелл: 1) прозрачные пузырьки диаметром 500 — 600 Å; 2) гранулы с плотным центральным зерном, наружный диаметр 500 — 1200 Å; 3) крупные гранулы диаметром 1200 — 1400 Å, почти целиком заполненные плотным содержимым; 4) крупные пустые пузырьки диаметром около 1500 Å, иногда содержащие небольшое количество аморфного материала слабой плотности. Второй и третий из указанных типов гранул и соответственно два типа окончаний принимают участие в иннервации мышечных клеток. Волокна и окончания второго типа идентифицированы как моноаминергические. 5. Окончания в мышце ноги. Приведём данные Роджерса, полученные на садовой улитке [275а]. Как и в ретракторе пениса, в ноге имеется нервное сплетение, в состав которого входят нервные клетки. Роджерс сообщает об этом, подтверждая старые данные световой микроскопии, но описания ультраструктуры этих нейронов не дает. Везикулярное содержимое нервных окончаний в сплетении относится к четырём различимым типам: 1) мелкие прозрачные пузырьки (400 — 700 Å); 2) крупные прозрачные пузырьки (1000 — 1500 Å); 3) пузырьки с очень плотным содержимым (1000 Å); 4) пузырьки с менее плотным содержимым (1000 — 2000 Å). Аксоны, иннервирующие мышечные клетки ноги, берут начало в нервном (подошвенном) сплетении, при этом моторные окончания бывают трёх разных типов и содержат, соответственно, пузырьки первого, третьего и четвёртого типов. Роджерс ссылается на физиологические данные, указывающие на существование в мышце ноги не только возбуждающих, но и тормозных моторных окончаний. 6. Окончания в миокарде. Вотношении миокарда виноградной улитки мы не имеем своих электронно-микроскопических результатов, но данные, полученные другими авторами (как ультраструктурные, так и физиологические), свидетельствуют об участии нескольких разных механизмов в экстракардиальной нервной регуляции. Физиологическими методами у виноградной улитки, как и у других исследованных гастропод, выявлены по крайней мере три таких механизма: тормозной холинергический, стимулирующий серотонинергический и ещё один стимулирующий, осуществляемый при посредстве «субстанции X» (см. 6.1., а также [133, 298, 333]). Соответственно у аплизии при внутриклеточном отведении найдено в сердечной мышце два типа возбуждающих постсинаптических потенциалов и один тип тормозных; более того, найдены клетки в ЦНС, дающие начало всем трём типам экстракардиальных волокон [233а, 239]. На виноградной улитке такие исследования пока не проводились. Электронно-микроскопические исследования миокарда виноградной улитки обнаружили, во-первых, множество аксонных терминалей, заполненных электронноплотными гранулами пептидергического типа диаметром 1000 — 2500 Å [134]. Скорее всего, это — неоднородная популяция пептидергических волокон, одни из которых,как аксонные терминали клетки ППа1, могут быть нейросекреторными и не участвовать в иннервации миокарда [162а, б], а другие — моторными. Последние могут содержать «субстанцию X», для которой известно, что это вещество с большим молекулярным весом (см. 6.1.). Терминали с крупными электронноплотными гранулами не являются единственным типом окончаний, хотя они и наиболее заметны [см., например, 217]. Тот факт, что люминесцентная микроскопия обнаруживает в сердце сеть моноаминергических волокон (4.3.2), является указанием на наличие ещё одного типа иннервации. Венгерский электронный микроскопист К. Элекеш находит в сердце виноградной улитки четыре типа моторных окончаний (личное сообщение). 7. Окончания на железистых клетках омматофоров. Из нескольких типов железистых клеток, представленных в щупальцах, мной исследована иннервация так называемых воротничковых клеток (collar cells), большое скопление которых находится в терминальной части щупалец на уровне пальцевидных выростов тентакулярного ганглия. Не обсуждая здесь не решённого до конца вопроса о функции этих клеток, заметим лишь, что имеются веские основания считать их эндокринными железами, контролирующими развитие гонады. Собранные в компактную массу, эти клетки могут рассматриваться как особый железистый орган. Железа богато иннервирована; нередко можно видеть несколько нервных окончаний на одной железистой клетке. Чаще всего нервное окончание бывает довольно сильно погружено в железистую клетку. По характеру секреторных органелл все окончания, найденные на воротничковых клетках, относятся к одному типу. Они содержат популяцию крупных гранул, в добавление к которым в местах активной секреции, как это обычно бывает, наблюдается скопление мелких прозрачных пузырьков. Крупные гранулы заполнены мелкозернистым содержимым, количество которого неодинаково в разных гранулах, отчего изменчива и их плотность, как правило, небольшая. Нередко под лимитирующей мембраной наблюдается светлая кайма, а иногда картина бывает обратной: темный материал под мембраной и светлая центральная часть. Вообще, нужно отметить, что из всех наблюдавшихся типов нервных окончаний окончания на воротничковых клетках выделяются наиболее изменчивым видом содержимого секреторных гранул. Размеры гранул 1000 — 1600 Å, в среднем 1260 Å. Сходный характер, судя по данным Роджерса, имеет иннервация воротничковых клеток у садовой улитки [276а].
4. 5. Заключение
Резюмируем полученные результаты, обращая внимание на те из них, которые важны для дальнейшего обсуждения поставленных вопросов. Отметим прежде всего электрофизиологические данные, свидетельствующие о том, что знак и механизм реакции на медиатор определяется специфическим характером рецепторов воспринимающей клетки. Ни к одному из медиаторных веществ, изученных на нейронах моллюсков, не приложимо название «возбуждающий медиатор» или «тормозной медиатор», каждое из них оказывает и возбуждающие, и тормозные воздействия. Далее, результаты исследования показывают, что клеточная популяция нервной системы улитки весьма разнородна и что её разнородность носит постоянный, строго фиксированный характер. Постоянство клеточной мозаики демонстративно выявляется электрофизиологически. Индивидуальные клетки или группы клеток обнаруживают особенные, только им присущие сочетания свойств — таких, как характеристики собственной электрической активности, своеобразие синаптического притока, т. е. связей с другими нейронами, знак и ионный механизм реакций на медиаторы и т. д. Эти особенные свойства выражены с таким постоянством, что исследователь может картировать клетки с известными характеристиками. В некоторых случаях индивидуальные клетки или группы нейронов можно опознавать визуально, в других они внешне не отличаются от окружающих нейронов, но могут быть идентифицированы в эксперименте. Также и гистохимическое исследование подтверждает важный факт гетерогенности и постоянства клеточной мозаики. Накопленные данные, хотя они и далеки от полноты, дают некоторое представление о том, насколько разнородна нейронная популяция у улитки. Особенно богатую информацию об этом дают электронно-микроскопические исследования. При рассмотрении этих результатов мы можем исходить из утверждённого опытом современной нейробиологии положения, что особенному типу передачи в химических синапсах соответствует особенный ультраструктурный тип секреторных органелл. Полученные нами данные, дополненные литературными, свидетельствуют, таким образом, о том, что нейроны и синапсы улитки относятся к большому числу разных химических типов. Одновременно электронно-микроскопические данные полностью подтверждают факт постоянства нейронной мозаики и картины нервных связей: клетки определенного типа имеют закономерное расположение и иннервация определённых эффекторных клеток всегда осуществляется совершенно определёнными типами нервных окончаний. Конечно, в разных участках нервной системы неизбежно обнаруживаются не только разные, но и одинаковые клетки или волокна. Так, варикозные терминальные волокна, содержащие первичный катехоламин (как это следует из данных гистохимии), обнаруживаются электронно-микроскопически в процеребруме и в тентакулярном ганглии, проявляя в том и другом месте одинаковые ультраструктурные характеристики; возможно, такими же являются волокна, дающие один из типов моторных окончаний в стенке тела, ноге и ретракторе пениса. Но эти единичные факты дублирования не должны заслонить общей картины разнообразия типов химической специфичности. Особенно бросается в глаза широкое распространение и обилие нейронов, содержащих тот или иной тип крупных, предположительно пептидергических гранул. Такие клетки не только богато представлены количественно, но и проявляют большое число разных типов. Это обращает на себя внимание как резкий контраст по сравнению с тем, что известно для млекопитающих. Кроме того, у млекопитающих пептидергические нейроны выполняют нейросекреторные функции, тогда как у улитки мы находим как нейросекреторные, так и синаптические окончания, заполненные теми или иными пептидергическими гранулами. Нельзя не обратить внимания и на то, насколько разнообразны у улитки типы нейро-эффекторных окончаний. В одном мышечном органе (например, сердце, нога) здесь деятельностью мышечных клеток управляют три-четыре типа нервных элементов, различающихся по своему медиаторному химизму, — и эти микроскопические данные находят полное подтверждение в результатах физиологических исследований. При этом остаются полностью неизученными нейро-эффекторные отношения в целых системах органов улитки — в половой, пищеварительной, выделительной и т. д., где медиаторное разнообразие, быть может, окажется ещё более выраженным. Однако даже из имеющегося ограниченного материала видно, что в распределении этого разнообразия эфферентных нервных элементов есть закономерность. В иннервации ретрактора пениса участвуют, судя по описаниям, те же типы нервных волокон, которые представлены в ноге. Смысл этого совпадения станет понятным, если вспомнить, что ретрактор пениса имеет у улитки педальное происхождение, т. е. является дериватом мышцы ноги. Другой изученный ретрактор — ретрактор щупальца происходит из другой, колумеллярной мышцы, и характер иннервации здесь совершенно иной, чем в ноге или ретракторе пениса, хотя оба ретрактора очень сходны по характеру своего функционирования. Значение этих данных будет подробно рассматриваться в главе 6, а пока нужно перейти к главе 5, которая посвящена вопросу о родственных отношениях клеточных структур нервной системы виноградной улитки.
5. КЛЕТОЧНЫЕ ГОМОЛОГИИ В НЕРВНЫХ СИСТЕМАХ ГАСТРОПОД
5. 1. Критерии установления гомологии
Зная хотя бы отчасти состав клеточной мозаики в нервной системе виноградной улитки, мы можем сравнивать его с нейронной популяцией какого-нибудь другого изученного вида,- например, аплизии. Сравнение покажет, что в этих разных нервных системах имеются клетки со сходными наборами свойств. Попытавшись проанализировать природу этого сходства, мы придем к выводу, что оно отнюдь не конвергентного характера: идентичными свойствами обладают нейроны одинакового происхождения — гомологичные нейроны. Мысль о существовании отдалённых гомологий на уровне индивидуальных клеток возникла у автора этих строк в конце 60-х годов, и то, что она подтверждается, казалось совершенно поразительным. Помню, каким большим волнением было в то время думать и писать, что наша ППа1 не просто похожа на R15 аплизии, а это та же самая клетка, только сидящая в другой нервной системе! Но проходит совсем немного лет, и с этим свыкаешься, и уже без эмоций, лишь с удовлетворением читаешь о поразительном (честно-то говоря) факте, что эти две клетки, находящиеся в таких далёких нервных системах, дают идентичную картину белкового синтеза, непохожую на ту, которая характерна для других, соседних нейронов [162, 162а]. Так, думаешь, и должно быть: ведь нейроны гомологичные. Читатель, знакомый с предметом, простит мне эту небольшую экскурсию за пределы делового стиля, потому что он сам в своей работе встречается с похожими вещами. Итак, нас сейчас интересуют критерии гомологии. Вопрос о критериях многократно пересматривался, и в настоящее время большинством эволюционистов приняты критерии гомологии, выдвинутые немецким морфологом Ремане. В отечественной литературе этому вопросу посвящён критический обзор М. С. Гилярова [13], из которого можно почерпнуть необходимые сведения о критериях Ремане. Выполняться должен один из трёх критериев. Критерий положения учитывает позицию структур в их отношении к другим сравнимым структурам. Критерий специального качества становится важным, когда позиции сравниваемых структур различны; при этом сходство должно наблюдаться по нескольким разным качественным признакам. Наконец, критерий непрерывности помогает гомологизировать несходные структуры, если между ними в сравнительном ряду имеются образования, отвечающие первым двум критериям. Прилагая эти критерии к нервным клеткам, нужно учитывать некоторые особенности этих структур. При рассмотрении вопроса о гомологии клеток, относящихся к разным нервным системам, нельзя ограничиваться данными о положении тел нейронов; очень важно располагать также данными о клеточных отростках и их связях. Не исключено, что изменения положения нейронов, связанные с процессами ганглионизации, концентрации ганглиев и т. п., гораздо сильнее сказываются на позициях клеточных тел, чем на положении рабочих отростков. Это можно иллюстрировать рядом примеров, ограничимся ссылкой на собственные данные о катехоламиновых нейронах педального отдела нервной системы гастропод (5.3.4.). Учёт этого обстоятельства позволяет применять критерий положения при гомологизировании нервных клеток, тела которых в ходе эволюции сильно изменили свои позиции. Далее, важно иметь в виду, что специфические нейроны часто меняют свою позицию не изолированно, а вместе с комплексом окружающих нейральных структур: так обычно происходит при концентрации ганглиев. Это обстоятельство также облегчает применение критерия положения. Наконец, важно отметить, что каждый нейрон обладает не какой-то одной особенной чертой, а комплексом специфических характеристик — химических, ультраструктурных и физиологических. Это делает возможным использование критерия специального качества, особенно важного при установлении отдалённых гомологий между нервными клетками. Сочетая разные критерии, можно с большой уверенностью прослеживать ряды гомологичных нервных клеток — от вида к виду, от семейства к семейству и дальше.
5. 2. Сравнительная анатомия о гомологии нервных органов у гастропод
Концепция гомологии широко применялась сравнительной анатомией в отношении нервных органов. На клеточном уровне она практически не могла применяться, потому что знания о качественных различиях между нейронами появились совсем недавно. По этой же причине при установлении гомологии нервных структур до сих пор почти не пользовались критерием специального качества. Критерий положения и критерий непрерывности, напротив, сыграли большую роль при решении вопросов о происхождении и эволюции различных нервных органов, в том числе ганглиев гастропод. Очевидно, что пользование этими критериями резко затрудняется при сравнении животных с разными планами организации — особенно при сравнении разных зоологических типов; по этой причине хорошо аргументированные представления о гомологии нервных органов имеются только для относительно близких форм, тогда как доказывать отдалённые гомологии крайне трудно. Даже внутри класса гастропод нелегко, пользуясь традиционными приёмами сравнительной анатомии, решить вопрос о гомологии некоторых нервных образований. Отсутствие промежуточных форм между Docoglossa и другими группами класса делает, например, невозможным выяснение эволюционной судьбы мантийного ганглиозного кольца. Неясны гомологии левого париетального ганглия лёгочных. Тем не менее имеется немало вопросов, уверенно решённых сравнительной анатомией, и некоторые её выводы будут использоваться нами при рассмотрении вопросов о гомологии клеток. Краткие сведения о системе современных гастропод содержатся ниже, указано также систематическое положение форм, которые в большей или меньшей степени изучаются нейробиологами:
ТИП МОЛЛЮСКИ (MOLLUSCA) КЛАСС БРЮХОНОГИЕ (GASTROPODA) 1. Подкласс Переднежаберные (Prosobranchia) Отряд Diotocardia Acmaea Отряд Monotocardia Viviparus, Ampullaria, Pila, Littorina [300]*, Pomatias , Bithynia [79], Buccinum [141, 300], Neptunea [64, 300] 2. ПодклассЗаднежаберные (Opisthobranchia) Отряд Pleurocoela Aplysia Отряд Pteropoda Clione [7, 49] Отряд Sacoglossa Отряд Acoela Pleurobranchaea, Navanax [232], Glossodoris, Aeolidia [22, 50], Dendronotus [23, 50], Tritonia [4, 8, 37, 144, 278, 282, 331, 347, 347a] 3. ПодклассЛегочные (Pulmonata) Отряд Basommatophora Lymnaea [2a, 11, 35, 200, 286, 288, 290, 336], Helisoma [206], Planorbarius [11, 92, 238] Отряд Stylommatophora Onchidium [255, 256], Succinea, Strophocheilus [175, 195], Achatina, Archachatina[110, 252], Limax[262, 266a, 275, 352, 353], Helix * В квадратных скобках даны ссылки на работы, выполненные на данном моллюске. Для Aplysia и Helix ссылки не даются, так как работ очень много.
По поводу этого нужно, однако, заметить, что система гастропод не устоялась и специалисты-малакологи зачастую не сходятся даже в вопросе о том, как делить этот класс на подклассы. Подробнее об этом можно прочитать в обзоре А. А. Шилейко [68]. Подробно эволюция нервной организации гастропод излагается в зоологических руководствах, в частности, в соответствующем разделе отечественного «Руководства по зоологии», написанном А. В. Ивановым [19]. Богатую информацию содержит глава, посвящённая гастроподам, в фундаментальном руководстве, по нейробиологии беспозвоночных Буллока и Хорриджа [102], дающем и обширную библиографию. Ограничусь небольшими извлечениями, цель которых — рассмотреть в ряду гомологичных органов ганглии виноградной улитки, взятой здесь за исходную форму. Напомню, что виноградная улитка относится к стебельчатоглазым пульмонатам; у всех других стебельчатоглазых имеются те же ганглии, что у виноградной улитки, и различия между разными видами касаются в основном лишь размеров и формы ганглиев, а также отчасти степени их слияния (особенно в висцеральной дуге). Парные церебральные ганглии имеются у всех гастропод, однако, здесь нет, по-видимому, полной гомологии. В частности, процеребральные отделы этих ганглиев, имеющиеся у стебельчатоглазых, развиваются в эмбриогенезе из отдельной закладки и скорее всего представляют собой новообразование, отсутствующее у передне- и заднежаберных. Ван-Моль [321] приводит доводы в пользу гомологии между процеребрумом стебельчатоглазых и латеральной долей церебрального ганглия сидячеглазых пульмонат, но это мнение представляется мне несколько сомнительным хотя бы потому, что у Succinea имеется как латеральная доля, так и процеребрум (правда, Succineidae — группа своеобразная и специализированная [см. 68]). Ещё меньше обоснована точка зрения, что процеребрум представляет собой часть тентакулярного ганглия, сместившуюся по ходу тентакулярного нерва [176]. Лучше всего признать, что вопрос о происхождении и гомологиях этого отдела церебральных ганглиев остается нерешённым. Но и то, что остаётся в церебральных ганглиях виноградной улитки после исключения процеребральных отделов, по-видимому, не вполне гомологично церебральным ганглиям низших гастропод. Скорее, это сборные органы, включившие в свой состав как церебральные, так и некоторые другие, в частности, лабиальные и оптические ганглии низших форм. От церебральных ганглиев три пары коннективов ведут у виноградной улитки к паре педальных ганглиев, паре плевральных ганглиев, на которых висит висцеральная дуга ганглиев, и паре буккальных ганглиев. Педальные ганглии улитки представляют собой центральную часть педального отдела нервной системы. Этот отдел, как и сама нога или её производные, имеется у всех гастропод, но педальные ганглии представлены не у всех форм. У некоторых переднежаберных им соответствуют так называемые педальные стволы — начальный этап ганглионизации педального отдела нервной системы. Подробнее об этом отделе речь пойдет ниже (5.3.4.). Плевральные ганглии представлены у всех гастропод, и принято считать, что они всегда гомологичны. Однако анализ клеточного состава указывает на то, что даже у одного и того же вида правый ганглий может не вполне соответствовать левому [192]. Из трёх ганглиев, лежащих у виноградной улитки на висцеральной дуге, представляется ясной история двух — правого париетального и висцерального (абдоминального). Первый считают гомологом супраинтестинального ганглия переднежаберных, т. е. ганглия, лежащего над кишкой на правом плевро-висцеральном коннективе. Второй произошёл в результате слияния собственно висцерального ганглия с субинтестинальным, т. е. с тем, который у переднежаберных лежит под кишкой на левом плевро-висцеральном коннективе. Вопрос о происхождении левого париетального ганглия улитки, как уже сказано, остаётся неясным. Насколько мне известно, недостаточно изучен вопрос о включении в состав ганглиев висцеральной дуги ряда периферических нервных узлов, представленных у многих гастропод и связанных нервами с висцеральной дугой (например, генитального ганглия, проксимального осфрадиального ганглия, и т. п.). Буккальные ганглии имеются у всех гастропод. От них берёт начало так называемая «симпатическая нервная система». Относящиеся к ней парные гастро-эзофагеальные ганглии могут сливаться с буккальными. Связанные с органами чувств периферические ганглии головного отдела виноградной улитки (такие, как ганглии передней и нижней пары щупалец, ганглии семперовского органа), по-видимому, не имеют гомологичных органов за пределами стебельчатоглазых пульмонат. Эти вступительные замечания позволяют перейти теперь к анализу родственных отношений нейронов виноградной улитки с клетками нервных систем других моллюсков.
5. 3. Установление клеточных гомологий
5. 3. 1. Гомологи метацеребральных клеток ПЦ1 и ЛЦ1
Содержащие серотонин парные гигантские нейроны виноградной улитки, расположенные на вентральной поверхности церебральных ганглиев, описаны в главе 4 и обозначены нами как ПЦ1и ЛЦ1. Примеры установления клеточных гомологий интересно начать с этих нейронов потому, что вопрос об их гомологах поднимался ещё в старой сравнительно-анатомической литературе. Дело в том, что у многих видов, относящихся к отряду стебельчатоглазых, на вентральной стороне церебральных ганглиев имеется пара клеток, выделяющихся крупными размерами, и было вполне естественно предположить, что эти клетки гомологичны [см., например, 102]. Следовательно, для близких форм существование клеток-гомологов признаётся давно. Но, опираясь только на такие признаки, как позиция и размер тела клетки, очень трудно прослеживать ряд гомологичных клеток. В данном случае этот ряд не выходил за пределы отряда. Однако, опираясь на критерий положения, можно установить не только ближние, но и отдалённые гомологии нейронов ПЦ1и ЛЦ1. Для этого нужно учитывать относительные позиции и форму не столько тел нейронов, сколько их отростков, о чём говорилось в разделе 5.1. Для клеток ПЦ1 и ЛЦ1виноградной улитки хорошо известно, куда следует каждая из трёх аксонных ветвей: одна покидает ганглий через наружный губной нерв, другая — через церебро-буккальный коннектив и третья уходит в церебральную комиссуру [204]. Коттрелл, применив инъекцию проционового жёлтого, показал, что клеточный отросток сначала делится на две ветви, из которых одна направляется к церебральной комиссуре, а другая в свою очередь даёт начало двум ветвям, названным выше [131]. Если сравнить эту картину с тем, что известно для парных гигантских церебральных нейронов голожаберного моллюска тритонии (клетки 8 и 19 [по 347]), то получается поразительное совпадение. Как нашла М. С. Манохина, у каждого из двух нейронов отросток сначала делится на две ветви, из которых одна входит в церебральную комиссуру, тогда как другая подходит к основанию нерва С4 и церебро-буккального коннектива и здесь снова двоится, посылая по ветви в каждый из этих стволов [см. 4]. Наконец, электрофизиологические исследования Дорсетта [144] и Уиллоуса [347], сделанные на двух разных видах тритонии, показали наличие связей этих гигантских нейронов с буккальными ганглиями. Каждый из нейронов посылает свои импульсы к буккальным ганглиям через оба церебро-буккальных коннектива — правый и левый. Даже в этой детали пара нейронов тритонии проявляет замечательное сходство с клетками ПЦ1 и ЛЦ1 виноградной улитки. Таким образом, несмотря на огромные различия в анатомии ЦНС виноградной улитки и тритонии, удаётся с большой надёжностью установить гомологию гигантских церебральных клеток, пользуясь критерием положения. Вместе с тем сходным оказывается и медиаторный химизм рассматриваемых нейронов виноградной улитки и тритонии. У виноградной улитки, как уже говорилось, клетки ПЦ1 и ЛЦ1 выделяются среди окружающих нейронов тем, что они содержат серотонин, который выполняет синаптические функции в аксонных терминалях этих нейронов (см. 4.). Точно так же в церебральных отделах церебро-плевральных ганглиев тритонии гигантские парные нейроны, по данным М. С. Манохиной и Л. В. Кузьминой [37], подтверждённым американскими нейрохимиками [331], содержат серотонин. Вывод о гомологии рассматриваемых нейронов виноградной улитки и тритонии я сделал в своё время, не располагая ещё данными о наличии серотонина в парных гигантских церебральных нейронах тритонии, но у меня были такие данные для другого голожаберного слизня — дендронота [54, 279]. Вывод этот означал, что и у других лёгочных, а также у других заднежаберных моллюсков, в церебральных отделах ЦНС можно найти крупные парные нейроны, содержащие серотонин, и что все эти клетки должны относиться к одному клеточному ряду. Данные, накопившиеся за последующее время, свидетельствуют, что это в самом деле так. Гистохимическое исследование показало, что у стебельчатоглазых улиток и слизней, имеющих пару гигантских нейронов в вентральном метацеребруме, эти клетки во всех изученных случаях содержат серотонин. Я сам это видел у Eobania vermiculata, Caucasotachea sp. (Helicidae), Bradybaena fruticum (Bradybaenidae), Limax maximus (Limacidae). Литературные данные имеются для Helix aspersa [292], Limax maximus [262] и для южноамериканской улитки Strophocheilus oblongus, представляющей древнее семейство Strophocheilidae [195]. Ещё раньше гигантскую пару нейронов, содержащих серотонин, мы нашли в церебральных ганглиях прудовика [286]. Вслед за тем английские авторы сообщили, что пара серотонинсодержащих гигантских нейронов имеется у другого представителя сидячеглазых пульмонат — катушки [238]. Недавно на прудовике я провёл дополнительное исследование, чтобы уточнить позицию гигантских клеток, содержащих серотонин. Оказалось, что они расположены на латеральных склонах передней доли каждого из церебральных ганглиев. При этом в правом ганглии кпереди от серотонинсодержащего нейрона лежит однородная группа нейронов среднего размера; в левом ганглии эта группа слабо представлена или отсутствует, так что гигантский нейрон занимает, как у тритонии, переднее положение.
 Рис. 11. Парные серотонинсодержащие гигантские нейроны в церебральных отделах нервной системы у представителей разных групп гастропод.
А — виноградная улитка; Б — слизень Limax; В — прудовик; Г — тритония. Парные церебральные ганглии или их производные (у тритонии — цереброплевральные ганглии) изображены с вентральной стороны, у одного из ганглиев (левого) показаны главные церебральные нервы, показаны также церебробуккальные коннективы и буккальные ганглии.
Рис. 11. Парные серотонинсодержащие гигантские нейроны в церебральных отделах нервной системы у представителей разных групп гастропод.
А — виноградная улитка; Б — слизень Limax; В — прудовик; Г — тритония. Парные церебральные ганглии или их производные (у тритонии — цереброплевральные ганглии) изображены с вентральной стороны, у одного из ганглиев (левого) показаны главные церебральные нервы, показаны также церебробуккальные коннективы и буккальные ганглии.
По-видимому, несимметричная группа нейронов передней доли соответствует мезоцеребральным нейронам виноградной улитки, которые также распределены несимметрично (правая группа значительно крупнее левой). Далее, я нашел, что отросток серотонинсодержащего нейрона делится на некотором расстоянии от клетки на две ветви, из которых одна направляется в церебральную коммиссуру. Наконец, удалось проследить, что описанный нами с И. Ж.-Надем аксон, лежащий в церебро-буккальном коннективе и имеющий «зелёное» свечение, на самом деле представляет собой аксонную ветвь гигантского серотонинсодержащего нейрона. В цитированной выше работе нами была допущена ошибка в оценке цвета люминесценции. Таким образом, для гигантских серотонинсодержащих клеток прудовика имеются, хотя и в меньшем объеме, данные, указывающие, что гомология этих клеток с нейронами ПЦ1 и ЛЦ1 подтверждается связями нейронов. Схематизированные иллюстрации к сказанному даны на рис. 11. Вывод о существовании в церебральных ганглиях лёгочных и заднежаберных моллюсков гомологичных нейронов, выделяющихся крупными размерами и специфическим химизмом, важен прежде всего в теоретическом плане, к чему мы вернёмся позже. Но хотелось бы обратить внимание и на возможные практические применения выводов такого рода. Во-первых, найденные с помощью цитохимических и цитофизиологических методов клетки-гомологи открывают новые возможности перед сравнительной анатомией, так как они позволяют находить топографические соответствия разных отделов ЦНС. Второй, не менее интересный практический выход — возможность прогнозировать свойства нейронов. В рассматриваемом случае можно с большой степенью уверенности дать следующий прогноз: у всех лёгочных и заднежаберных моллюсков пара выделяющихся крупными размерами нейронов, лежащих симметрично на передних концах церебральных ганглиев или (у стебельчатоглазых) сдвинутых на вентральную поверхность ганглиев, должна содержать серотонин и контролировать своими серотонинергическими окончаниями работу буккальных нейронов. Полезность прогнозов такого рода уже подтверждена практикой. У аплизии, как уже отмечалось выше, по неизвестной причине не удаётся, пользуясь гистохимическим методом Фалька и Хилларпа, выявлять биогенные амины и, следовательно, картировать аминергические клетки. Это, несомненно, тормозит развитие нейробиологических исследований на нейронах аплизии. Как я знаю из переписки с американским нейрохимиком Р. Мак-Каманом, руководимая этим исследователем группа, разработав методы количественного определения медиаторных аминов в отдельных нейронах, безуспешно пыталась искать моноаминергические клетки у аплизии вслепую. Задача намного упростилась, когда эти исследователи применили метод клеточных гомологии. В церебральныхганглиях аплизии были найдены парные гигантские нейроны, содержащие серотонин [331], что в свою очередь позволило французским физиологам получить исчерпывающие доказательства синаптической функции серотонина при изучении возбуждающих воздействий этих нейронов на клетки буккальных ганглиев аплизии [264].
5. 3. 2. Гомологи нейрона ППа1
Среди нейронов виноградной улитки гигантская клетка ППа1 выделяется уникальным типом залповой активности (см. 4.2.5.). Значительно раньше того времени, когда мы с Я. Шаланки нашли и описали эту клетку у виноградной улитки [283], совершенно такой же тип залповой активности был отмечен у одного из гигантских нейронов абдоминального ганглия аплизии. Разные авторы, работавшие с этим нейроном на аплизии, давали ему разные названия. В лаборатории Арванитаки и Халазонитиса его называют «Бр-клеткой», так как он находится в том месте, откуда вырастает бранхиальный (жаберный) нерв; соответственно, сами залпы называют «активностью Бр-типа». В работах Тауца некоторое время та же клетка носила несколько экстравагантное название «Оберон». Струмвассер называл её «параболическим залповиком», имея в виду, что межспайковые интервалы меняются в ходе залпа по параболической зависимости. Наконец, после работы по картированию абдоминального ганглия аплизии, проведённой группой Кандела, за клеткой утвердился индекс R15 (т. е. 15-я клетка правого полуганглия — по-русски П15) и это обозначение стало почти общепринятым. Список работ, выполненных на клетке П15 аплизии, настолько велик, что мы сошлёмся лишь на несколько важнейших, дающих библиографию [82, 115, 152, 160, 302]. Эта клетка имеется у разных видов рода аплизия, она занимает постоянную позицию в абдоминальном ганглии. Что касается клетки ППа1 виноградной улитки, то здесь нам приходится ограничиваться собственными данными. Но у других хелицид в соответствующем месте правого париетального ганглия тоже имеется гигантская клетка с такой же залповой активностью, несомненно гомологичная клетке ППа1. Гомология здесь не вызывает сомнений, так как совпадают не только свойства, но и позиции этих нейронов [162а, 1626, 210, 211, 214, 215]. Я предложил рассматривать ППа1 улитки и П15 аплизии как гомологичные клетки, опираясь на критерий специального качества, т. е. на совпадение широкого ряда специфических черт [54, 279]. Приведу аргументы в пользу этого мнения, суммируя, с одной стороны, результаты, полученные на клетке П15 разных видов рода аплизия, а с другой — на ППа1 виноградной улитки и на соответствующем нейроне других хелицид. Цвет - у аплизии прозрачно-оранжеватый у молодых особей, белёсый до белого у взрослых. У просмотренного нами большого числа особей виноградной улитки клетка, идентифицированная электрофизиологически, была чаще всего прозрачно-оранжеватой, но иногда в цитоплазме было белое содержимое, что придавало клетке белесоватый оттенок. Зависимость цвета нейрона ППа1 от возраста не изучена. Нейроцитологи знают, что белыми выглядят нейроны, содержащие так называемые элементарные нейросекреторные (лучше сказать — пептидергические) гранулы, светопреломляющие свойства которых и обеспечивают белизну. В самом деле, гранулы этого типа найдены в большом числе в рассматриваемых залповых нейронах как у аплизии [см. 302], так и у хелицид [см. 162а]. В последнем случае указан их диаметр, равный 2000 — 2500 Å. Потенциалы действия - как и в ППа1, в П15 аплизии регистрируются однокомпонентные спайки, генерируемые в соме и предваряемые медленным препотенциалом, имеющие большой овершут, задержку на нисходящей фазе, причём длительность задержки закономерно меняется в течение залпа. Такое сочетание свойств потенциалов действия уникально. Как у аплизии, так и у улитки имеется ряд нейронов, в которых спайки имеют большой овершут и задержку на нисходящей фазе, но во всех идентифицируемых клетках, кроме ППа1 и П15, такие потенциалы начинаются аксонным компонентом. Одинакова ионная зависимость потенциалов действия. В обоих случаях они продолжают генерироваться при исключении как ионов натрия, так и ионов кальция, но исключение обоих этих ионов подавляет генерацию. Вывод о том, что в обоих рассматриваемых нейронах входящий ток переносится ионами натрия и кальция, подтверждается и другими экспериментами (тетродоксин не подавляет потенциалов действия при нормальном содержании кальция, но подавляет при сниженном; ионы кобальта подавляют генерацию при исключении из раствора ионов натрия, и т. д.). Тип залповой активности, описанный для ППа1 в разделе 4.2, у обеих клеток одинаков, причём в обоих случаях он выявляется в чистом виде в отсутствие синаптического притока. Этот тип активности также уникален. У виноградной улитки он наблюдался нами лишь в клетках группы D, где такие залпы представлены довольно растянуто и невнятно. У аплизии залпы того же типа были отмечены лишь у одной из клеток белой группы [196]; по-видимому, залповая активность не является для них характерной, тогда как в клетке П15 эта активность наблюдается регулярно. Синаптический приток. Описание, данное нами для ППа1, вполне приложимо к П15. Здесь также представлены ВПСП, ТПСП, в числе которых наблюдаются имеющие начальную деполяризующую фазу гиперполяризующие ПСП, кумулирующиеся в торможение большой длительности. Реакция на медиаторные вещества у клетки П15 такая же, как у ППа1. Ацетилхолин вызывает сильную деполяризацию (ацетилхолин оказывает заметно более умеренное влияние на окружающие нейроны в том и другом случае). Серотонин также деполяризует обе клетки, тогда как дофамин в обоих случаях вызывает гиперполяризацию. Специфический химизм обоих нейронов одинаков и выражается в том, что они, в отличие от других исследованных нейронов, синтезируют какой-то полипептид (или, может быть, смесь полипептидов) молекулярным весом около 5000. X. Гайнер, обнаруживший этот факт, высказывает предположение, что этот специфический продукт синтеза может представлять собой активное начало секреторных гранул или белок-носитель активного начала [162, 162а]. Подводя итог, можно сказать, что клетки ППа1 и П15 идентичны по целому ряду качественных характеристик, что позволяет считать их гомологичными. Гомология, устанавливаемая с помощью критерия специального качества, получает полное подтверждение при рассмотрении этих клеток с позиций критерия положения. В отличие от нейрона ППа1, находящегося в окологлоточном кольце ганглиев, нейрон П15 лежит далеко от окологлоточного кольца, в области желудка, где у аплизии находится абдоминальный ганглий. Но это различие позиций двух нейронов на поверку оказывается сходством, потому что в обоих случаях рассматриваемые клетки лежат в составе одинаковых по происхождению нервных структур. Как уже упоминалось, правый париетальный ганглий улитки гомологичен супраинтестинальному ганглию примитивных гастропод; ему же гомологичен правый полуганглий абдоминального ганглия аплизии, в состав которого, возможно, входят также клеточные элементы паллиального ганглия предковых форм [192]. Во многом обоснованные сравнительно-анатомическими исследованиями представления о происхождении абдоминального ганглия аплизии, ганглиев висцеральной дуги улитки, а также соответствующих отделов ЦНС тритонии схематически показаны на рис. 12. Нейроны ППа1 и П15 не только лежат в участках, являющихся производными супраинтестинального ганглия, но и в пределах этих родственных областей занимают сходные позиции: обе клетки расположены на дорзальной поверхности, медиальнее корешка нерва, выходящего из этой области. Гомология нервов легко доказывается: у аплизии этот нерв иннервирует осфрадий (специальный орган химической чувствительности); у виноградной улитки осфрадия нет, но у имеющих этот орган водных и наземных пульмонат он иннервируется именно тем нервом, который выходит из правого париетального ганглия — внутренней ветвью правого мантийного нерва. Сходны не только позиции нейронов (если их рассматривать по отношению к другим сравнимым структурам), но и ход отростков. В обоих случаях аксон следует в переднем направлении, затем поворачивает налево и, покинув область супраинтестинального происхождения, через соответствующий коннектив попадает в область висцерального происхождения (у аплизии — это медиальная часть левого полуганглия, а у улитки — медиальная часть висцерального ганглия).
 Рис. 12. Сходные по ряду свойств залповые нейроны у представителей разных групп гастропод.
А — клетка ППа1 виноградной улитки (RPa1); Б — клетка П15 аплизии (R15); В — клетка № 22 тритонии. Показаны позиции нейронов в дорзальной клеточной коре соответствующих ганглиев — правого париетального (А), абдоминального (Б) и правого цереброплеврального (В) (верхний ряд рисунков). Показаны позиции названных ганглиев в теле соответствующего моллюска (средний ряд) и эволюционное происхождение разных частей ЦНС(нижний ряд). Все три клетки расположены в гомологичных участках нервной системы. Обозначения происхождения областей ЦНС для нижнего ряда рисунков: 1 — педальное, 2 — паллиальное, 3 — церебральное, 4 — субинтестинальное, 5 — плевральное, 6 — висцеральное, 7 — супраинтестинальное.
Рис. 12. Сходные по ряду свойств залповые нейроны у представителей разных групп гастропод.
А — клетка ППа1 виноградной улитки (RPa1); Б — клетка П15 аплизии (R15); В — клетка № 22 тритонии. Показаны позиции нейронов в дорзальной клеточной коре соответствующих ганглиев — правого париетального (А), абдоминального (Б) и правого цереброплеврального (В) (верхний ряд рисунков). Показаны позиции названных ганглиев в теле соответствующего моллюска (средний ряд) и эволюционное происхождение разных частей ЦНС(нижний ряд). Все три клетки расположены в гомологичных участках нервной системы. Обозначения происхождения областей ЦНС для нижнего ряда рисунков: 1 — педальное, 2 — паллиальное, 3 — церебральное, 4 — субинтестинальное, 5 — плевральное, 6 — висцеральное, 7 — супраинтестинальное.
Здесь аксон поворачивает назад и покидает ганглий через нерв, иннервирующий дериваты целома (т. е. через перикардиальный и генитальный нервы у аплизии и через гомологичный им интестинальный нерв у улитки). Кроме того, у П15 описана аксонная ветвь, направляющаяся в осфрадиальный (бранхиальный) нерв. По-видимому, такая ветвь есть и у соответствующей клетки наземных улиток; по крайней мере в одной из работ, вышедших из лаборатории Керкута, указывается на её существование [211], но в другой работе из той же лаборатории упомянуто только главное направление хода аксона [210]. Таким образом, клеточная гомология, опирающаяся на критерий специального качества, подтверждается анализом позиций клеток и их отростков, т. е. применением критерия положения. Следует добавить, что при картировании центральных нейронов тритонии также была найдена клетка (№ 22) с залпами, характерными для ППа1 и П15 [347]. Эта слабо пигментированная и имеющая белое содержимое клетка расположена на дорзальной поверхности правого церебро-плеврального ганглия в том его участке, который имеет супраинтестинальное происхождение [см. 102]. Вряд ли можно сомневаться в том, что клетка № 22 тритонии относится к рассматриваемому ряду гомологичных нейронов. К сожалению, в более поздней публикации [347а] авторы допустили непостижимую ошибку, положив в основу новой нумерации нейронов мысль, что каждый заметный нейрон должен быть парным,- мысль, ложность которой очевидна каждому, кто знаком с историей развития нервной системы гастропод. Наличие идентичных индивидуальных нейронов у таких далёких друг от друга форм, как наземная улитка, голожаберный моллюск тритония и представитель покрытожаберных — аплизия, дает полное основание прогнозировать существование нейрона с такими же свойствами у других заднежаберных и лёгочных. Этот нейрон должен быть расположен в составе клеточной группы супраинтестинального происхождения, на её дорзальной поверхности, должен выделяться крупными размерами и может иметь (в большем или меньшем количестве) белое содержимое в своей цитоплазме. В самом деле, после опубликования идеи о том, что клетка ППа1 имеет гомологов у других пульмонат и у заднежаберных, появилась статья, в которой впервые описана идентичная клетка у представителя отряда сидячеглазых — Helisoma [206]. Любопытно, что у этой улитки соответствующая клетка лежит не в правом, а в левом париетальном ганглии, но так и должно быть, если иметь в виду гомологию ганглиев (у планорбид, к которым относится Helisoma, расположение ганглиев и других внутренних органов представляет собой как бы зеркальное отражение того, что наблюдается у большинства гастропод). Следовательно, позиция залповой клетки у Helisoma полностью соответствует сформулированному выше прогнозу.
5. 3. 3. Гомологи группы пептидергических клеток
Следующим будет рассмотрен пример, демонстрирующий возможности электронно-микроскопического анализа при решении вопросов о клеточных гомологиях. Как было показано выше (4.4), в ЦНС виноградной улитки имеется много нейронов с секреторными гранулами пептидергического типа, причём характеристики этих гранул различны в разных группах клеток. С этой точки зрения хорошо изучен также прудовик, Lymnaea stagnalis, у которого детально описана ультраструктура пептидергических нейронов [336]. Мы попытались получить ответ на следующий вопрос: одинаковы ли характеристики секреторных гранул в соответствующих нейронах виноградной улитки и прудовика? Иными словами, выяснялся вопрос об устойчивости ультраструктурных характеристик секреции в ряду гомологичных клеток. Сравниваемые виды моллюсков достаточно далеки друг от друга в систематическом отношении (они представляют разные отряды подкласса пульмонат) и имеют совершенно различную экологию; вместе с тем у них легко найти гомологичные участки ЦНС и, в некоторых благоприятных случаях, можно, пользуясь критерием положения, найти гомологичные нейроны. Группа D правого париетального ганглия виноградной улитки представляет такую возможность. Клетки этой группы образуют особую долю ганглия, расположенную в нём дорзолатерально; для них характерна бледная или белёсая, до белой, окраска. При рассмотрении в микроскоп изолированного окологлоточного ожерелья прудовика в правом париетальном ганглии без труда обнаруживается расположенная латерально выпуклая доля, состоящая из однородных, довольно крупных бледных или белёсых клеток. Позиции этих долей в правых париетальных ганглиях обоих видов почти идентичны, разница лишь в том, что у виноградной улитки латеральный край ганглия не свободен, а прижат к плевральному, из-за чего доля с бледными клетками сдвинута на дорзальную поверхность (рис. 13). Нейроны, лежащие в этой доле у прудовика, изучены электронно-микроскопически. Они функционируют как нейросекреторные клетки, секретируя гранулы пептидергического типа под оболочку правого мантийного нерва или соматическими отростками — под оболочку ганглия [336]. Чтобы достовернее убедиться в гомологии рассматриваемых клеточных групп, мы провели два дополнительных исследования.
 Рис. 13. Сходные по ультраструктурным свойствам группы пептидергических нейро-секреторных клеток в правом париетальном ганглии виноградной улитки (А — собственные данные) и прудовика (Б - по 336). Показаны позиции клеточных тел и области секреции нейрогормона (оболочка ганглия и оболочка правых мантийных нервов). Обозначения см. на рис. 5.
Рис. 13. Сходные по ультраструктурным свойствам группы пептидергических нейро-секреторных клеток в правом париетальном ганглии виноградной улитки (А — собственные данные) и прудовика (Б - по 336). Показаны позиции клеточных тел и области секреции нейрогормона (оболочка ганглия и оболочка правых мантийных нервов). Обозначения см. на рис. 5.
Во-первых, названные клетки прудовика были исследованы электрофизиологически с помощью внутриклеточного электрода. Оказалось, что характеристики клеток соответствуют тем, которые известны для клеток группы D виноградной улитки (см. 4.2.5). Во-вторых, посредством инъекции проционового желтого в нейроны группы D виноградной улитки было определено направление их отростка. Результаты исследования [284] показали, что аксоны, как и у соответствующих клеток прудовика, следуют в правый мантийный нерв (в обе его ветви). Помимо этого, аксон каждой из исследованных клеток давал веточку в синаптический нейропиль. Как у прудовика, клетки посылают тонкие соматические отростки в оболочку ганглия. Представляется, что гомология рассматриваемых клеточных групп не вызывает сомнений. Обратимся теперь к ультраструктурным характеристикам секреторных гранул. Венделаар Бонга, описавший шесть типов пептидергических гранул в клетках ЦНС прудовика [336], отмечает, что гранулы рассматриваемой группы нейронов являются крупнейшими: их средний диаметр в аксонных окончаниях 2300 Å, а максимальные размеры достигают 3500 Å (в теле клетки средний диаметр несколько меньше — около 2100 — 2200 Å). Такие крупные гранулы найдены ещё в одной группе клеток, расположенной в висцеральном ганглии, больше нигде в ЦНС клетки с этим типом гранул не встречаются. Под наружной мембраной гранулы имеется прозрачная зона, остальная часть занята электронноплотным содержимым, которое разрешается как мелкозернистая масса. Очевидно, что эти характеристики совершенно приложимы к секреторным гранулам клеток группы D виноградной улитки (см. раздел 4.4.3). Как показали наши исследования, гранулы этих клеток — крупнейшие среди всех типов пептидергических гранул, найденных в ганглиях виноградной улитки, их внешний вид и диаметр такие же, как у гомологичных клеток прудовика. Добавим, что у обоих видов только в этих клетках ганглиев висцеральной дуги были найдены концентрически организованные агрегаты эргастоплазмы — субстанция Ниссля. Таким образом, ультраструктурные корелляты специфического химизма оказались одинаковыми в гомологичных клетках этих разных видов.
5. 3. 4. Гомологи катехоламиновых нейронов педальных ганглиев
Во всех разобранных до сих пор примерах клеточных гомологий имеющиеся материалы не давали возможности обсуждать вопрос о том, каким способом специфические нейроны появились в ганглии. Напомним, что априорно можно представить два пути их появления: либо клетки с разной химической специфичностью дифференцировались в процессе эволюции исходной однородной клеточной популяции ганглия, либо они были особенными ещё до ганглионизации. Клетки, о которых пойдёт речь теперь, позволяют исследовать этот вопрос. Сравнительные данные дают возможность проследить историю катехоламиновых нейронов, имеющихся в некотором числе в составе педальных ганглиев виноградной улитки. Из детального описания её моноаминергических нейронов, данного в разделе 4.3, извлечём несколько положений, важных для последующих сопоставлений. Во-первых, педальные ганглии, связанные с иннервацией ноги, богаче катехоламиновыми элементами, чем все другие ганглии, кроме, может быть, церебральных. Во-вторых, катехоламиновые нейроны составляют лишь малую часть нейронов педальных ганглиев. В-третьих, они расположены в основном группами и в значительной степени у корешков педальных нервов, а иногда в самих этих нервах. В-четвёртых, катехоламиновые аксоны, приходящие по педальным нервам, богато представлены в мышце ноги (их много и в дериватах педальной мускулатуры — в стенке пениса и в ретракторе пениса). Педальный отдел нервной системы претерпевает в пределах класса гастропод особенно сильную эволюцию, что делает его удобным объектом для анализа эволюционных закономерностей. Среди ныне живущих гастропод имеются представители древних групп, у которых педальный отдел находится на чрезвычайно примитивном уровне организации: ганглии отсутствуют, нейроны образуют в ноге род диффузной сети, несколько усложнённой наличием внутримышечных нервных тяжей. Таковы Docoglossa, представитель которых, акмея, был взят для данного сравнительного исследования. У форм с более сложной организацией этого отдела нервной системы значительная часть педальных нейронов ганглионизируется, и образующиеся при этом парные педальные ганглии выносятся за пределы ноги, а затем вместе с другими ганглиями ЦНС подвергаются концентрации, образуя компактный мозг. Одновременно идёт процесс гистологической дифференциации педальных ганглиев: в них появляются гигантские нейроны, стратифицируется синаптический нейропиль. В таком продвинутом виде педальные отделы представлены не только у виноградной улитки и других стебельчатоглазых, но и у ряда заднежаберных моллюсков, например у голожаберных (хотя у всех, по-видимому, гастропод какая-то часть педальных нейронов остаётся на периферии, в педальном нервном сплетении). Представитель подкласса заднежаберных — дендронот также был взят для данного сравнительного исследования. Кроме этих форм, представляющих «низший» и «высший» пределы эволюции педального отдела, исследованию были подвергнуты несколько других представителей класса. Катехоламинергические клетки и волокна выявлялись водным формальдегидным методом. Работа проводилась совместно с А. В. Сахаровой. Чтобы не потерять из виду интересующие нас клетки, мы двигались небольшими шагами, с каждым шагом раздвигая на один таксономический ранг круг охваченных объектов. Кроме представителей рода Helix, о которых шла речь в главе 5, мы исследовали некоторых других хелицид: Eobania vermiculata и Caucasotachea sp. Анатомическая организация педальной нервной системы у них такая же, как у виноградной улитки. Картина катехоламиновых элементов также сходна: богатый синаптический нейропиль в педальных ганглиях; группы преимущественно мелких нейронов, расположенные так же, как у виноградной улитки; большое число аксонов в мышце ноги, где наблюдаются островки синаптического нейропиля рядом с группками нейронов, не содержащих биогенных аминов. Вслед за хелицидами были исследованы представители трёх других семейств отряда стебельчатоглазых. Xeropicta candaharica (сем. Hygromiidae). Небольшое отличие от хелицид заключается лишь в том, что люминесцирующие катехоламиновые аксоны образуют в ноге заметное терминальное сплетение непосредственно под эпителием. Впрочем Роджерс [275а] отмечает то же самое у садовой улитки, но у виноградной выходы неварикозных аксонов под эпителий, хотя и имеют место, не носят такого выраженного характера. Bradybaena fruticum (сем. Bradybaenidae). Заметных отличий от хелицид в картине катехоламиновых элементов педальной системы не наблюдается. Limax maximus (см. Limacidae). Хорошо выражены неварикозные, довольно толстые терминальные ветвления катехоламиновых аксонов под эпителием подошвы ноги; варикозный синаптический нейропиль организован в ноге не беспорядочными островками, как у рассмотренных выше видов, а строго ориентированными двумя продольными рядами таких островков. Эти наши наблюдения не соответствуют данным Осборна и Коттрелла, которые, изучив тот же вид наземных слизней, пишут, что в ноге «наблюдается очень мало моноаминовых волокон» [262]. Тела катехоламиновых нейронов расположены в педальных ганглиях примерно в той же позиции, что у виноградной улитки. Размеры мелких, расположенных группами, клеток, по измерениям Осборна и Коттрелла, около 15 мк, тогда как большая непарная клетка, вероятно соответствующая непарной клетке виноградной улитки, достигает у слизня 100 мк. Литературные данные имеются, кроме того, для Strophocheilus oblongus (сем. Strophocheilidae). Эта гигантская тропическая улитка изучена гистохимически бразильскими авторами [195] и полученные результаты очень интересны: в педальных ганглиях катехоламиновые нейроны отсутствуют. Факт этот тем более примечателен, что в церебральных ганглиях картина серотониновых и катехоламиновых нейронов оказалась типичной для стебельчатоглазых пульмонат, в частности, как уже отмечалось, бразильская улитка имеет пару гигантских нейронов — гомологов ПЦ1 и ЛЦ1. Сообщается, что нейропиль всех подглоточных ганглиев содержит катехоламиновые аксоны, которые предполагаются отростками клеток, лежащих на периферии. К сожалению, авторы не исследовали ногу бразильской улитки. Кроме отряда стебельчатоглазых, в подкласс лёгочных входит отряд сидячеглазых улиток. Мы исследовали два вида. У прудовика, Lymnaea stagnalis, имеющего хорошо выраженные педальные ганглии, катехоламиновые нейроны педального отдела отличаются от соответствующих клеток виноградной улитки только, пожалуй, значительно более крупными размерами. Группы этих нейронов расположены в основном в латеральных частях обоих ганглиев. Непарная правая клетка, расположенная, как и у виноградной улитки, дорзально, может быть отнесена к гигантским нейронам. Нейропиль богат катехоламиновыми контактами. В ноге — только аксоны, они выходят к эпителию только у края подошвы. В пенисе, стенка которого имеет педальное происхождение, много варикозных волокон на мышечных клетках субэпителиального слоя мышц. У американской катушки Helisoma (из планорбид) нами найдена примерно такая же, как у прудовика, картина в педальных ганглиях и несколько отличная в ноге. Здесь совсем отсутствуют островки синаптического нейропиля (у прудовика их было очень мало); гладкие, неварикозные катехоламиновые аксоны, прослеживаемые вдоль педальных нервов от самых педальных ганглиев, проходят через толщу ноги и достигают субэпителиальной области, где далее следуют параллельно поверхности под эпителиальными клетками. Литературные данные имеются для педальных ганглиев катушки Planorbarius corneus; здесь, как и у прудовика, отмечен непарный гигантский катехоламиновый нейрон, достигающий 280 — 300 мк [238], который, как показали последующие исследования, является интернейроном, возбуждающим одни и тормозящим другие центральные нейроны [92]. Рассмотрим теперь несколько форм, наиболее далеких от виноградной улитки и представляющих два других подкласса — переднежаберных и заднежаберных. Из заднежаберных имеются данные для двух видов голожаберных слизней — тритонии и дендронота. В педальных ганглиях тритонии М. С. Манохина и Л. В. Кузьмина не нашли катехоламиновых нейронов [37]. Также не нашли их и мы в педальных ганглиях дендронота. В отличие от тритонии, у дендронота мы смогли исследовать не только ЦНС, но и периферию. Как в ноге, так и в её деривате — стенке пениса имеется чрезвычайно богатая сеть варикозных катехоламиновых волокон, контактирующих с мышечными клетками. Эти волокна являются отростками клеток, которые расположены в самих этих органах. Каждая катехоламиновая клетка посылает также гладкий, неварикозный отросток в ЦНС, где такие отростки дают начало варикозному синаптическому нейропилю. Лежащие на периферии педальные катехоламиновые нейроны дендронота были подвергнуты дополнительному электронномикроскопическому исследованию. В подтверждение данных люминесцентной гистохимии было найдено, что в пенисе эти клетки всегда расположены у внутреннего края мышечной стенки, они мелкие, биполярные. Один отросток идет между мышечными клетками в периферическом направлении. Он древовидно ветвится; по ходу ветвей имеются типичные пресинаптические аксональные расширения, контактирующие с клетками второго (снаружи) мышечного слоя. Эти расширения наполнены секреторными гранулами, которые (типично для катехоламинов) имеют плотное зерно при глутаральдегидной префиксации, но лишены его при фиксации одной лишь четырёхокисью осмия. Никогда не наблюдаются контакты между этими аксонами и мышечными клетками наружного слоя, которые вообще не иннервированы. С противоположной стороны от тела нейрона отходит другой отросток, который вступает в состав нервного стволика, идущего между стенкой пениса и семепроводом. Очевидно, это нерв, следующий в ЦНС. На теле катехоламинового нейрона нередко можно видеть синаптические окончания. Обратимся теперь к переднежаберным моллюскам. Литературных данных о педальных катехоламиновых нейронах для этого подкласса нет, нами исследованы три вида. У примитивной акмеи Testudinalia (Acmaea) tessellata, не имеющей педальный ганглиев, катехоламиновые нейроны лежат в ноге на небольшом расстоянии от эпителия. Эти клетки биполярные: наружный отросток, лишённый варикозных утолщений, ветвится под клетками эпителия; внутренний направляется к находящемуся в толще ноги педальному ганглиозному стволу, вступая по пути и в самом стволе в состав варикозного катехоламинового синаптического нейропиля. В самих парных ганглиозных педальных стволах, богатых катехоламиновыми нервными окончаниями, отсутствуют нейроны, содержащие катехоламины.
 Рис. 14. Центральная нервная система переднежаберного моллюска Pomatiaselegansи позиции центральных нейронов, содержащих биогенные амины. Обозначение ганглиев: 1 — церебральный, 2 — педальный, 3 — плевральный, 4 — супраинтестинальный, 5 — висцеральный, 6 — субинтестинальный, 7 — буккальный, 8 — тентакулярный. В ганглиях 4 — 6 выявление биогенных аминов не проводили. В плевральных и тентакулярных ганглиях моноаминергические нейроны не обнаружены.
Рис. 14. Центральная нервная система переднежаберного моллюска Pomatiaselegansи позиции центральных нейронов, содержащих биогенные амины. Обозначение ганглиев: 1 — церебральный, 2 — педальный, 3 — плевральный, 4 — супраинтестинальный, 5 — висцеральный, 6 — субинтестинальный, 7 — буккальный, 8 — тентакулярный. В ганглиях 4 — 6 выявление биогенных аминов не проводили. В плевральных и тентакулярных ганглиях моноаминергические нейроны не обнаружены.
Другой исследованный вид — аквариальная «ампуллярия», которая, по сообщению А. А. Шилейко, представляет собой Pila (globosa?) из сем. Pilidae. Улитки этого семейства относятся к группе Architaenioglossa, для которой также характерно отсутствие педальных ганглиев. Парные педальные ганглиозные тяжи связаны многочисленными комиссурами. Такое строение педальной нервной системы рассматривается как древний признак. В главных педальных стволах и в дочерних тяжах, доходящих почти до подошвы, богато представлены участки синаптического нейропиля с варикозными катехоламиновыми волокнами. Тела катехоламиновых нейронов встречаются в составе ганглиозных стволов. В стенке пениса тела этих нейронов расположены более периферически. Нейроны биполярны, причём периферический отросток не только доходит до эпителия, но и даёт веточки между эпителиальными клетками. Внутренний отросток следует к островкам синаптического нейропиля, расположенным тут же, в стенке органа. Наконец, третий вид — Pomatias elegans (сем. Pomatiasidae) относится к группе Taenioglossa, представители которой имеют компактные педальные ганглии. В составе этих ганглиев, наряду с большинством нейронов, лишённых биогенных аминов, имеются латерально расположенные группки мелких катехоламиновых клеток (рис. 14). Нейропиль ганглиев богат варикозными катехоламиновыми волокнами. В ноге тоже имеются мелкие катехоламиновые нейроны. Их отростки не подходят к эпителию подошвы. В стенке пениса, которая у этой улитки не является дериватом педальной мышцы, а вырастает из спинной области, катехоламиновых волокон нет совсем. Эти сравнительные данные позволяют реконструировать картину эволюции катехоламиновых нейронов, входящих в состав педальной нервной системы. Вряд ли можно сомневаться, что их исходная позиция представлена у примитивной акмеи, где эти клетки занимают субэпителиальное положение в подошве ноги. Биполярные эпителиальные или субэпителиальные нейроны обычно рассматриваются как нейроны с сензорной функцией, которую выполняет периферический отросток. Отростки педальных катехоламиновых нейронов проявляют, однако, три рода функциональных отношений. Неварикозный отросток, расположенный под эпителием или входящий в него, очевидно, выполняет сензорные функции; варикозный периферический или центральный отросток, контактирующий с мышечными клетками, мы вправе считать моторным; наконец, варикозный отросток, контактирующий с другими нейронами, служит для осуществления межнейронных связей. Все три рода отношений можно найти в разных группах гастропод. По-видимому, педальные катехоламиновые нейроны, исходно мультифункциональные, в ходе эволюции приобретали разную специализацию, став сензорными в одних случаях, моторными — в других (например, в пенисе дендронота), интернейронами — в третьих. Непарный дофаминергический интернейрон заслуживает особого упоминания. Берри и Коттрелл [92] исследовали синаптические эффекты этого гигантского нейрона в ЦНС катушки, где он расположен в левом педальном ганглии (ганглий ошибочно обозначен правым в работе [238]). Сравнительно-гистохимические данные, изложенные выше, показывают, что гомологи этой клетки легко прослеживаются в ряду пульмонат. По моим наблюдениям, у прудовика этот нейрон особенно велик и его легко найти в правом педальном ганглии (гомологичном левому ганглию планорбид) рядом с плевро-педальным коннективом; крупные размеры нейрон имеет у слизня, но у хелицид он невелик. В разных подклассах гастропод педальные катехоламиновые нейроны по-разному вовлечены в процесс ганглионизации. Налицо какая-то избирательность этого процесса, но природа её неясна, хотя общая тенденция прослеживается чётко. Так, среди лёгочных только в примитивном семействе Strophocheilidae нейроны этого типа не обнаружены в составе педальных ганглиев. То, что такие нейроны остались на периферии и у голожаберных, возможно, объясняется их моторными функциями. Теперь можно ответить на вопрос о том, каким способом катехоламиновые нейроны появились в составе нейронной популяции педальных ганглиев виноградной улитки. Сравнительные данные позволяют с уверенностью утверждать, что клетки с этим типом химической специфичности не возникли в процессе эволюционного дифференцирования исходно однородной нейронной популяции ганглиев: они как специфичная группа существовали до возникновения ганглиев. В процессе дивергентной эволюции гастропод эти клетки ганглионизировались или оставались на периферии; одновременно они приобретали разную функциональную специализацию, что, однако, не сказывалось на их медиаторном химизме.
5. 3. 5. Другие примеры клеточных гомологий
В контексте данной книги, рассматривающей вопросы теории, важен самый факт существования отдалённых клеточных гомологии в нервных системах — вернее говоря, важны свидетельства устойчивости специфического химизма в гомологичных клетках. С этой точки зрения приведённых выше примеров вполне достаточно. Всё же можно упомянуть несколько других примеров, представляющих результаты проведённой работы и полезных для последующего обсуждения. Катехоламиновые нейроны. Напомню, что у виноградной улитки нейроны, содержащие первичный катехоламин (по-видимому, дофамин), имеются в церебральных, педальных, буккальных ганглиях и в ганглиях нижней пары щупалец. Гомология педальных катехоламиновых нейронов рассмотрена выше. Коротко об остальных клетках. В церебральных ганглиях всех исследованных нами стебельчатоглазых улиток и слизней (Eobania, Caucasotachea, Bradybaena, Limax, Deroceras) общий характер картины одинаков. Группы мелких катехоламиновых нейронов и исключительно богатый их отростками нейропиль всегда имеются в педальной доле метацеребрума. Несколько крупных нейронов этого типа и ветвление их отростков с крупными варикозными расширениями найдены на дорзальной стороне метацеребрума. У прудовика, несмотря на сильное анатомическое отличие его церебрального ганглия от ганглия стебельчатоглазых, характер распределения в принципе тот же: вентральная доля, соответствующая педальной доле стебельчатоглазых, резко выделяется богатством катехоламинового нейропиля и присутствием групп мелких нейронов этого типа; ещё одна группа, по-видимому, соответствует той, которая у виноградной улитки лежит возле гигантского серотонинового нейрона (но у прудовика она несколько отдалена от серотонинового нейрона); наконец, на дорзальной поверхности имеется несколько обособленно лежащих крупных катехоламиновых клеток, в том числе одна клетка, размером около 50 мк, посылающая отросток в латеральную дольку мозга. Наиболее интересно, что и у обеих переднежаберных улиток, у которых мы изучили церебральные ганглии (Testudinalia tessellata и Pomatias elegans), сохраняется тот же характер распределения: богатая катехоламиновым нейропилем задне-вентральная часть ганглия резко ограничена от почти не проявляющей люминесценции передне-дорзальной части нейропиля. Катехоламиновые нейроны, как и у лёгочных, расположены в клеточной коре отдельными небольшими группками. Буккальные ганглии, исследованные нами у тех же самых видов, всегда содержат небольшое число катехоламиновых нейронов, расположенных примерно так же, как у виноградной улитки. Исключение, по литературным данным, составляет примитивная легочная улитка Strophocheilus oblongus, в буккаль-ных ганглиях которой, как и в педальных, авторы [195] не нашли катехоламиновых клеток. Напротив, в нижней паре щупалец упомянутые авторы нашли у бразильской улитки скопление катехоламиновых нейронов. Очень любопытно, что эти клетки расположены в эпителии, тогда как у виноградной улитки в эпителий приходят лишь их периферические отростки, а тела нейронов лежат в щупальцевом ганглии. Нет сомнения, что здесь мы встречаемся с тем же явлением миграции исходно периферических нейронов, которое выше рассматривалось на материале педальных нейронов. Другими сравнительными данными по этим клеткам мы пока не располагаем. Группа дающих люминесценцию нейронов, занимающая область границы между правым париетальным и висцеральным ганглием, обнаруживается в той же позиции у всех исследованных стебельчатоглазых. Такая же группа жёлтых клеток описана в соответствующем месте ЦНС сидячеглазой улитки — катушки [238]. Ранее мы для прудовика отметили наличие «зелёных», т. е. катехоламиновых, клеток в висцеральном ганглии [286], что, по-видимому, было ошибкой в оценке цвета люминесценции: эти результаты нуждаются в переисследовании. Данных по висцеральной дуге переднежаберных пока нет. Таким образом, соответствие в распределении катехоламиновых клеток и волокон обнаруживается в широком диапазоне форм, от примитивного переднежаберного моллюска — акмеи до представителей разных групп лёгочных. Добавим, что вплевральных ганглиях у всех изученных моллюсков, как и у виноградной улитки, отсутствуют моноаминергические клетки. Волокна, проявляющие высокую активность дегидрогеназы глюкозо-6-фосфата (ДГ Г6Ф). Нами с А. В. Сахаровой было обнаружено, что в ЦНС улиток имеется особая система волокон, которые выделяются на общем фоне высокой активностью дегидрогеназы глюкозо-6-фосфата (тетразолиевый гистохимический метод). Помимо виноградной улитки, реакция для выявления этого фермента была проведена на нескольких видах стебельчатоглазых (Caucasotachea sp., Limax maximus, Deroceras reticulatum). Из сидячеглазых был исследован прудовик, из заднежаберных — голожаберный моллюск эолидия (Aeolidia papillosa) и из переднежаберных — Pomatias elegans. Во всех случаях исследовалась ЦНС (в полной серии срезов) и иногда дополнительно — периферические ткани головной области. У всех изученных стебельчатоглазых волокна этого типа распределены в совершенно одинаковых участках нейропиля церебральных ганглиев. Отсюда единичные волокна этого типа следуют в ганглии подглоточного комплекса. У прудовика такие волокна в ЦНС не обнаружены (периферия не исследована). У эолидии в церебральных отделах обоих церебро-плевральных ганглиев имеется чётко выраженный пучок таких волокон, распадающийся на тонкие ветви в синаптическом нейропиле. Проявляющие высокую активность ДГ Г6Ф крупные пресинаптические окончания этих волокон хорошо видны на срезах. Волокна этого типа у эолидии не выходят за пределы церебральных отделов ганглиев, т. е. не проникают в области, соответствующие подглоточным ганглиям улитки. Наконец, у Pomatias тоже только в церебральных ганглиях имеется участок нейропиля (передняя его зона), проявляющий активность этого фермента. Некоторые дополнительные сведения дают наши электронномикроскопические данные [353]. В церебральных ганглиях стебельчатоглазых волокна, богатые ДГ Г6Ф, заполняют внутренний нейропиль процеребрума, но не соседний терминальный нейропиль. Обе зоны нейропиля изучались в электронный микроскоп у виноградной улитки и слизня Limax maximus. У того и другого вида в зоне, которая богата дегидрогеназой глюкозо-6-фосфата, электронный микроскоп обнаруживает большое число особых тонких аксонов, содержащих не пузырьки или везикулы, а «извитые трубочки». Аксоны этого рода отсутствуют в терминальном нейропиле. Добавлю, что помимо указанных волокон, в процеребруме слизня найдены другие нервные окончания, соответствующие по своей ультраструктуре окончаниям этой же области виноградной улитки [352, 353]. Подводя итог этим во многом ещё предварительным данным, следует отметить, что до сих пор не выяснено, где у виноградной улитки или другого вида находятся тела нейронов, аксоны которых в своей концевой, пресинаптической части содержат активную дегидрогеназу глюкозо-6-фосфата. Неизвестно, каков медиатор этих окончаний — по крайней мере, ни один из известных медиаторов не проявляет характерной связи с этим ферментом. Своеобразная ультраструктура этих аксонов, по-видимому, свидетельствует о том, что они относятся к какому-то новому, ранее не изученному химическому типу. Вместе с тем ясно, что этот тип волокон прослеживается от переднежаберных до лёгочных моллюсков. Сравнительные данные, хотя их и немного, говорят о том, что первичная область распространения таких волокон — ограниченная зона волокнистого вещества церебральных ганглиев. У наземных лёгочных они распространяются отсюда в подглоточные ганглии, а у водных сидячеглазых лёгочных (прудовик), возможно, остаются на периферии или выпадают по какой-то причине из состава ЦНС. Волокна, иннервирующие мышцы. В заключение мы попытаемся сопоставить отрывочные сведения о медиаторах моторных окончаний на мышцах гастропод. Вообще, у моллюсков очень много разных образований, содержащих мышечные клетки, и можно только догадываться о том, насколько разнообразны медиаторные механизмы нервной регуляции мышц. Упомянем, что только на мышечные клетки сердца действуют нервные окончания нескольких разных типов. Почти не изучена иннервация мышечных клеток желудочно-кишечного тракта и половой системы. И всё же имеющиеся данные, при всей их фрагментарности, очень интересны в плане обсуждающихся в этой книге вопросов. Дело в том, что у гастропод мышечные органы одинакового происхождения часто выполняют разную функцию (например, у виноградной улитки орган локомоции — нога, совокупительный орган и его ретракторная мышца имеют одинаковое педальное происхождение) и, наоборот,- мышцы разного происхождения могут выполнять одинаковую функцию (например, стенка пениса может образовываться не только из педальной мышцы, но и из мышцы спинной области, из головного щупальца [см. 19]). Сравнительные данные о специфичности мотонейронов должны показать, чем эта специфичность определяется — функцией или происхождением. Из практического опыта многих исследователей известно, что выпадение моллюска из раковины, т. е. расслабление колумеллярной мышцы, вызывается холинолитиками — например, тубокурарином или сукцинилхолином. Эти вещества оказывают расслабляющее действие на колумеллярную мышцу всех гастропод, включая примитивных, но вместе с тем не вызывают расслабления мышцы ноги или стенки тела. Этот факт сам по себе служит указанием на сходную по химизму иннервацию родственных мышц. Ретрактор глазного щупальца стебельчатоглазых пульмонатсчитается дериватом колумеллярной мышцы. Как уже было отмечено, по нашим электронно-микроскопическим данным, моторные окончания на мышечных клетках ретрактора содержат прозрачные синаптические везикулы диаметром 470 Å, т. е. везикулы, сходные с теми, которые содержатся в холинергических нервных окончаниях позвоночных. В опытах на изолированном препарате, содержащем тентакулярный нерв и ретрактор щупальца виноградной улитки, мы совместно с Л. В. Сорокиным нашли, что ацетилхолин действительно вызывает сокращение мышцы, тогда как холинолитики снимают его эффект и блокируют нервно-мышечную передачу. Из ряда испытанных литиков наиболее активными оказались D-тубокурарин (амплитуда сокращений в ответ на непрямое раздражение снижалась вдвое при 2•10-6 г/мл) и сукцинилхолин (то же, при 1,2•10-5 г/мл). Митолон — сильнейший блокатор холинорецепции в сердце улитки — оказался неактивным. Эти данные показывают, что разные мышцы улитки имеют совершенно разные холинорецепторы и что по своим рецепторным свойствам изученная ретракторная мышца представляет собой вариант того же типа, который включает соматическую мускулатуру позвоночных, мышцу стенки тела нематод и спинную продольную мышцу аннелид. Случайно ли это сходство, должны показать специальные исследования, посвящённые отдалённым гомологиям мышц. Нельзя, однако, не упомянуть совершенное независимое от фармакологических данных мнение известного сравнительного морфолога Лемке, что колумеллярная мышца заднежаберных гастропод филогенетически соответствует продольной мышце аннелид [230, стр. 59 - 60]. Все эти наблюдения указывают на то, что колумеллярная мышца и её дериват, ретрактор омматофора, получают моторные окончания от нейронов холинергического типа. В отличие от только что рассмотренной мышцы, имеющей, судя по фармакологическим и электронно-микроскопическим данным, однородную иннервацию, педальная мышца и её дериваты имеют моторные окончания нескольких разных типов. Эта смесь представлена одинаково у разных моллюсков и в разных мышцах педального происхождения. Электронно-микроскопически один и тот же набор характерных типов нервных окончаний, различающихся по ультраструктуре секреторных гранул, найден в таких разных мышцах, как мышца ноги садовой улитки [275а], ретрактор пениса виноградной улитки [см. 4.3.4], стенка пениса голожаберного моллюска — дендронота (собственные наблюдения). Для двух последних из названных объектов, имеющих регулярное строение, показано, что мышечные клетки с разными типами иннервации, расположены в разных слоях соответствующего мышечного органа. Это наблюдение свидетельствует о том, что педальная мускулатура имеет сборное происхождение и представлена разнородными мышечными (и соответственно нервными) элементами. В составе педальной мускулатуры закономерно встречаются мышечные элементы, получающие иннервацию от катехоламиновых клеток, о чём уже шла речь выше [5.3.4].Нужно, однако подчеркнуть, что мышечные элементы с этим типом иннервации характерны не только для ноги и её дериватов — их распространение гораздо шире. Так, варикозные катехоламиновые волокна неоднократно наблюдались нами у разных видов гастропод в мышечном слое пищевода, а также в сборной, по-видимому, мускулатуре стенки тела. Вместе с тем в некоторых мышцах, например в колумеллярной, катехоламиновые нервные волокна закономерно отсутствуют. Ещё раз отмечу, что катехоламиновые нервные волокна никогда не наблюдались нами в мышечной стенке пениса Pomatias elegans. У этого вида, в отличие от большинства гастропод, пенис образуется на спинной стороне животного. У всех форм, имеющих пенис педального происхождения, катехоламиновые волокна в его стенке обнаруживаются без исключения. Отмечу ещё, что мы нашли катехоламиновые нервные волокна в контакте с мышечными клетками в половом биче наземных лёгочных, т. е. в том органе, в котором X. С. Коштоянц впервые отметил исключительно высокую чувствительность мышцы к физиологически активным веществам. В сердечной мышце прудовика на тотальных плёночных препаратах я наблюдал негустую сеть аксонов, проявлявших специфическое свечение жёлтого, как мне показалось, цвета. Наличие моноаминергических волокон отмечено рядом авторов в сердце некоторых других видов гастропод [см. обзоры 333, 334].
5. 4. Заключение
Если в предыдущей главе было показано, что нейроны виноградной улитки качественно неодинаковы и что клеточная мозаика нервной системы проявляет у этого моллюска постоянство и повторяет свой рисунок от особи к особи, то в настоящей главе представлены свидетельства тому, что идентичные элементы клеточной мозаики — нейроны и группы нейронов можно обнаруживать при переходе от вида к виду, от одной группы моллюсков к другой. Даже у далеких друг от друга форм можно находить клеточные соответствия, в основе которых, как показал анализ, лежит гомология. Одновременно обнаружено, что именно гомологичные нейроны проявляют одинаковый специфический (медиаторный) химизм. Так, гигантские парные церебральные нейроны лёгочных и заднежаберных моллюсков можно гомологизировать, опираясь на данные об их иннервационных отношениях и пользуясь критерием положения; при этом оказывается, что все клетки, относящиеся к этому гомологическому ряду, содержат одно и то же медиаторное вещество — серотонин. Устойчивость медиаторного химизма облегчает обратную операцию — установление гомологии, опирающееся на знание специфических качеств нейрона. В этой главе был также представлен фактический материал, свидетельствующий о том, что нейроны не приобретают специфический медиаторный химизм в ходе прогрессивной эволюции ганглия — они приходят в ганглий с готовым особенным химизмом, который имеют на примитивной стадии своей истории, до ганглионизации. Таким образом, представленные данные и их анализ позволяют с полным основанием заключить, что можно гомологизировать клетки, входящие в состав нервной системы разных гастропод, и что для гомологичных нейронов характерен медиаторный изохимизм. Эти главные выводы можно сопроводить небольшими комментариями. Во-первых, хотелось бы обратить внимание на то, что сделанные выводы относятся ко всем трём подклассам гастропод. Хотя основная часть рассмотренных экспериментальных данных получена на гигантских нейронах (а таковые имеются только у заднежаберных и лёгочных), доступные факты о нервных клетках переднежаберных показывают, что в этом подклассе состав нейронной популяции примерно такой же, как и в двух других. Известно, что заднежаберные и лёгочные произошли от переднежаберных, поэтому мы вправе были бы ожидать, что нервная организация переднежаберных, по крайней мере древних групп этого подкласса, должна проявлять черты примитивности. Такие черты в самом деле можно обнаружить, если принимать во внимание такие показатели, как степень ганглионизации нервных клеток, степень концентрации ганглиев, дифференциация нейронов по размерным характеристикам, стратификация синаптического нейропиля. Тем более интересно, что по качественному разнообразию нейронов переднежаберные не проявляют заметных отличий, насколько можно судить из имеющихся данных о моноаминергических (5.3.5, а также [300]) и пептидергических [79] нейронах. Наблюдая изохимизм гомологичных нейронов внутри класса гастропод, мы не имеем оснований сомневаться, что и за пределами класса должно продолжаться то же явление, т. е. что одинаковую медиаторную специфичность будут проявлять клетки одинакового происхождения. В самом деле, доступные данные показывают, что при переходе от класса к классу гомологичные системы сохраняют у моллюсков свой особенный химизм. В качестве примера можно сослаться на двустворок: в их педальной системе имеются катехоламиновые нейроны, соответствующие таковым гастропод, такие же соответствия наблюдаются в качественном составе экстракардиальных нервных элементов и т. д. [см. ссылки в 279 и 333]. Наконец, данные об изохимизме гомологичных нейронов важны потому, что они позволяют полнее представить картину разнородности нейронной популяции гастропод. С этой целью мы вправе объединить сведения, накопленные при изучении разных представителей класса. В нейробиологии позвоночных это делается уже давно, и когда нужно представить медиаторное разнообразие нервной системы человека, с полным основанием учитываются факты, добытые на крысе, кошке, кролике и т. д. Теперь у нас есть уверенность в том, что так же можно объединять данные по гастроподам. Сведя эти данные воедино, мы сможем оценить богатство медиаторного фонда этих животных, что в свою очередь позволит нам в следующей главе основательнее обсуждать интересующие нас вопросы теории.
6. ПРОИСХОЖДЕНИЕ СПЕЦИФИЧЕСКОГО ХИМИЗМА НЕРВНЫХ КЛЕТОК
Теперь можно, видимо, вернуться к нашему главному вопросу — вопросу о причинах качественного разнообразия, нейронов у млекопитающих. По крайней мере, некоторые стороны этого вопроса должны проясниться. Начнем с того, что попробуем представить себе суммарную картину качественной разнородности популяции нервных клеток у гастропод — подобно тому, как это делалось в главе 2 для нейронной популяции млекопитающих. Консервативность медиаторного химизма, продемонстрированная в предыдущей главе, позволяет оценивать клеточный состав нервной системы гастропод, используя совокупно данные, полученные на разных представителях класса. Ещё раз отметим, что для предстоящего обсуждения особенно важно знать, в какой степени разнородна эта нейронная популяция и насколько «чисто» в ней представлен каждый из типов специфичности. Когда это возможно, будем также принимать во внимание, какую функцию выполняют клетки, относящиеся к тому или иному типу медиаторной специфичности.6. 1. Медиаторная специфичность нейронов гастропод
Холинергические нейроны. Исчерпывающие доказательства медиаторной функции ацетилхолина получены для некоторых крупных нейронов аплизии. Измеряя активность ацетилтрансферазы холина в индивидуальных клетках абдоминального ганглия две группы исследователей независимо друг от друга нашли, что энзим имеется только в трёх крупных нейронах — П2, Л10 и Л11; клетка, парная к П2 и расположенная в левом плевральном ганглии, также способна синтезировать ацетилхолин [169, 241]. Клетка Л10 — это интернейрон, оказывающий как возбуждающие, так и тормозные влияния на другие клетки ганглия; эти влияния во всех отношениях подобны эффектам апплицируемого ацетилхолина [96]. При инъекции в тело нейрона меченого холина последний превращается клеткой П2 в ацетилхолин и в фосфорилхолин, тогда как контрольные нейроны, не содержащие ацетилтрансферазу холина, используют холин только для синтеза фосфорилхолина [222]. Микрохимически ацетилхолин определяется в перикарионе названных выше холинергических нейронов аплизии в концентрации 0,35±0,01 мМ, в нехолинергических гигантских клетках уловить присутствие ацетилхолина не удаётся, так что специфичность химизма очень высока [241а]. Кроме клетки Л10, у аплизии описаны другие холинергические интернейроны, расположенные в плевральных и буккальных ганглиях. На других гастроподах изучение таких интернейронов не проводилось, но полагаю, что вполне реальными были бы поиски, например, гомолога Л10 у других лабораторных гастропод, в частности, у виноградной и садовой улитки, у которых эта клетка должна, по-видимому, располагаться на вентральной поверхности абдоминального ганглия. Шагом на пути к идентификации холинергических интернейронов может стать работа по обнаружению клеток, способных к синтезу ацетилхолина. Судя по доступным тезисам доклада, такую работу начала группа английских авторов [150]. Можно с уверенностью думать, что холинергическими у гастропод бывают не только интернейроны. Давно считается, что ацетилхолин служит медиатором в некоторых нервных окончаниях на сердце моллюсков, при этом со стороны сердечной мышцы известно два типа ответов на ацетилхолин — торможение биений и тоническая реакция [см. обзор 333, а также 233а]. Клетки, дающие начало тормозным волокнам сердечного нерва, идентифицированы только у аплизии, у которой они расположены в левом полуганглии абдоминального ганглия [233а, 239]. Наши данные о фармакологии передачи с моторных волокон тентакулярного нерва на ретрактор глазного щупальца у виноградной улитки, а также об ультраструктуре секреторных везикул в окончаниях этих волокон указывают на возможную холинергическую природу соответствующих мотонейронов, которые расположены в плевральной доле церебрального ганглия. Блокирующие эффекты холинолитиков отмечаются на моторных окончаниях других дериватов колумеллярной мышцы и самой этой мышцы, о чём уже говорилось. Холинергическую природу колумеллярных моторных нейронов нельзя, однако, считать строго доказанной; более того, имеются экспериментальные данные в пользу медиаторной функции глутамата в моторных окончаниях на одной из мышц этой системы (ретрактор глотки) [212] — эти данные в свою очередь подвергались серьёзной критике [см. 157, 166]. Моноаминергические нейроны. По крайней мере, существование дофаминергических и серотонинергических нейронов вряд ли может сейчас вызывать сомнение. По-видимому, этими двумя типами медиаторной специфичности разнообразие моноаминергических нервных клеток у гастропод не ограничивается, но в отношении других типов данных немного. Среди серотонинергических нейронов хорошо изучены только парные церебральные нейроны виноградной улитки (ПЦ1 и ЛЦ1), а также гомологичные им клетки тритонии и аплизии. Для этих клеток представлены хорошие доказательства тому, что их медиатором служит серотонин [131, 264]. Эксперименты на изолированных идентифицируемых нейронах виноградной улитки показали, что указанные клетки способны синтезировать серотонин, т. е. декарбоксилировать 5-окситриптофан, тогда как другие проверенные нейроны такой способностью не обладают [137, 259, 263]. Измеренное с помощью высокочувствительного микрометода содержание серотонина оказалось в парных церебральных нейронах тритонии и аплизии равным примерно 4 — 7 пМ на клетку (таково же содержание серотонина в дающих жёлтую люминесценцию педальных нейронах тритонии). В этих клетках измеримых количеств катехоламинов не оказалось [331, 240]. Синаптические эффекты серотонинсодержащих церебральных нейронов на клетки буккальных ганглиев и влияние этих нейронов на иннервированные их аксонами губные мышцы подобны эффектам апплицируемого серотонина [130, 266а]. Клетки, о которых идёт речь, обладают весьма специфичными системами захвата своего медиатора и его метаболического предшественника: сам серотонин захватывается терминальными участками аксона, а 5-окситриптофан — телом нейрона; контрольные нейроны, не содержащие серотонина, ни той, ни другой способностью не обладают и в их цитоплазме можно обнаружить лишь экзогенный триптофан [266]. Иннервационные отношения клеток ПЦ1 и ЛЦ1 указывают на то, что каждая из этих клеток сочетает функции интернейрона и мотонейрона [266а]. Выделение серотонина нейромоторными окончаниями показано у гастропод на сердечной мышце [298], где серотонин усиливает биения. Противоположное — расслабляющее действие серотонин оказывает на некоторые другие мышцы (например, ретрактор омматофора — собственные наблюдения); механизм такого эффекта давно изучается на ретракторе биссуса двустворчатого моллюска — мидии, где серотонин служит медиатором тормозных нервных окончаний [см. 166, 350]. На ультраструктурном уровне органеллы, содержащие серотонин, описываются в нейронах гастропод как везикулы диаметром около 1000 Å, содержащие центральное или эксцентричное плотное ядро; содержимое этих органелл избирательно выявляется при обработке бихроматом по методу Вуда [135, 266а, 331]. Преимущественным катехоламином нервной системы гастропод является дофамин [см. 166, 209, 334], и медиаторная функция этого вещества в ганглиях доказывается широкой системой критериев [83, 211, 306, 328]. Исследование идентифицируемого гигантского нейрона катушки, содержащего дофамин, дало окончательные свидетельства медиаторной роли этого катехоламина, который, подобно ацетилхолину в клетке Л10 аплизии, осуществляет как возбуждающие, так и тормозные синаптические эффекты [92]. Имеются все основания думать, что описанный в 5 главе гигантский непарный педальный нейрон прудовика, дающий зелёную люминесценцию, является гомологом названного интернейрона катушки; клетки этого ряда, как отмечалось в главе 5, имеются и у сидячеглазых пульмонат. Наши и литературные данные указывают на то, что дофаминергические нейроны гастропод могут выполнять также функции моторных и сензорных нейронов. О моторных окончаниях на некоторых клетках педальной мускулатуры выше уже говорилось. Такие окончания, по-видимому, имеются и в других мышечных органах моллюсков: в пищеводе (собственные наблюдения), сифоне [248 — данные для аплизии] и т. д. Помимо дофамина, в нервной системе гастропод отмечен, хотя и в меньших количествах, другой первичный катехоламин — норадреналин [260]. Гистохимический метод, позволяющий дифференцировать между дофамином и норадреналином, пока не прилагался к нейронам гастропод, но использование этого метода на других беспозвоночных (аннелиды, членистоногие) показало, что у них имеются как дофаминергические, так и норадренергические нейроны [149, 219]. Не исключено, что такова же ситуация у гастропод. Во всяком случае, эффекты этих двух катехоламинов на один и тот же нейрон улитки могут быть разными [170]. В экстрактах из нервной ткани разных моллюсков найдено несколько физиологически активных веществ, одно из которых имеет люминесцентные характеристики, присущие катехоламинам, не являясь в то же время ни дофамином, ни норадреналином, ни адреналином [73]. Вполне возможно, что эти находки указывают на существование неизвестного медиаторного амина. Имеется и другая сходная возможность: гигантские парные нейроны гастро-эзофагальных ганглиев тритонии проявляют, при обработке по методу Фалька и Хилларпа, жёлтое свечение, характерное для серотонина и других индольных соединений [37], микрохимические же исследования этих клеток показывают, что в них имеется неидентифицированное вещество, отличное от серотонина (Р. Мак-Каман, личное сообщение). Таким образом, кроме трёх известных медиаторных моноаминов, у гастропод, вполне возможно, существуют и другие вещества этой группы. Наконец, клетки, найденные у лёгочных улиток в составе висцеральной дуги, как сообщают Керкут и соавторы, содержат два медиаторных амина, дофамин и серотонин [213]. Это весьма необычная ситуация, и я специально вернусь к ней в разделе 6.7. Нейроны, имеющие медиаторами аминокислоты. У млекопитающих мы называли три таких медиатора: глицин, глутамат и гамма-аминомасляная кислота. Пока никто не исследовал, имеются ли у гастропод нейроны, медиатором которых служит глицин. Глицин присутствует в их нервной системе [258], а у самих нейронов имеются рецепторы к глицину, фамакологически похожие на соответствующие постсинаптические рецепторы позвоночных [255], но этих данных, конечно, недостаточно, чтобы утверждать, что у гастропод представлен этот тип синаптической передачи. Примерно такая же ситуация с гамма-аминомасляной кислотой. Нервная ткань гастропод отличается низким содержанием этого вещества [257]. Следовательно, нейронов этого типа может быть очень мало, а возможно, что их нет совсем. Если верно последнее, то моллюски отличаются в этом отношении не только от позвоночных, но и от таких беспозвоночных, как ракообразные и насекомые [см. 61]. Однако чувствительностью к гамма-аминомасляной кислоте нейроны гастропод обладают, при этом рецепторы блокируются пикротоксином и бикукуллином, как и у позвоночных [326]. Глутамат скорее всего служит медиатором у гастропод. Этот вывод на основании совокупности экспериментальных данных делают в специальном обзоре Керкут и соавторы [212].Дополнительно упомяну, что особое двухфазное синаптическое торможение клеток ПЦ1 и ЛЦ1 садовой улитки во всех деталях воспроизводится глутаматом [304]. Из этого не следует, что глутамат выступает исключительно в роли тормозного передатчика: одни нейроны отвечают на него торможением, другие — возбуждением [167, 256, 288]. До сих пор никто, однако, не пытался идентифицировать нейроны, секретирующие глутамат,- в частности, неизвестны клетки, оказывающие тормозные синаптические эффекты на ПЦ1и ЛЦ1. Кроме трёх названных аминокислот, можно упомянуть аргинин (или его производное), как будто высвобождающийся некоторыми нервными окончаниями улитки [298]. Пептидергические нейроны. Наши электронно-микроскопические данные, полученные на ЦНС виноградной улитки, показывают поразительное обилие и разнообразие нейронов, содержащих гранулы пептидергического типа. Об этом же говорят исследования, проведённые на переднежаберных улитках [79], а также на сидячеглазых пульмонатах [200, 336]. Во всех этих случаях наблюдается постоянство топографии клеток, содержащих гранулы определённой категории (рис. 15). Разные группы предположительно пептидергических нейронов, различающиеся по ультраструктуре секреторных гранул, выступают как в роли нейросекреторных клеток, так и в роли «обычных» нейронов, иннервируя, в частности, мышечные клетки оболочки ганглиев. У некоторых видов моллюсков такие нейроны картированы, но знания о строении секретируемых веществ пока что очень ограниченны. Ни в одном случае их идентификация не доведена до конца, но во всех изученных случаях эти вещества в самом деле оказались пептидами. С начала 60-х годов ведутся работы по выделению «субстанции X» — вещества, стимулирующего сердце и содержащегося в пептидергических гранулах соответствующих нервных волокон. Эти волокна подходят к сердцу в составе сердечного нерва не только у гастропод, но и у двустворок, где подсчитано, что они составляют около 5% волокон нерва. Отмечают, что активное начало представляет собой «большую и устойчивую молекулу» и что оно инактивируется протеолитическими ферментами; фракционированием выделены два активных вещества молекулярным весом около 700 — 1500 [см. обзор 333, а также 175]. Про нейросекреторную клетку ППа1 и её гомологи, синтезирующие полипептид м. в. около 5000, говорилось в разделе 5.3.2. Успешно развиваются исследования нейронов, выделяющих нейрогормон, который вызывает откладывание яиц у аплизии. Две группы таких клеток, по 400 нейронов в каждой, расположены вокруг плевро-висцеральных коннективов рядом с абдоминальным ганглием. Клетки электротонически связаны между собой и генерируют потенциалы действия синхронно, чем достигается одновременная секреция большого количества нейрогормона.

Рис. 15. Разнообразие пептидергических нейронов в ЦНС прудовика Lymnaea stagnalis [из 336]. А — схема окологлоточного ожерелья ганглиев (педальные комиссуры перерезаны и педальные ганглии отвёрнуты) и позиции клеток, различающихся по ультраструктуре секреторных гранул. Обозначение ганглиев: 1 — правый церебральный, 2 — левый церебральный, 3 — правый педальный, 4 — левый педальный, 5 -правый плевральный, 6 — левый плевральный, 7 — правый париетальный, 8 — левый париетальный, 9 — висцеральный.
 Рис. 15 (продолжение) Б — гистограммы диаметров секреторных гранул для некоторых групп нейронов из изображённых на рис. 15, А. Заштрихованные столбцы — гранулы в телах нейронов, светлые — гранулы в аксонных терминалях. Латинскими буквами обозначены сокращённые названия клеточных групп.
Рис. 15 (продолжение) Б — гистограммы диаметров секреторных гранул для некоторых групп нейронов из изображённых на рис. 15, А. Заштрихованные столбцы — гранулы в телах нейронов, светлые — гранулы в аксонных терминалях. Латинскими буквами обозначены сокращённые названия клеточных групп.
Показано, что он представляет собой полипептид молекулярным весом около 6000. Секреторные гранулы, имеющие типичную для пептидергических нейронов ультраструктуру, выделяются, по-видимому, в оболочку коннективов и абдоминального ганглия, где слепо заканчиваются аксоны этих нервных клеток. Выделившийся нейрогормон оказывает свой эффект, вызывая сокращение неиннервированных мышечных клеток протока гермафродитной железы [80, 81, 123, 125, 227, 315]. Значительное большинство пептидергических нейронов остаётся пока неисследованным ни в отношении функции, ни тем более в отношении строения активного начала. У прудовика различают 10 типов таких нейронов [200], но, на мой взгляд, эта цифра занижена. Нейроны с неизвестными медиаторами. Многие авторы, изучающие постсинаптические потенциалы в ганглиях виноградной улитки и аплизии, отмечали, что далеко не во всех случаях синаптические эффекты воспроизводятся каким-либо из известных или предполагаемых медиаторов. Очевидно, что медиаторные функции у моллюсков выполняются многими веществами неизвестной природы. Так, из интернейронов абдоминального ганглия аплизии только один [Л10] оказывает свои эффекты с помощью известного медиатора — ацетилхолина. Есть основания предполагать, что и в нервных окончаниях на эффекторных органах гастропод действует ряд пока что неизвестных медиаторных механизмов. Назовём только один пример, известный нам по собственному опыту работы. У свободноплавающей личинки — велигера голожаберных моллюсков имеются ярко выраженные тормозные нервные влияния на биения моторных ресничек паруса. Г. Н. Бузников и Б. Н. Манухин, детально исследовавшие это явление, нашли, что на той стадии развития, когда устанавливаются эти нервные влияния, в гомогенате личинок начинает обнаруживаться вещество, воспроизводящее этот эффект. Одновременно появляется вещество, воспроизводящее другой синаптический эффект — сокращение мышцы паруса. Ни одно из двух веществ не похоже ни на один из известных или предполагаемых медиаторов [см. 15, гл. 6]. Позже мы с Г. Н. Коробцовым снова пытались изучать механизм синаптических влияний на биения ресничек и, в частности, показали, что активное начало при хроматографировании экстракта в системе уксусная кислота: бутиловый спирт: вода (1: 4: 1) обнаруживается в области Rf = 0,9 и даёт пятно, которое не красится нингидрином [22]. Большего пока об этом неизвестном медиаторе сказать нельзя. Неизвестной остаётся также природа тех нервных окончаний, описанных в разделе 5.3.5., для которых характерна высокая активность дегидрогеназы глюкозо-6-фосфата. О наличии у моллюсков неизвестных медиаторов, возможно, свидетельствует и постоянное присутствие в их нервной ткани нескольких неидентифицированных физиологически активных веществ [73, 172]. Подытожив данные о химической (медиаторной) разнородности нейронной популяции гастропод, мы сравним их с соответствующими данными для млекопитающих, чтобы получить ответ на вопрос, зависит ли гетерогенность нейронной популяции от уровня нервной организации.
6. 2. У кого больше медиаторов?
Уровни нервной организации, представляемые млекопитающими и гастроподами, очень различны. Мощно развитая нервная система первых содержит, как считают, 109 - 1010 нейронов, у вторых их число в миллион раз меньше (103 - 104). Простым отношениям соподчинения, существующим в нервных центрах гастропод, противостоят сложнейшие иерархические системы нейронов мозга млекопитающих и, соответственно, высокая степень функциональной специализации нейронов и нервных центров, многоэтажные надстройки над рефлекторными дугами и т. д. Короче говоря, различия очевидны. Но при сравнении качественного состава нейронных популяций у организмов, относящихся к этим двум далеким друг от друга классам, разница вовсе не бросается в глаза. Хотя нейронная популяция у млекопитающих исследована несомненно с большей тщательностью, не может быть и речи о том, что у них разнокачественность нейронов представлена ярче, чем у гастропод. Скорее, можно говорить о противоположном общем впечатлении, а в отношении моторных нейронов это уже не впечатление, а непреложный факт. При всей фрагментарности и неполноте данных об иннервации мышц гастропод обращает на себя внимание необычайное (по сравнению с позвоночными) разнообразие типов моторных окончаний. О холинергическом и дофаминергическом моторных механизмах говорилось в предыдущих разделах. Серотонин выделяется в сердце виноградной улитки при раздражении интестинального нерва и стимулирует биения [298], этот же медиатор, по-видимому, выделяется в моторных окончаниях нейронов ПЦ1 и ЛЦ1 на некоторых губных мышцах [266а]. Мышечные клетки периневрия получают моторные окончания от нейронов пептидергического типа. В иннервации таких мышечных органов, как нога и сердце, участвует несколько типов моторных клеток. Имеющиеся сведения об иннервации сердца особенно интересны. Напомню, что обнаружено по крайней мере три типа постсинаптических потенциалов на клетках миокарда, и эти типы связаны с разными идентифицированными нейронами [239]. В медиации влияний экстракардиальных нервных волокон, вероятно, принимают участие ацетилхолин, серотонин, «субстанция X» [см. 333], но в сердце электронно-микроскопически выявляется не один тип пептидергических окончаний, а больше. Так, на ахатинидах показано, что пептидергические гранулы разные в нервных волокнах, приходящих в предсердие и желудочек [252]. Мы с Н. К. Остроумовой только в предсердии одной из улиток этого семейства (Achatina fulica) нашли два типа пептидергических окончаний с совершенно разными гранулами. Конечно, не исключено, что некоторые из волокон являются нейросекреторными и их секрет не действует на миокард, как это утверждается для заканчивающихся в предсердии аксонных ветвей клетки ППа1 [162б]. Короче говоря, в иннервации мышц моллюсков участвуют нейроны, относящиеся к большему числу специфических типов, чем у позвоночных. Хотя позвоночные изучены в этом отношении лучше моллюсков, до сих пор у них найдено всего три типа мотонейронов: холинергические, иннервирующие как скелетную так и висцеральную мускулатуру; адренергические, контролирующие различные висцеральные мышцы; и пуринергические, иннервирующие мышечные клетки кишечника и его дериватов. Между тем, с точки зрения гипотезы о плюрихимической примитивной нервной клетке, которая дала якобы начало разным химически специализированным типам нейронов, должно было бы быть наоборот: у млекопитающих с их высоко развитой нервной системой и мощными дифференцированными двигательными системами разнообразие медиаторов должно было бы быть более выраженным, чем у малоподвижных и простых гастропод. На деле в примитивной нервной системе моллюска, содержащей всего несколько тысяч нейронов, обнаруживается большее разнообразие химических типов мотонейронов, чем у высших млекопитающих с их миллиардами нервных клеток и высокодифференцированными структурами нервной и мышечной систем. Сравнение центральных нейронов, взятых в целом, также не дает преимущества ЦНС млекопитающих перед ганглиями гастропод в отношении разнородности популяции.
6. 3. Каковы отношения между медиаторной специфичностью и функциональной специализацией нейрона?
Тот факт, что гомологичные клетки разных гастропод проявляют один и тот же тип медиаторной специфичности, сам по себе недостаточен для отрицания связи между типом химизма и функцией. Ведь можно было бы думать, что гомологичные клетки выполняют одинаковую функцию, и в этом видеть объяснение однотипности их химизма. Такое рассуждение приложимо и к гомологичным нейронам позвоночных. Опровергнуть это объяснение было бы просто, если бы в ряду гомологичных нейронов, стойко сохраняющих свою медиаторную специфичность, наблюдались бы разные направления функциональной специализации. Думается, педальные катехоламиновые нейроны представляют собой именно такой сравнительный ряд. Однако имеющиеся в нашем распоряжении данные нельзя пока принимать в качестве решающих, так как о сензорной и моторной функциях этих клеток мы судим только на основании морфологических критериев; для большей уверенности нужны были бы и физиологические данные, которых пока недостаёт. Правда, имеются другие яркие примеры. В эволюции некоторых рыб часть скелетной мускулатуры полностью утратила свою прежнюю функцию, превратившись в электрические органы. Хорошо известно, что на медиаторной специфичности эффекторных нервных окончаний это радикальное изменение функции никак не сказалось — они остались холинергическими. Ещё удивительнее пример с нервной регуляцией молокоотделения. Известно, что ответственные за эту функцию мышечные (так называемые миоэпителиальные) клетки молочной железы не пришли в этот новый орган из старых мышечных систем, а развились заново из эпителия. У низших позвоночных эти эпителиальные клетки принимают участие в водно-солевом обмене, и их ионная проницаемость регулируется гипоталамическим нейрогормоном. Новые мышечные клетки стали выполнять совершенно иную функцию, но сокращаться их заставляет, меняя ионную проницаемость, гипоталамический нейрогормон — окситоцин. Имеет смысл расмотреть вопрос шире. Располагает ли вообще сравнительная физиология данными, которые указывали бы на существование зависимости между химической природой медиатора и его синаптическим эффектом? На первый взгляд, существование такой зависимости может показаться самоочевидным. Вероятно, не одному физиологу приходила в голову мысль, что неспроста быстрые скелетные мышцы запускаются в действие холинергическим механизмом, тогда как тонические сокращения гладких мышц контролируются адренергическими окончаниями. При этом привычно упускается из виду, что тот же самый синаптический ацетилхолин вызывает сокращения не только быстрых тетанических, но и медленных тонических скелетных мышц позвоночных, что он же на сердечную мышцу оказывает тормозные влияния, и т. п. В самом деле, сравнительные данные показывают, что один и тот же медиатор может обеспечить разнообразные синаптические эффекты, различающиеся по ионным механизмам, знаку синаптического действия, эффективности и физиологическим параметрам передачи. О разнообразии ионных эффектов ацетилхолина на постсинаптические мишени уже говорилось: этот медиатор может открывать ионные каналы для натрия, калия, хлора; кроме того, описан способ действия ацетилхолина, когда он не повышает, а понижает ионную проницаемость постсинаптической мембраны [224,329]. Избирательное повышение проницаемости для одного из названных выше ионов, а также избирательное понижение проницаемости для ионов натрия или калия лежит в основе замечательного разнообразия эффектов серотонина на нейроны гастропод [165, 166, 168, 264а; о разнообразии эффектов серотонина на клетки мозга млекопитающих см. 193]. Помимо этого, известны эффекты серотонина, при осуществлении которых он, по-видимому, действует внутриклеточно: таково его действие на биения моторных ресничек личинок моллюска [23], на ретракторную мышцу биссуса мидии [350]. Сходные факты, хотя и в меньшем объеме, можно представить для эффектов других медиаторных веществ. Ограничив клеточные мишени нейронами гастропод, можно найти данные о нескольких способах действия дофамина [83, 92], глутамата [167,256, 288], гамма-аминомасляной кислоты [12]. Глицин, который вместе с гамма-аминомасляной кислотой часто именуется тормозным медиатором, тоже способен деполяризовать некоторые нейроны гастропод [255]. Для синаптического норадреналина известны противоположные по знаку эффекты на гладкомышечные клетки позвоночных [например, 69, 84]. Короче говоря, не может быть сомнений в том, что знак и ионный механизм синаптического действия определяется молекулярным устройством воспринимающих структур постсинаптической мишени — клеточных рецепторов. Разные медиаторы могут вызывать сходные синаптические эффекты, действуя сходным образом на ионные каналы постсинаптической мембраны, и вместе с тем один и тот же медиатор может действовать по-разному. Всё это важно иметь в виду, потому что вопрос о факторах, обусловливающих знак синаптического действия, излагается путано или неверно даже весьма именитыми авторами. Так, Экклс в своей книге «Тормозные пути центральной нервной системы» [71]не только широко оперирует понятием «тормозной медиатор», но и устанавливает некий принцип, который гласит, что «во всех синаптических терминалях нервной клетки медиатор открывает ионные поры лишь одного типа, характерного либо для возбудительных, либо для тормозных синапсов» (стр. 145). Упоминая данные о том, что у моллюсков разные ветви одного интернейрона могут оказывать противоположные по знаку синаптические эффекты (например, [203, 301]), Экклс утверждает, что эти эффекты не противоречат сформулированному им «принципу», так как в данном случае и в возбудительных, и в тормозных синапсах открываются-де поры для ионов хлора, а эффекты различны из-за различных внутриклеточных концентраций хлора в постсинаптических клетках. Это утверждение не соответствует действительности. Холинергический интернейрон аплизии Л10 тормозит некоторые клетки, открывая поры для ионов хлора, возбуждает другие, повышая проницаемость для натрия, а в клетке Л7 повышает проницаемость для обоих этих ионов [96]. Приходится с сожалением констатировать, что многие авторитетные специалисты в области физиологии и морфологии нервной системы, внесшие заметный вклад в изучение химических синапсов, допускают подчас непростительную небрежность в обращении с химической концепцией синаптической передачи. Можно лишь удивляться, например, легковерию, с каким многие восприняли дилетантскую идею Учизоно [318], что знак синаптического действия можно идентифицировать по форме и размеру синаптических пузырьков (см., например, в той же книге Экклса, стр. 136). Ведь очевидно, что эта идея подразумевает невероятное — что молекулярная организация органелл секреции должна зависеть от того, с какой клеткой продукт секреции встретится во внеклеточном пространстве. Что касается упомянутого выше «принципа», то здесь необходимо отметить, что Экклс провозгласил его, практически не располагая экспериментальными фактами в его пользу. Ведь моллюски — единственные животные, у которых до сих пор удавалось регистрировать синаптические эффекты в разных постсинаптических клетках, иннервированных одним и тем же нейроном, и факты, полученные на моллюсках, противоречат «принципу» Экклса. Немногие факты, касающиеся млекопитающих, тоже не говорят в пользу этого «принципа». Так, терминальное сплетение норадренергических симпатических нейронов деполяризует мышечные клетки одних кровеносных сосудов, гиперполяризуя мышечные клетки других. Представляется крайне маловероятным, что одни участки сплетения образованы «возбуждающими» симпатическими нейронами, а другие «тормозными»,- скорее, это отростки одинаковых клеток, антагонистически действующие на разные мышечные рецепторы. Но детально это никем не исследовано. Итак, важнейшие функциональные характеристики синапса — знак и ионный механизм синаптического эффекта могут определяться особенностями структур, воспринимающих действие медиатора. Другие характеристики — такие, как размеры, форма и длительность постсинаптического потенциала, облегчение, десенситизация, пластичность и т. п., определяются не без участия самого пресинаптического нейрона, но, судя по всему, без какой-либо необходимости менять медиатор. Упомянем в этой связи моторные окончания на быстрых и медленных мышцах членистоногих: разная эффективность синаптического действия обеспечивается здесь при одном и том же медиаторе — глютамате различиями в размерах терминалей, числе везикул, числе активных зон синапса и т. п. [см. обзор 279]. В нескольких работах, совместных с Т. М. Турпаевым, нами детально рассмотрены некоторые физиологические механизмы, обеспечивающие изменение параметров передачи в холинергических синапсах, в частности, влияние способа инактивации ацетилхолина на свойства синапса (см. гл. 7). Нет и фатальной зависимости между типом медиаторного химизма и «макрофункцией» нейрона — его местом в рефлекторной дуге. Холинергические нейроны бывают и сензорными [86, 157], и вставочными, и эффекторными (примеры приводились выше). В тех же ролях выступают дофаминергические нейроны: сензорными они бывают у многих беспозвоночных, интернейроны хорошо известны у млекопитающих (см. главу 2) и гастропод [92], о существовании мотонейронов этого типа говорилось выше. Видимо, по мере накопления сравнительных данных наши знания о функциях, выполняемых нейронами того или иного химического типа, будут всё более расширяться. Они уже сейчас достаточно широки, чтобы окончательно отказаться от терминов «возбуждающий медиатор» и «тормозной медиатор». В свете всего сказанного в этом разделе предположение о том, что химический тип нейрона может задаваться функциональным прессом — специализацией ли нейрона или режимом функционирования синапса, представляется лишенным основания. Из этого, конечно, не следует, что за нейронами определенного химического типа в определенном нервном образовании не может закрепиться определенная функциональная специализация [см., например, 2а]. Напротив, наличное разнообразие медиаторов предоставляет удобную возможность дифференцировать сигналы разного назначения: так, в каких-то мышцах медиатор А может быть возбуждающим, а Б — тормозным, но в других мышцах А и Б могут поменяться ролями.
6. 4. Специфичен ли химизм примитивных нейронов?
Из представления о плюрихимической примитивной нервной клетке с неизбежностью вытекает, что в простой нервной системе химическая специфичность нейронов должна быть выражена не так четко, как в сложной, эволюционно продвинутой Факты говорят о противоположном. В разделе 6.1. приводились данные, говорящие о совершенной специфичности холинергических, серотонинергических, дофаминергических нейронов гастропод. Результаты микрохимических исследований на их гигантских нейронах получают полное подтверждение в этом отношении в электрофизиологических наблюдениях, которые однозначно показывают, что в том или ином синапсе действует индивидуальный передатчик, а не смесь таковых. Никаких данных, указывающих на меньшую, по сравнению с млекопитающими, степень медиаторной специфичности нейронов, нет и для других животных, обладающих простой нервной системой [см., например, обзоры 166, 279]. Даже у наиболее примитивных из организмов, имеющих центральную нервную систему, — у плоских червей (планарии и трематоды) разные группы нейронов закономерно различаются по своей специфичности. Об этом свидетельствуют данные люминесцентной гистохимии биогенных аминов: позиции и связи нейронов, содержащих серотонин, отличаются от позиций и связей нейронов, содержащих первичный катехоламин [89, 335]. У плоских червей описаны также системы пептидергических нейронов. Более того, специфичность нейронов четко выражена у кишечнополостных — организмов, не обладающих ещё нервными центрами. Нервную сеть кишечнополостных обычно рассматривают как наиболее примитивную из нервных систем, развившихся в ходе эволюции многоклеточных животных. До недавнего времени оставались неясными даже принципы строения сети, вернее двух сетей, независимо развивающихся в эктодермальном и энтодермальном слоях кишечнополостных. Ныне полностью опровергнута идея о синцитиальной организации сети: даже у наиболее примитивных представителей типа, как и у относительно сложных, электронно-микроскопически доказано наличие истинных химических синапсов между нейронами. При этом обнаружился замечательный факт: сеть всегда построена из нейронов разных типов, с разными ультраструктурными характеристиками секреторных везикул [341 — 343]. Выводы электронной микроскопии подтверждаются гистохимическими данными. Присутствие первичного катехоламина обнаружено у актиний в закономерно расположенной категории эктодермальных нейронов — в веретеновидных клетках, тела которых находятся в эпителии щупалец, а отростки, имеющие характерные для катехоламиновых нейронов варикозные пресинаптические утолщения, образуют субэпителиальное сплетение нервных волокон в самих щупальцах и вокруг рта. Авторы, описавшие эти клетки у некоторых актиний [140, 47], сочли их сензорными нейронами, принимая во внимание типичную для чувствительных клеток позицию в наружном эпителии. Я переисследовал этот вопрос на актинии Bunodactis stella и нашел явные синаптические контакты между указанными отростками и мышечными клетками эктодермального слоя мускулатуры щупалец [рис. 1 и 2 в работе 281]. По-видимому, эти клетки выполняют и сензорную и моторную функции. Это — примитивное свойство, свидетельствующее о низком уровне функциональной дифференциации клеток в нервной системе актинии. Но никаких признаков низкого уровня химической дифференцированности нейронной популяции не видно: специфичность катехоламиновых нейронов выражена у актиний так же отчетливо, как у млекопитающих. Кстати, примитивные мультифункциональные сензо-моторные нейроны, существование которых ещё недавно отрицалось [188], почти одновременно были найдены у трех разных организмов — и именно там, где такие нейроны следовало ожидать, т. е. в нервных образованиях с низким уровнем функциональной дифференциации. Кроме упомянутых только что нейронов актинии, такие клетки описаны у гидры, где они сочетают функции сензорных, моторных и вставочных нейронов [342а], а также в гениталиях аплизии [124]. Последний случай не должен удивлять. В целом гастроподы, конечно, намного сложнее кишечнополостных по уровню нервной организации, но если взять, периферические нервные сплетения гастропод, то они вряд ли намного сложнее, чем нервная сеть актинии или гидры. Это общее правило: даже у человека, имеющего высокоразвитый мозг, уровень функциональной организации некоторых отделов нервной системы весьма примитивен, и, скажем, интрамуральные ганглии кишечника недалеко ушли в этом отношении от ганглиев беспозвоночных. Возвращаясь ещё раз к мнению Т. Лентца о том, что комбинации нескольких медиаторов в одном нейроне есть отголосок прошлого [231], нужно сказать следующее. Лентц в этом мнении лишь предложил свое объяснение явлению, которое 10 лет назад считалось установленным фактом. В то время большинство нейробиологов принимало, что ацетилхолин имеется в симпатических норадренергических нейронах и гипоталамических пептидергических клетках и что он каким-то образом участвует в этих клетках в синаптической передаче. В частности, электронными микроскопистами в те годы описывались в окончаниях этих нейронов, помимо специфических гранул с электронно-плотным содержимым, скопления мелких прозрачных пузырьков, которые повсеместно считались вместилищем ацетилхолина и назывались «синаптическими везикулами» — в противовес более крупным «секреторным гранулам». В поддержку этих представлений привлекались данные о локализации холинэстеразы. Напомню, что чрезвычайно популярным было объяснение этих явлений, исходящее из предположения, что из любого окончания сначала секретируется ацетилхолин, который затем помогает выделиться «главному» медиатору — например, норадреналину [103]. Сейчас эти взгляды представляют только исторический интерес, так как вся их фактическая база сведена к нулю. Специальная проверка, проведённая с помощью разных методов, показала отсутствие измеримых количеств ацетилхолина в адренергических нервных окончаниях млекопитающих [128, 148]. Мелкие пустые гранулы окончаний этих клеток перестали быть пустыми, как только микроскописты научились с помощью тех или иных предосторожностей сберегать находящийся в них катехоламин. Также и в пептидергических окончаниях скопления пустых пузырьков получили естественное объяснение, когда выяснилось, что их число увеличивается по мере того, как «секреторные гранулы» теряют своё содержимое. Таким образом, у высших животных практически неизвестны нейроны с комбинациями медиаторов и нейрогормонов. Сравнительные данные показывают, что и у низших животных химическая специфичность нейронов выражена в полной мере.
6. 5. Един ли гистогенетический источник нервных клеток?
Существует несколько мнений о том, от каких клеток могли произойти примитивные нейроны, потомками которых являются клетки современных нервных систем. Чаще всего в качестве непосредственного источника нейронов называют эпителиальные клетки — мнение, восходящее к прошлому веку [3, 187, 188]. Имеются и другие точки зрения: известный биофизик Г. Грундфест считает предками нейронов железистые клетки, а Л. Пассано — мышечные [см. 231]. На мой взгляд, спора здесь нет, так как это взаимно дополняющие, а не исключающие мнения. В самом деле, легко найти аргументы в пользу каждого из трёх названных источников. Присутствие нейронов в составе разных эпителиев дает поддержку классическому взгляду. Вторая точка зрения может опереться не только на данные о цитофизиологических механизмах секреции, общих для железистых и нервных клеток, но и на возможное существование переходных форм клеток, примером которых могут служить так называемые «мелкие» клетки симпатических ганглиев млекопитающих, сочетающие, как считают, свойства железистых клеток хромаффинной ткани и нейронов. Труднее, казалось бы, иллюстрировать третью точку зрения, но и это возможно. Известно, что эволюция сердечной мышцы у позвоночных выражается всё увеличивающимся разделением функций между сократительными элементами и «специфической мускулатурой», принимающей на себя функцию генерации и проведения электрических импульсов. В ходе эволюции наблюдается прогрессивная утрата миофибрилл элементами специфической мышечной ткани. Если вообразить этот процесс продолжающимся, то легко представить, что миофибриллы будут утрачены полностью, и тогда по любому из цитологических или физиологических критериев получившиеся клетки нужно будет назвать нейронами. Одни из них будут выполнять пейсмекерную функцию, другие проводить возбуждение к сократимым элементам миокарда, получится сердце с нейрогенной автоматией. Вполне возможно, что эти этапы эволюции, воображаемые для сердца позвоночных, являются свершившимся фактом в историческом развитии сердца у членистоногих. Понятно, что эти спекуляции допустимы только в рамках представления о множественном происхождении нейронов. Если же исходить из точки зрения о едином филогистогенетическом источнике всех нейронов, то приходится каким-то двум из трёх предполагаемых предков давать отставку. Нужно, однако, уточнить, что представление о едином источнике нейронов может рассматриваться в качестве рабочей гипотезы лишь для ограниченного круга объектов. Претендуя на универсальность, оно вступило бы в непримиримые противоречия с известными фактами о независимом развитии эктодермальных, энтодермальных и иногда целомических нейронов в онтогенезе ряда беспозвоночных типов. Давно считается, что экто- и энтодермальные нервные сплетения независимо друг от друга развиваются у кишечнополостных [см., однако, 267]. Слабее это изучено для гребневиков, но и там возможна такая же ситуация. У низших вторичноротых (кишечножаберные, кишечнодышащие, иглокожие) тоже представлены два названных источника нервных сплетений, к которым добавляется третий — эпителий целома. Находки нейронов в эпителии средней кишки у аннелид, моллюсков и членистоногих указывают на то, что и у этих животных эктодерма служит гистогенетическим источником не для всех нейронов. Подробности и ссылки на оригинальные работы можно найти у В. Н. Беклемишева [3, гл. 3]. Любопытно, что в гистологии (зоологии это не касается) широко признаётся множественное происхождение различных специализированных клеток: мышечных, железистых и т. д., и лишь для нервных традиционно делается исключение. Наиболее глубоко, на сравнительной и экспериментальной основе вопрос о конвергентном развитии и, следовательно, независимом происхождении разных специальных тканей рассмотрен Н. Г. Хлопиным [66], который, однако, вслед за своими предшественниками считал, что к нейронам это не относится. Зная старые работы о развитии нейронов в производных разных эмбриональных пластов у беспозвоночных, Н. Г. Хлопин призывал переисследовать это явление и хотел думать, что нейроны появляются среди клеток неэктодермального происхождения в результате миграции из эктодермы. Единственным основанием для идеи о родстве всех нейронов, как это отмечал сам Хлопин, служило поразительное морфофизиологическое сходство всех нейронов. Ныне нам известно, что это впечатление сходства было результатом несовершенства методов исследования, существовавших в 30 — 40-х годах. На самом деле нейроны очень разные — качественно разные. Особенно важно, что качественно различны нейроны, развивающиеся в разных источниках. Так, катехоламиновые нейроны ни в одном зоологическом типе не встречаются в нейронных популяциях неэктодермального происхождения [см. обзор 279]. К сожалению, имеется очень мало исследований, из которых можно было бы извлечь данные о составе нейронных популяций разного происхождения. Наиболее удобным объектом таких исследований представляются иглокожие, и то немногое, что известно об их нейронах, ярко демонстрирует обусловленность специфического химизма происхождением. Как известно, у иглокожих сосуществуют, взаимодействуя, несколько нервных систем, различающихся по месту и источнику своего происхождения: эктоневральная, развивающаяся в эктодерме; энтодермальное сплетение, развивающееся из кишечного эпителия; гипоневральная и апикальная системы, являющиеся производными разных областей целомического эпителия (подробности см. у В. Н. Беклемишева [3]). Функция локомоции выполняется в разных классах иглокожих мышцами разного происхождения и, соответственно, находится под контролем разных нервных систем: апикальной у морских лилий, экто- и гипоневральной у представителей других классов. Другие функциональные отправления также выражены совершенно по-разному в разных классах иглокожих, столь непохожих друг на друга, что роднит их лишь общность плана строения. И тем не менее, у иглокожих наблюдается поразительное сходство в характере распределения нейронов специфических типов в разных по происхождению отделах нервной системы:
| Происхождение системы, в которой найдены | ||
|---|---|---|
| Класс | катехоламиновые нейроны | пептидергические нейроны |
| Морские лилии | нет данных | целомическое [185] |
| Морские звезды | эктодермальное [136, 178] | то же [178, 229] |
| Офиуры | то же [136] | » » [265] |
| Морские ежи | » » [121] | нет данных |
| Голотурии | » » [287] | то же |
Ограниченность сравнительных сведений о качественном составе нейронных популяций, различающихся по своему происхождению, не должна заслонять главного факта: нейроны в самом деле могут возникать независимо друг от друга. Никакого единого источника всех нейронов в реальности не существует. В какой степени сказанное относится к позвоночным, мы обсудим несколько позже (6.7).
6. 6. Трудности функционального объяснения
Вопросы, подлежащие проверке, были поставлены в такой форме, чтобы ответы, которые априорно даёт на них одна из гипотез, были противоположны тем, которые даёт другая. Сопоставляя эти априорные ответы с результатами исследований, мы вправе заключить, что трудности, с которыми сталкивается функциональная гипотеза, непреодолимы. В самом деле, она предсказывает, что разных медиаторов должно быть тем больше, чем выше уровень нервной организации. Это не соответствует реальной действительности. По функциональной гипотезе, определённому типу специфичности нейрона должна соответствовать определённая специализация нейрона и (или) синапса. Этого тоже нет. Ожидалось далее, что химические различия между нейронами, хорошо выраженные у высших животных, не должны быть столь же четкими у организмов, имеющих простую нервную систему. И это ожидание не оправдывается. Наконец, функциональная гипотеза подразумевает существование единого гистогенетического источника всех нейронов. В действительности же источники бывают разными.
6. 7. Трудности гипотезы полигенеза нейронов
Те же самые факты, которые не оставляют места для функциональной гипотезы, находятся в хорошем согласии с альтернативным представлением, предполагающим консервативность медиаторного химизма и сосуществование в нервной системе разных (по происхождению) линий нервных клеток. В рамках этого представления, т. е. гипотезы полигенеза, следует ждать, что: 1) у животных, сильно различающихся по уровню нервной организации, не должно быть заметных различий в разнообразии медиаторов; 2) нейроны, имеющие один и тот же тип химизма, могут сильно различаться в функциональном отношении; 3) химические различия между нейронами должны быть равно выраженными как в простых, так и в сложных нервных системах; 4) нервные клетки могут развиваться из разных тканевых источников. Однако мне не хотелось бы проходить мимо фактов, которые, по крайней мере на первый взгляд, могут показаться несовместимыми с данным представлением. Прежде всего обратимся к случаям, которые трактуются как примеры «комбинации медиаторов». Ещё несколько лет назад к этой категории явлений многие относили постулированное участие ацетилхолина в секреции норадреналина симпатическими нейронами и нейрогипофизарных нейрогормонов гипоталамическими пептидергическими нейронами; сейчас, однако, это участие вряд ли будет кем-нибудь из специалистов рассматриваться всерьез (см. об этом в разделе 6.4.). В нейробиологии беспозвоночных упоминания о «комбинации медиаторов» ещё сохранились, и наиболее известным примером служат моноаминергические клетки висцеральной дуги пульмонат, про которые утверждают, что в них представлены два медиатора — серотонин и дофамин. Единственное сравнимое явление известно для теплокровных, у которых симпатические нервные волокна в эпифизе содержат как норадреналин, так и серотонин. В этом случае природа явления известна, оно связано с некоторой неточностью механизма захвата аминов из внеклеточной среды. Содержащийся в паренхиме эпифиза серотонин благодаря захвату попадает внутрь симпатических волокон, где уже имеется присущий им норадреналин. Доступные экспериментальные данные о рассматриваемых висцеральных нейронах пульмонат указывают на возможность такого же происхождения «комбинации» и в этом случае. Нужно заметить, что эти данные очень невелики [213, 292]. На препаратах нервной системы садовой улитки, полученных по методу Фалька и Хилларпа, авторы отметили, что названные клетки имеют свечение жёлто-зелёного оттенка, которое становится зелёным, если улиткам ввести диоксифенилаланин (предшественник дофамина), и жёлтым, если им ввести 5-окситриптофан (предшественник серотонина). Не проводилось ни снятие спектров люминесценции, ни микрохимическое определение аминов в этих клетках; совсем неизвестны медиаторные механизмы их синаптических эффектов. В гистохимии биогенных аминов принято за правило не ограничиваться оценкой цвета люминесценции при окончательных суждениях о природе амина — такая оценка чревата ошибками [129]. В одной из цитированных работ [292] говорится, что люминесценция висцеральных нейронов улитки выцвечивается ультрафиолетом — свойство, присущее серотонинсодержащим клеткам, но отнюдь не тем, в которых содержатся катехоламины. Опыты с усилением свечения под действием предшественников малодоказательны: эти предшественники сами при обработке по методу Фалька и Хилларпа могут дать свечение. Известно также, что они декарбоксилируются у моллюсков одним и тем же ферментом, так что неточность механизма захвата предшественников из среды неминуемо может привести к появлению в цитоплазме медиаторного амина, который несвойствен самому нейрону. Я думаю, что для вывода о двух медиаторах, уживающихся в одном нейроне, имеющиеся данные совершенно недостаточны. С большей степенью вероятности можно думать, что в ганглиях висцеральной дуги у пульмонат имеется группа серотонинергических нейронов. Обратимся теперь к мотонейронам ракообразных. При микрохимическом исследовании одиночных клеток с известным действием на соматическую мускулатуру было найдено, что в тормозных и возбуждающих нейронах содержание глутамата примерно одинаково; тормозные отличаются от возбуждающих наличием значительного количества гамма-аминомасляной кислоты [см. обзор 279]. В данном случае комбинацию объяснить несложно. Прежде всего нужно подчеркнуть, что на самом деле никакой комбинации медиаторов здесь нет; как медиатор глутамат выделяется лишь в возбуждающих нейронах, синаптические же эффекты тормозных мотонейронов воспроизводятся гамма-аминомасляной кислотой, которая и является здесь единственным медиатором. Присутствие глутамата в тормозных нейронах имеет лишь косвенное отношение к синаптической функции: глутамат является метаболическим предшественником гамма-аминомасляной кислоты. О генеалогических отношениях между этими двумя типами нейронов см. также ниже (7.2.). Более трудной и неясной представляется ситуация в случае декарбоксилазы ароматических аминокислот. Медиаторные моноамины нейронов моллюсков — серотонин и дофамин образуются, как только что говорилось, в результате декарбоксилирования своих метаболических предшественников, 5-окситриптофана и диоксифенилаланина. В обоих случаях реакцию осуществляет один и тот же фермент, декарбоксилаза ароматических аминокислот (ДАА). Естественно было бы ожидать, что ДАА должна избирательно проявлять высокую активность в моноаминергических клетках. Проверка с помощью микрохимического метода, осуществлённая группой Р. Мак-Камана на отдельных нейронах тритонии и аплизии, показала, что, действительно, у тритонии в серотонинергических нейронах активность ДАА примерно в 500 раз выше, чем в контрольных нейронах. Однако на клетках аплизии результат получился иной: здесь ДАА в серотонинергических клетках лишь в 10 раз активнее, чем в остальных [330, 331]. Причина расхождения пока не найдена. Определённо, что оба вида представляют одинаковые уровни нервной организации, так что было бы неразумным связывать это расхождение с «эволюцией». Не исключено, что у аплизии ДАА может иметь высокую активность в глие, от которой практически невозможно очистить крупные нейроны, и тогда результаты стали бы понятными. Вообще, мне по собственному опыту известно, что межвидовые различия в ферментах глии у гастропод очень велики: так, холинэстераза имеется в сателлитной глие у одних видов и отсутствует у других. Наконец, хотелось бы специально рассмотреть вопрос об эктодермальном происхождении нервной системы позвоночных. Чаще всего обращают внимание на то, что гипотеза, объясняющая множественность медиаторных механизмов множественным происхождением нейронов, противоречит общепринятому взгляду, что все нейроны имеют единое эктодермальное происхождение. Вообще-то говоря, если бы гипотеза не противоречила общепринятым взглядам, то в ней не было бы никакой потребности. На протяжении всей этой книги я рассматриваю материалы, свидетельствующие об обоснованности предложенного объяснения и о невозможности объяснить множественность химических механизмов другим способом. Если это так, то в упомянутом противоречии, возможно, повинен общепринятый взгляд на происхождение нейронов позвоночных, который может оказаться неверным или неточно сформулированным. При более пристальном рассмотрении оказывается, что никакого общепринятого взгляда на эктодермальное происхождение всех нейронов не существует. В литературе имеется довольно много формул, различных в зависимости от того, насколько критически оценивают авторы фактическую сторону дела. Что касается фактической стороны, она заключается в следующем: 1) в онтогенезе позвоночных нервные клетки развиваются из эктодермальных закладок (спорным остается только онтогенетическое происхождение интрамуральных ганглиев, которые, по мнению многих, в том числе современных, авторов представляют в этом плане исключение [см. обзор этой дискуссии в 78]); 2) в онтогенезе многих групп беспозвоночных нейроны дифференцируются в составе как эктодермы, так и других клеточных пластов, о чём уже шла речь выше (6.5.). Если игнорировать факты, относящиеся ко многим беспозвоночным, и считать, что «онтогенез повторяет филогенез», то получается самая простая формула: все нейроны имеют эктодермальное происхождение, при этом «происхождение» трактуется расширительно, в историческом и онтогенетическом плане. Такая точка зрения вовсе не представляет редкости, она особенно характерна для учебных руководств, но порой попадает и в специальные издания. Например, в книге, дающей сравнительный обзор нейрогормонов и гормонов в животном царстве, утверждается, что у всех животных нейросекреторные клетки, «будучи нейронами», имеют эктодермальное происхождение [198]. В действительности, если взять, например, иглокожих, то у них пептидергические нейросекреторные клетки имеются только среди нейронов целомического происхождения. Более критические авторы уточняют, говоря об эктодермальном происхождении нейронов, что речь идёт о позвоночных. И, наконец, самые пунктуальные дают понять, что имеется в виду лишь онтогенетическое развитие нейронов у позвоночных, и в таком виде формула, возможно, соответствует фактам. За этой формулой встаёт интересное и во многом загадочное явление. В самом деле, почему в онтогенезе позвоночных все нейроны развиваются из эктодермальной закладки, если у беспозвоночных, в том числе и у вторичноротых, близких к предкам позвоночных, дело обстоит иначе? Думается, в анализе этого явления не отделаться заклинанием «онтогенез повторяет филогенез». Предположение, что прямые предки позвоночных существенно отличались от других низших многоклеточных, имея нейроны в составе только дериватов эктодермы, представляется маловероятным, — в таком случае из предков позвоночных пришлось бы исключить организмы, подобные кишечнополостным и низшим вторичноротым. Не исключено, конечно, что в процессе эволюции у предков позвоночных имела место утрата всех нейронов, кроме тех, которые произошли в эктодерме. Другая возможность — что здесь мы встречаемся с явлением мерогенеза, т. е. смещения разнородных закладок нервной системы в одну, эктодермальную. В онтогенезе позвоночных центральные и периферические нейроны происходят из нервной трубки (и плакод). Сама трубка является закладкой ЦНС — спинного и головного мозга. Нет сомнений в том, что история нервной системы не начинается с ЦНС, ЦНС возникает на относительно поздней стадии эволюции нервной системы и, следовательно, такой способ закладки нервной системы тоже является относительно поздним приобретением. Таким образом, онтогенетические данные мало что могут дать для понимания исторического прошлого нейронов позвоночных, по крайней мере, для понимания тех этапов их истории, которые предшествовали образованию ЦНС. Но если даже в самом деле оказалось бы, что у позвоночных сохранились лишь эктодермальные нейроны, то и в этом случае противоречия с гипотезой о множественном происхождении не было бы. Лёгкость, с какой нервные клетки возникают у низших животных в разных клеточных пластах, убеждает в том, что нейроны должны независимо друг от друга неоднократно появляться в ходе исторического развития разных производных одного и того же пласта. Я вовсе не утверждал и не утверждаю, что типов нейронов столько, сколько клеточных пластов. Напротив, нейронная популяция, развившаяся в пределах одного пласта (например, в эктодерме актиний или в дериватах целома морских звёзд), всегда неоднородна, и отсюда скорее всего следует, что такая популяция сама имеет множественное происхождение.
6. 8. Заключение
Результаты исследований, рассмотренных в предыдущих главах, легли в этой главе в основу обсуждения, к которому были также привлечены дополнительные литературные и отчасти собственные данные. В ходе обсуждения имеющиеся факты были сопоставлены с двумя гипотетическими схемами, описывающими возможный путь возникновения медиаторной специфичности нервных клеток. Обнаруживается, что гипотеза полигенеза, связывающая химическую разнородность нейронов с множественным и независимым их происхождением, хорошо соответствует фактам, тогда как альтернативная гипотеза, объясняющая множественность медиаторных механизмов различной специализацией нейронов, сталкивается со значительными, практически непреодолимыми трудностями. Вывод о том, что нервные клетки имели тот или иной тип химической специфичности с момента своего возникновения, означает, что явление специфичности не возникло в ходе эволюции нервной системы. Из этого, однако, не следует, что процесс функциональной эволюции, связанной с прогрессивным развитием нервной системы, не коснулся медиаторных механизмов. Напротив, правильное решение вопроса о происхождении медиаторной специфичности, является необходимой предпосылкой для понимания того, как шла эволюция медиаторных механизмов нервной системы. Этому вопросу будет посвящена следующая глава.
7. ЭВОЛЮЦИОННАЯ СТОРОНА ПРОБЛЕМЫ СПЕЦИФИЧНОСТИ НЕЙРОНОВ
7. 1. Методологические замечания
Хотя с позиций защищаемого в этой работе представления множественность механизмов химической передачи не появляется в процессе эволюционного совершенствования нервной системы, из этого не следует, что эволюционный процесс обошёл стороной синаптические механизмы и химическую специфичность нейронов. Напротив, именно представление о множественном происхождении нейронов и консерватизме медиаторного химизма открывает, на мой взгляд, возможность изучать действительные закономерности эволюции нейронов и синапсов, а не явления, ошибочно принимаемые за эволюционные. Поясню, в чём важность верного решения вопроса о происхождении нейронов для изучения их эволюции. Хорошо известно, что суждения, касающиеся эволюции биологических структур (органов, тканей, клеток и т. д.), а также эволюции их функций, складываются при анализе сравнительных данных. Однако не любые сравнительные данные пригодны для сравнения. Опыт эволюционной биологии утверждает, что объекты, составляющие сравнительный ряд, должны быть гомологичными: только в этом случае они сравнимы и могут дать материал для эволюционных суждений. Так, например, изучая крыловой аппарат в ряду насекомых, можно понять, как происходило эволюционное совершенствование механизмов полёта. Эволюцию механизмов полёта можно реконструировать и из сравнения птиц, но это будут совсем другие механизмы и другая эволюция. Исследователь совершил бы ошибку, пытаясь понять, как совершенствовались механизмы полёта, из сравнения чайки со стрекозой или, допустим, с белкой-летягой. Эти особенности методологии эволюционных исследований необходимо иметь в виду, обращаясь к эволюционной стороне проблемы химической специфичности нейронов и синапсов. Поскольку нейроны разнородны, нельзя делать эволюционные выводы, сравнивая произвольно взятые нейроны и синапсы разных животных, а также сравнивая клетки, сходные в функциональном отношении. Какие же нейроны и синапсы сравнимы? В общем виде ответ на этот вопрос представляется нам таким: сравнимы те, которые относятся к одному типу химической специфичности, — в каждом типе совершенствование синаптических механизмов выражалось по-своему. Гораздо сложнее конкретизировать объём того или иного типа химизма: представляют ли собой нейроны, секретирующие медиатор А, особый тип или же они имеют общее происхождение с нейронами, секретирующими медиатор Б? Этот непростой комплекс вопросов составляет особую задачу — задачу построения естественной системы нервных клеток.
7. 2. Эволюция медиаторов и естественная система нервных клеток
Нет сомнений в том, что в ходе эволюции имели место изменения строения медиаторов, чему есть прямые свидетельства. Из этого следует, что нейроны общего происхождения не обязательно имеют одинаковые медиаторы, хотя «глубинный» тип химизма у них должен быть одинаковым. Если нервные клетки систематизировать на генеалогических основаниях, т. е. строить естественную систему нейронов, то в такой системе родственные нейроны должны, очевидно, группироваться гнёздами — семействами, в которых каждый член характеризуется особым видом медиатора. Исследовав генеалогические отношения между членами семейства, можно выяснить, как в ходе эволюции менялась структура медиаторного вещества (оговорю ещё раз, что, когда речь идёт о медиаторной специфичности, имеются в виду и медиаторы, и нейрогормоны — см. 2.1.). В принципе, в этой идее нет ничего нового. В нейробиологической литературе уже высказывалось соображение, что три медиаторных катехоламина являются результатом диверсификации одного исходного типа химизма. Кажется, только Пшайдт интересовался вопросом о генеалогии катехоламинового семейства нервных клеток. Исходя из несколько произвольной посылки, что в процессе эволюции должно происходить укорочение цепи реакций, связанных с синтезом медиатора, Пшайдт в качестве исходного медиатора принимает адреналин, полагая норадреналин следующим членом ряда и дофамин — позднейшим продуктом эволюции [273]. С этой схемой определённо не согласуются данные сравнительной физиологии. У беспозвоночных, в том числе у кишечнополостных, вкатехоламиновых нейронах обнаруживаются, как правило, первичные катехоламины [47, 132, 140, 166, 209, 279, 287, 334]. В некоторых (пока немногих) случаях, когда это специально исследовали, оказалось, что дофамин и норадреналин выступают в качестве медиаторов в разных нейронах [149, 219]. В современной литературе мне известна только одна работа, в которой у беспозвоночных (один из видов аннелид) найден в нервной системе адреналин [237а]. Адреналин, по-видимому, играет роль медиатора в периферической нервной системе амфибий, в тех клетках, гомологи которых у всех других позвоночных, в том числе и у рыб, используют в качестве медиатора норадреналин. Вряд ли можно сомневаться, что адреналиновые нейроны возникли из норадреналиновых. Создаётся, таким образом, впечатление, что в эволюции катехоламинового семейства нейронов имело место не укорочение, а удлинение цепи ферментативных реакций синтеза передатчика. Нет, однако, оснований полагать, что это должно быть общим правилом. Начальные этапы эволюции катехоламиновых нейронов остаются неясными, и данных для их реконструкции пока нехватает, хотя существование этих нейронов прослеживается от кишечнополостных. Ранее я уже обращал внимание на то, что у организмов, имеющих нейроны не только эктодермального происхождения, катехоламиновые нейроны закономерно обнаруживаются лишь среди эктодермальных нейронов, чем косвенно подтверждается гомология этих клеток у животных, относящихся к разным зоологическим типам [279]. Учитывая позицию катехоламиновых нейронов у кишечнополостных (см. 6.4.), можно предполагать, что история этого семейства нейронов имела исходным пунктом циркумбластопоральную нервную сеть примитивных предков многих типов многоклеточных, но это суждение носит лишь самый предварительный характер. По сравнению с семейством катехоламиновых нейронов, более ясными представляются генеалогические отношения в семействе нейронов, секретирующих октапептиды. Сравнительными данными прочно установлено, что все мутантные формы как окситоцина, так и вазопрессина ведут начало от общего предкового вещества. Предполагают, что помимо мутаций, связанных с заменой той или иной аминокислоты в молекуле октапептида, здесь в процессе эволюции имела место дупликация гена, что привело к появлению двух родственных веществ, которые приобрели разную специализацию. Этапы эволюции этой линии нервных клеток хорошо прослежены в ряду позвоночных [см. ссылки в 67]. К сожалению, не было серьёзных попыток найти гомологичные клетки у современных беспозвоночных, среди которых многие формы имеют нейроны с такой же гистохимией и ультраструктурой, какие характерны для клеток, продуцирующих нейрогипофизарные нейрогормоны. Насколько мне известно, не было попыток определить, обусловлены ли эти сходства тем, что в соответствующих нейронах беспозвоночных имеются специфические белки — нейрофизины. Использование меченых антител к нейрофизинам позвоночных сделало бы такую задачу вполне выполнимой. Нейронами, секретирующими катехоламины и октапептиды, ограничивается список клеток, в отношении которых в литературе обсуждались вопросы генеалогии. Действительно, в этих случаях родственные отношения между нейронами бросаются в глаза. Но необходимо обсуждать и выяснять генеалогические отношения также и тех нейронов, для которых эти отношения не самоочевидны, в противном случае трудно и даже невозможно заниматься эволюцией синаптических механизмов. В этом отношении предстоит трудная работа, начинать которую нужно несмотря на то, что на медиаторной карте человека и других животных остаётся ещё много белых пятен. В первую очередь следует исследовать те случаи, где имеются более или менее ясные указания на возможность родства. Например, вполне вероятно родство между нейронами, секретирующими глутаминовую и гамма-аминомасляную кислоту. Первая из кислот является непосредственным метаболическим предшественником для второй. У членистоногих, где оба медиатора часто действуют на одни и те же мышечные клетки, гамма-аминомасляная кислота всегда выполняет роль тормозного агента, как бы дополнительного к главному, пусковому агенту — глутамату. Вторичный характер этих тормозных влияний выражается и в том, что чувствительность к глутамату присуща самим мышечным клеткам и сохраняется у них после денервации, тогда как чувствительность к гамма-аминомасляной кислоте устанавливается лишь при контакте с тормозными нервными окончаниями и при денервации исчезает. Все эти факты — указание на вероятное происхождение нейронов, секретирующих гамма-аминомасляную кислоту, от нейронов, секретирующих глутамат. Но возможно, что и ацетилхолин — член того же семейства медиаторов. Синаптические везикулы в холинергических и глутаматергических окончаниях очень сходны; известно, далее, что синтез ацетилхолина идёт (по крайней мере отчасти) через глутаминовую кислоту [225]. В этой связи интересны данные венгерских исследователей, которые нашли, что в митохондриях пресинаптических окончаний цикл Кребса видоизменён и направлен на продукцию глутамата [177]. Если бы оказалось, что такие митохондрии характерны для окончаний, секретирующих глутамат, ацетилхолин и гамма-аминомасляную кислоту, но не характерны, допустим, для катехоламиновых аксонных окончаний, это было бы сильным доводом в пользу сделанного сейчас предположения о родственных отношениях холинергических нейронов. В идеале изучение истории нейронов должно прояснить как генеалогические отношения внутри каждого семейства, так и источники происхождения разных семейств нервных клеток. Такие знания в совокупности позволили бы построить естественную систему нейронов. Вопросы, связанные с построением естественной системы животных тканей, исчерпывающе обсуждаются в книге Н. Г. Хлопина, много сделавшего для развития этой проблемы [66]. Хотя сам Хлопин считал, что нервная ткань, в отличие от других животных тканей, однородна, развитые и аргументированные им теоретические положения приложимы к нервной ткани и должны найти применение при построении естественной системы нейронов. Такая система, несомненно, будет отличаться от любой из существующих классификаций нейронов. Существующие классификации не учитывают естественных, реально действовавших в природе причин различий между нейронами, а берут за основу тот или иной произвольно взятый признак. Так, нередко нейроны классифицируют в зависимости от того, к какому животному они относятся («нейроны человека», «нейроны рыб», «нейроны моллюсков»). В других случаях клетки делят по их месту в рефлекторной дуге, по их размерам, по числу и длине отростков и т. д. Общий недостаток искусственных систем в том, что они почти ничего не сообщают о нейроне, кроме названного признака. Зная про клетку, что это «нейрон моллюска», мы практически ничего больше не можем про неё сказать. Деление нейронов в зависимости от их медиаторов ближе всего к естественной системе, но в существующем сейчас виде и такое деление во многом искусственно. Так, серотонин произвольно объединяют с катехоламинами в группу «биогенных аминов»; в группу нейронов с аминокислотными медиаторами попадают, по-видимому, совершенно неродственные клетки; вместе с тем родственные нейроны могут оказываться в разных группах. Преимущество естественной системы в том, что она даёт исследователю априорное знание свойств клеток. Я уже приводил пример из недавней практики, когда нейрон с нужными свойствами был найден у аплизии благодаря тому, что авторы опирались на понимание клеточных гомологий (5.3.1). Естественная система широко открыла бы возможности такого рода.
7. 3. Совершенствование механизмов передачи
Эволюция самих медиаторных веществ, рассмотренная в предыдущем разделе, представляет лишь одну сторону эволюции химических синапсов. Другая сторона — прогрессивные изменения химизма, не сопровождающиеся изменениями в строении медиатора. Здесь уже не раз говорилось, что синапсы, работающие при посредстве одного и того же медиатора, могут проявлять совершенно разные функциональные свойства (6.3). Теперь нужно специально обратить внимание на то, что некоторые из этих свойств являются достижениями эволюции. Так, например, долгие годы и даже десятилетия считалось, что присутствие ацетилхолинэстеразы в области синаптического контакта — это непременное свойство холинергических синапсов. Однако оказалось, что это свойство присуще далеко не всем холинергическим синапсам и достигается ими в процессе эволюции, причём в разных линиях эволюции многоклеточных функциональная задача инактивации синаптического ацетилхолина решается по-разному. Если обратиться, например, к ганглиям гастропод, в которых имеются настоящие холинергические интернейроны и, следовательно, холинергические синапсы, то оказывается, что здесь зачастую ещё не решена задача обеспечения холинергических контактов локальным ферментом, гидролизующим ацетилхолин. При попытке локализовать такую активность на срезах центральных ганглиев у трёх видов лёгочных улиток мы получили следующие результаты. У наземной улитки Caucasotachea фермент выявлялся в нервных стволах, отходящих от ганглиев, и в трактах, продолжающих эти стволы, т. е. не в синаптической области. У другой наземной улитки, Helix lucorum, фермент не выявлялся нигде и лишь при очень длительной инкубации небольшая активность отмечалась в нейропиле (её, вероятно, наблюдали у близкого вида Керкут и соавторы при попытке выявить фермент на уровне электронного микроскопа [249]). Наконец, у прудовика Lymnaea активность обнаруживалась вокруг тел нейронов, в сателлитной глие, т. е. и на этот раз вне синаптической области [285]. В полном совпадении с этими наблюдениями находятся электрофизиологические данные о том, что ингибиторы холинэстеразы не вызывают у гастропод удлинения холинергических постсинаптических потенциалов [309]. Те и другие данные дают основание считать, что синаптическое действие медиаторного ацетилхолина прекращается в указанных синапсах гастропод не вследствие гидролиза ацетилхолина холинэстеразой, а по другой причине. Здесь мыслимы два примитивных механизма. Во-первых, диффузия ацетилхолина во внеклеточное пространство. Известно, что у гастропод оболочка ганглиев не является барьером для макромолекул гемолимфы, омывающей ганглий. В гемолимфе же имеется значительная холинэстеразная активность [64]. Следовательно, медиатор, диффундирующий от области контакта, может быть инактивирован во внеклеточном пространстве. Достаточен ли этот механизм для эффективной синаптической передачи? Имеются расчёты, которые показывают, что в синаптическом контакте диаметром 2 мк и шириной щели 200 Å (а это близко к реальным размерам) благодаря диффузии количество ацетилхолина в щели должноуменьшаться вдвое каждые 0,15 мсек [см. ссылки в 317]. Такая скорость уменьшения концентрации ацетилхолина у рецепторов может обеспечить весьма быстрые постсинаптические потенциалы. Имеется и другой механизм, впервые обнаруженный на сердце двустворчатого моллюска [57] и затем подробно исследованный на этом объекте С. Н. Нистратовой [40, 41]. В этом случае синаптическое действие ацетилхолина прекращается благодаря тому, что вслед за ацетилхолином в синаптической области появляется вещество, снижающее сродство мышечных холинорецепторов к ацетилхолину, благодаря чему постсинаптическая структура теряет чувствительность к медиатору. Результаты, полученные на сердце моллюска и других нейро-эффекторных системах, послужили Т. М. Турпаеву основанием для развития представления о саморегуляции медиаторного процесса механизмом обратной связи [63]. В отличие от первого способа (диффузия), когда холинорецепторы после ухода ацетилхолина готовы к приёму новой порции медиатора, при втором способе (выделение антагониста) прекращение синаптического действия ацетилхолина сопровождается изменением рецепторов, их десенситизацией. Какой из двух примитивных способов действует в ганглиях гастропод, где нет ещё механизма ферментативной инактивации ацетилхолина в синаптической щели? По-видимому, оба. Литературные данные и наш собственный опыт показывают, что в ганглиях моллюсков имеются холинореактивные структуры, реагирующие на ацетилхолин без десенситизации (как должно быть при простой диффузии) и с выраженной десенситизацией (как должно быть при инактивации рецепторов или при сочетании её с диффузией). Нужно, однако, заметить, что в центральных холинергических синапсах моллюсков не обязательно должен действовать именно тот механизм десенситизации, который найден у моллюсков в сердце. Итак, обеспечение холинергических синапсов ферментом, гидролизующим ацетилхолин, представляет собой относительно позднее приобретение. Так, среди моллюсков этим механизмом в полной мере, по-видимому, обеспечены головоногие, у которых в некоторых чётко локализованных областях синаптического нейропиля выявляется высокая активность фермента, гидролизующего ацетилхолин. Здесь имеется одна крайне интересная деталь: в двух разных линиях эволюции головоногих — у октопод и декапод для гидролиза ацетилхолина в синаптических структурах нервной системы используются разные ферменты — факт, на который я в своё время обратил внимание при гистохимическом исследовании и который нашёл подтверждение при изучении природы этих ферментов [см. ссылки в 279]. Этот факт, возможно, указывает на то, что у общих предков двух групп головоногих холинергические синапсы были ещё не обеспечены гидролизующим ферментом и что это усовершенствование в каждом из двух случаев было изобретено самостоятельно, вследствие чего и ферменты, привлечённые к выполнению этой функции, оказались разными. Эволюция механизмов, посредством которых прекращается действие ацетилхолина, выброшенного нервным окончанием в синаптическую щель,- это только один из аспектов эволюции холинергических синапсов, и этот аспект был взят мной в качестве примера лишь потому, что здесь имеется собственный опыт работы. Другими авторами исследована и прослежена эволюция других аспектов передачи. Так, большой сравнительно-фармакологический материал позволил М. Я. Михельсону и его сотрудникам анализировать вопрос о молекулярной эволюции холинорецепторов [39]. Н. А. Вержбинская с сотрудниками пришли к интересным заключениям о разных способах прогрессивной интенсификации функции ацетилхолинэстеразы в эволюции нервных клеток у водных и наземных позвоночных [9]. Можно обсуждать вопросы об эволюции механизмов секреции ацетилхолина, об эволюционных причинах и следствиях появления диффузионных барьеров для ацетилхолина в синаптической щели, и многие другие. Такие же многообразные проявления эволюции можно найти в синапсах, где действуют другие медиаторы.
7. 4. Неравноценность медиаторов и отбор нейронов
Внеоднородной нейронной популяции клетки с разными типами химизма вряд ли имеют одинаковые шансы на успех в эволюции. Это должно создавать предпосылки к процветанию одних типов медиаторного химизма и уменьшению числа, вплоть до полного исчезновения, клеток с относительно неудачным химизмом, т. е. к отбору нейронов. Различие в шансах на успех, по-видимому, выявляется в ходе прогрессивной эволюции нервной системы по мере того, как всё более усложняются функциональные задачи, встающие перед нейроном и синапсом. Активный образ жизни — непременное условие прогресса нервной системы нуждается в синапсах, которые способны длительно и без искажения передавать сигналы с высокой частотой. Для этого от аксонной терминали требуется способность создавать запасы медиатора и быстро восстанавливать их, когда медиатор расходуется на производство синаптического эффекта. Одновременно медиатор должен обладать свойствами, позволяющими ему мгновенно исчезать из синаптической щели, лишь только он произвел своё действие. При столкновении с этими задачами сами собой обнаруживались различия в перспективности веществ, оказавшихся в роли медиаторов. Другим испытанием для нейронов должно было стать увеличение размеров животных, которое также закономерно сопряжено с их прогрессивной эволюцией. Увеличение размеров тела животного сопровождалось увеличением протяжённости отростков нервных клеток, что было связано с дополнительной нагрузкой на клеточные аппараты синтеза. Далее, потребность в повышении скорости проведения влекла за собой утолщение аксонов (механизм сальтаторного проведения — довольно позднее эволюционное приобретение, он представлен только у позвоночных), а утолщение аксонов в свою очередь требовало интенсификации белкового синтеза. Если принять во внимание, что нейроны — это клетки с особенно активным метаболизмом и напряжённым белковым синтезом [5], то становится понятным, что такие дополнительные нагрузки легче переносились клетками, у которых синтез медиаторов требовал меньших энергетических и метаболических затрат. По-видимому, в первую очередь должны были страдать нейроны с тяжелыми медиаторными молекулами, в частности, нейроны, секретирующие пептиды. Известно, что у нейронов с низкомолекулярными медиаторами лишь небольшая доля медиатора доставляется в секреторные терминали из тела клетки, в основном же аксонное окончание само покрывает свои потребности в медиаторе путем местного синтеза или реутилизации медиаторных молекул, захватываемых терминалью из внеклеточной среды. Напротив, пептидные медиаторные молекулы преимущественно, если не целиком, производятся в околоядерной области нейрона и затем транспортируются по аксону к секреторным окончаниям. Клетке с такими свойствами гораздо труднее длительно поддерживать высокую активность, чем, допустим, симпатическому нейрону, аксонные окончания которого захватывают катехоламин из внеклеточной среды. С другой стороны, трудно обеспечить высокую лабильность передачи, осуществляемой тяжелыми пептидными молекулами, трудно освободить от этих молекул синаптическую щель, и т. д. Доступные сравнительные данные показывают, что действительно в ходе прогрессивной эволюции нервной системы уменьшается относительная доля пептидергических нейронов в смешанной нейронной популяции; другая эволюционная тенденция выражается в том, что секретируемые пептиды теряют медиаторную функцию, становясь нейрогормонами. Так, у млекопитающих распространение пептидергических нейронов ограничено, по всей вероятности, подбугровой областью головного мозга, где такие нейроны функционируют преимущественно в качестве нейросекреторных клеток. У рыб в дополнение к этому имеется хорошо развитая система пептидергических нейронов в спинном мозге. Что касается гипоталамических пептидергических нейронов, то они у низших позвоночных нередко секретируют своё активное начало непосредственно на клетку-мишень, т. е. не выступают в роли нейросекреторных клеток. У организмов, представляющих относительно низкие уровни нервной организации, пептидергические нейроны широко распространены, разнообразны и нередко осуществляют прямую иннервацию клеток-мишеней. Разобранные нами гастроподы являются хорошим, но далеко не единственным примером — разнообразные нейро-эффекторные окончания пептидергической природы широко представлены и у насекомых [163, 270]. Любопытно, что у головоногих, ушедших далеко вперёд по сравнению с другими моллюсками по числу нейронов и сложности нервных центров, в центральном нейропиле волокна с секреторными гранулами пептидергических типов занимают весьма скромное место по сравнению с тем, что известно для гастропод. Зато в мозге головоногих очень высока концентрация ацетилхолина и велико число волокон, идентифицируемых как холинергические [см. ссылки в 279]. Если у гастропод встречаются лишь единичные холинергические нейроны [169, 241, 241а], то у головоногих наблюдается мощный расцвет клеток этого типа. Нет сомнения, что этот расцвет связан с эволюционной перспективностью холинергических нейронов. Очевидно, что не только в типе моллюсков, но и в других линиях эволюции многоклеточных животных холинергические нейроны демонстрируют свой прогрессивный характер, становясь одним из доминирующих типов. Для позвоночных мы не располагаем знаниями об эволюционных изменениях относительной численности холинергических нейронов, но зато сравнительной физиологией получены хорошие свидетельства их территориальной экспансии: в ходе эволюции позвоночных холинергические нейроны распространили свои влияния на области, которые ранее не относились к сфере их иннервации. Это на ряде примеров убедительно показывает Дж. Бурнсток в своём обзоре, касающемся эволюции автономной нервной системы позвоночных. Экспансию здесь проявляют не только холинергические, но и адренергические нейроны: любопытно, что в разных классах позвоночных независимо наблюдается выселение адренергических нейронов в висцеральные органы с образованием новых, ранее отсутствовавших ганглиев [104]. В отличие от периферической нервной системы, в ЦНС позвоночных наблюдается не экспансия моноаминергических нейронов, а противоположный процесс: катехоламиновые нейроны исчезают в процессе эволюции из спинного мозга, уменьшается представительство и область распространения серотонинергических нервных клеток, полностью исключаются нейроны хромаффинного типа, хорошо представленные в головном мозге круглоротых и имеющиеся ещё у некоторых других низших позвоночных [88]. Молодые формации мозга — такие, как кора большого мозга и кора мозжечка, возникают благодаря мощной пролиферации нервных клеток, относящихся к каким-то иным и, возможно, немногочисленным химическим типам. Очевидно, что далеко не все типы медиаторного химизма, представленные в древних, стволовых отделах мозга, используются при формировании нейронных популяций молодых его отделов. Было бы интересно попытаться понять, какими конкретно преимуществами располагают нейроны, получившие прописку в новых формациях и ставшие благодаря этому одними из самых многочисленных и важных в организме,- в частности, клетки, секретирующие гаммааминомасляную и, как думают многие, глутаминовую кислоту.
7. 5. Вопросы, встающие на очередь
Кажется, считается общим правилом, что попытка дать ответ на какой-то вопрос оборачивается в науке постановкой новых вопросов. В нашем случае это определённо так, и гипотеза полигенеза, объясняя разнокачественность наших нейронов консервативностью их химизма, с неизбежностью вынуждает задаться вопросом о причине исходных различий химизма у нейронов, относящихся к разным (по происхождению) клеточным линиям. Думаю, что этот вопрос обсуждать ещё рано, и ограничусь небольшими замечаниями. С одной стороны, кажется, что функцию передачи сигнала от клетки к клетке способно в принципе выполнять любое вещество. Здесь нет фатальной зависимости от строения молекулы — той зависимости, вследствие которой, например, мышечные клетки, имеющие разное происхождение, конвергентно приходят к одинаковой химической основе своей сократительной функции. У нервных клеток разного происхождения шансы на совпадение специфического химизма, казалось бы, незначительны: каждая клетка имеет бесчисленное множество разных метаболитов и каждый из них имеет шансы стать сигнальной молекулой, передающей информацию от данной клетки к другой. И в самом деле, среди веществ, передающих такую информацию, т. е. среди медиаторов и гормонов, мы находим представителей самых разных классов химических соединений — от нуклеотидов до стероидов, от одиночных аминокислот до крупных полипептидов. Но, может быть, такое рассуждение не совсем верно. X. С. Коштоянцем уже давно высказывалась мысль, что медиаторами становились те вещества, которые и до возникновения нервной системы выполняли регуляторные функции: самый акт синаптической передачи Коштоянц понимал как процесс, при котором медиатор — продукт метаболизма нервной клетки активно вмешивается в функциональный метаболизм иннервированной клетки [31, 33]. Эта «энзимо-химическая гипотеза», заслонённая в течение четверти века фактами, которые, как казалось, ограничивают синаптическое действие медиатора поверхностью клеточной мембраны, сейчас вновь становится актуальной в связи с обнаружением явлений, когда синаптический передатчик безусловно действует не с поверхности, а «по Коштоянцу». Если всё это так, то на роль медиаторов могли претендовать не все многочисленные метаболиты, а лишь те относительно немногие, которые занимали ключевые позиции в регуляции внутриклеточного обмена. Напомню, что в свое время именно с этих позиций Г. А. Бузников и Б. Н. Манухин рассматривали смену функций медиаторных веществ в онтогенезе: от регуляции дробления к функции локального гормона и затем от неё к функции синаптического передатчика [см. 6]. Здесь нелишне сослаться также на обоснованную экспериментальными данными мысль Н. Н. Демина, что эффекты медиаторных веществ на поверхностную клеточную мембрану есть, может быть, лишь частный случай их мембранных эффектов [17]. Сокращение списка веществ, потенциально годящихся на роль медиатора, может стать источником такой ситуации, когда у независимо появившихся нервных клеток медиатор будет одним и тем же. Насколько реальны такие ситуации, сейчас невозможно сказать, но нужно всё же иметь эту возможность в виду. С такой же неизбежностью встаёт на очередь вопрос о природе рецепторной специфичности. На наших скелетных мышцах эффект ацетилхолина блокируется кураре, на сердце — атропином. Но что за этими различиями стоит? Оттого ли различны постсинаптические холинорецепторы, что различны синаптические функции ацетилхолина (пусковое возбуждающее действие в одном случае, модуляция автоматизма — в другом)? Или дело в разном происхождении мышц (эктодермальное в одном случае, целомическое — в другом)? Или здесь важна какая-то третья причина? Не возьмусь отвечать и на этот вопрос, мало ещё фактов, но на некоторые биологические реальности хочу обратить внимание. Интересный вывод можно извлечь из сравнительно-физиологических данных об изменениях ионной проницаемости клеточной мембраны при взаимодействии медиатора с рецептором. Эти данные весьма однозначно показывают, что у высших организмов ионные эффекты медиаторов не более специфичны, чем у примитивных. Пока дело касается ионных каналов, эффект медиаторного вещества в равной степени избирателен — взята ли исследуемая мишень от организма с высокоразвитым мозгом или от очень простого многоклеточного. Фактическая сторона детально рассматривается Г. Гершенфельдом [166]. Фактом является и древность некоторых, во всяком случае, специфических воспринимающих субстанций. Известна группа алкалоидов, специфически блокирующих реакцию между медиатором и рецептором в синапсах млекопитающих (см. 2). Те же алкалоиды блокируют чувствительность к соответствующим медиаторным веществам на клетках беспозвоночных: действие гамма-аминомасляной кислоты снимается пикротоксином, глицина — стрихнином, ацетилхолина — атропином в одних случаях, тубокурарином — в каких-то других; при этом, кажется, вовсе не обязательно, чтобы у клеток, располагающих такими рецепторами, они участвовали в синаптических функциях. Приходит на ум пример с другим алкалоидом — колхицином, который оказывает на нейроны весьма специфическое действие, блокируя транспорт веществ по аксону. Никто не питает иллюзий относительно нейротропности колхицина: колхицин взаимодействует с тубулином, а обеспечение тубулином аксонного транспорта есть не более как частный случай внутриклеточных функций, выполняемых построенными из этого белка микротрубочками. Не вправе ли мы думать, что точно так же и макромолекулы, реагирующие с тубокурарином, стрихнином, бикукуллином и т. д., не появились в процессе эволюционного совершенствования синаптической передачи, а существовали до возникновения нервной системы, выполняя какие-то неизвестные нам клеточные функции? Несомненный и важный факт заключается, наконец, в том, что клетка может располагать и, как правило, располагает несколькими специфическими рецепторами к разным медиаторным веществам, а нередко и несколькими рецепторами к одному медиатору, которые порой размещаются в разных участках поверхностной мембраны. Единичный характер медиаторного химизма проявляется у нервных клеток только в секреторном плане, рецепции это не касается. Спекуляции о происхождении постсинаптических рецепторов и их молекулярной эволюции должны исходить из этой реальности. В связи со сказанным хотелось бы напомнить мысль А. Г. Гинецинского, что у скелетной мышцы позвоночных в процессе эволюции имело место «уточнение хеморецепции» путём сужения фармакологического спектра веществ, способных вызывать в мышце акт возбуждения. «Примитивная соматическая мышца, — писал Генецинский, — не обладает строго избирательной реактивностью к специализированным химическим раздражителям. Она поливалентна и отвечает стереотипным сокращением на многие биологически активные вещества» [14, стр. 5 — 6]. В принципе здесь мыслимы два возможных механизма «уточнения хеморецепции», и без специального исследования нельзя сказать, какова же примитивная ситуация: поливалентна ли единичная рецепторная субстанция, которая затем специализируется, претерпевая молекулярную эволюцию, или имеется исходная множественность рецепторных субстанций, часть которых затем утрачивается. Это вопрос, которым активно занимаются сравнительные фармакологи, и мы вскоре должны получить на него чёткий ответ. В любом случае ясно, что вопрос о природе и эволюции рецепторной специфичности должен решаться самостоятельно, и решение вопроса о природе медиаторной специфичности нервных клеток мало чем может здесь помочь.
7. 6. Заключение
Вэтой последней главе были рассмотрены некоторые следствия, вытекающие из представления о множественном происхождении нейронов и о консерватизме их медиаторного химизма, — а именно следствия, важные для лучшего понимания эволюции нервной системы. Было подчёркнуто, что о медиаторном консерватизме говорится только в том смысле, что в процессе эволюции нейроны наследуют предковый тип химизма независимо от той специализации, которую они приобретают при функциональной дифференциации. В пределах унаследованного типа медиаторный химизм, несомненно, подвержен эволюционным изменениям. Можно выделить три группы явлений, выражающих эволюционные изменения: 1) появление семейств медиаторов, каждое из которых имеет свой исходный предковый тип химизма; 2) совершенствование механизмов передачи, по-своему выраженное для каждого из медиаторов; и 3) отбор нейронов, т. е. элиминация одних химических типов нейронов и расцвет других. Вероятно, будут названы и другие проявления эволюционных изменений, касающихся химических синапсов. Существенно важно при анализе эволюционных закономерностей придерживаться правильной методологии — считать сравнимыми гомологичные системы нейронов и нервных окончаний.
8. ОБЩЕЕ ЗАКЛЮЧЕНИЕ
Среди всего, что есть в природе и изучается наукой, мозг человека был и долго ещё будет самым интригующим объектом. Наверняка, в этой области появятся новые подходы, как некогда возник кибернетический; однако эволюционные биологические подходы останутся главными — просто потому, что своим появлением в природе наш мозг обязан биологической эволюции. Путь, приведший к рождению мозга, не был строго предопределен; на разных его этапах имелись возможности выбора направлений развития, и реализация этих возможностей привела к тому, что в природе, помимо мозга человека, существует мозг пчелы или, скажем, осьминога. В каких-то важных отношениях, однако, выбор был невелик и возможности эволюции ограничивались свойствами исходного материала и тем, что этот материал мог меняться только в сфере действия биологических законов развития. В этой книге говорилось о фактах, позволяющих думать, что химическое разнообразие синаптических механизмов, представленных в нашем мозге, обусловлено не какой-то его высшей сложностью, а условиями возникновения нервной системы у примитивных многоклеточных животных. Такой вывод только на первый взгляд может показаться странным. На самом деле ситуация вполне банальна, и для теоретической биологии давно уже не секрет, что способ организации высокоразвитых биологических систем находится в теснейшей зависимости от их далекого и примитивного прошлого. «Структура высокоспециализированного органа заключает в себе всю историю его развития», — писал по этому поводу Л. А. Орбели [42]. Взгляд на нервную систему как на систему разнородных нейронов — систему, в которой каждая из составляющих её клеточных линий обладает собственным функциональным химизмом, но все вместе взаимодействуют в обеспечении единой нервной функции, — быть может, окажется полезным для понимания не только множественности медиаторных механизмов, но и других сторон работы мозга.ЛИТЕРАТУРА
1. Айрапетян С. Н. 1969. Влияние температуры на мембранный потенциал гигантских нейронов улитки. — Биофизика, 14, 1 - 15. 2. Алешин Б. В. 1972. Адренергические механизмы гипоталамуса.- Успехи совр. биол., 74, 142 - 163. 2а. Анохин П. К., Орлов И. В., Осиповский С. А. 1973. Изучение химической чувствительности и реакций на синаптическую стимуляцию нейронов Lymnaea stagnalis. — Нейрофизиология, 5, 510 - 518. 3. Беклемишев В. Н. 1964. Основы сравнительной анатомии беспозвоночных. Том 2, Органология. М., Наука. 4. Боровягин В. Л., Сахаров Д. А. 1968. Ультраструктура гигантских нейронов тритонии (атлас). М., Наука. 5. Бродский В. Я. 1966. Трофика клетки. М., Наука. 6. Бузников Г. А. 1967. Низкомолекулярные регуляторы зародышевого развития. М., Наука. 7. Вагнер Н. П. 1885. Беспозвоночные Белого моря. Том I. СПб. 8. Вепринцев Б. Н., Крастс И. В., Сахаров Д. А. 1964. Нервные клетки голожаберного моллюска Tritonia diomedia Bergh. — Биофизика, 9, 327 - 335. 9. Вержбинская И. А. 1964. Некоторые черты функциональной эволюции ферментных систем клеточного обмена нервной ткани позвоночных. — В кн. «Эволюция функций». М., Наука, стр. 219 - 228. 10. Герасимов В. Д. 1969. Роль малых концентраций ионов натрия в генерации потенциалов действия нейронами виноградной улитки. — Нейрофизиология, 1, 315 - 322. 11. Герасимов В. Д., Костюк П. Г., Майский В. А. 1964. Возбудимость гигантских нервных клеток различных представителей лёгочных моллюсков (Helix, Limnaea, Planorbis) в растворах, не содержащих ионов натрия.- Бюлл. эксперим. биол. и мед., 58, 3 — 7. 12. Герасимов В. Д., Холодова Ю. Д. 1971. Действие аминокислот на электрическую активность нейронов моллюсков Helix pomatia. — Журн. эвол. биох. физиол. 7, 156 — 161. 13. Гиляров М. С. 1964. Современные представления о гомологии. Успехи совр. биол., 57, 300 — 316. 14. Гинецинский А. Г. 1961. Об эволюции функций и функциональной эволюции. М., Изд-во АН СССР. 15. Гинецинский А. Г. 1970. Химическая передача нервного импульса и эволюция мышечной функции. Л., Наука. 16. Говырин В. А. 1967. Трофическая функция симпатических нервов сердца и скелетных мышц. Л., Наука. 17. Демин Н. Н. 1972. Некоторые пути влияния ацетилхолина на свойства мембранных структур нервной ткани. — В сб. «Физиологическая роль медиаторов». Казань, стр. 82. 18. Заварзин А. А. 1941. Очерки по эволюционной гистологии нервной системы. М., Медгиз. 19. Иванов А. В. 1940. Класс брюхоногих моллюсков (Gastropoda). В кн. «Руководство по зоологии», том 2. М., Биомедгиз, стр. 323 — 455. 20. Карякин А. В. 1971. Электрофизиологическое исследование идентифицированных нейронов париетальных ганглиев виноградной улитки. Автореф.канд. дисс. Кишинев. 21. Константинова М. С. 1971. Распределение моноаминов в гипоталамусе и гипофизе позвоночных. — Журн. эвол. биох. физиол., 7, 213-218. 22. Коробцов Г. Я. 1970. Физиологическое изучение моторных клеток велигеров эолидии. — В сб. «Проблемы биологии развития». М., Издание ИБР АН СССР, стр. 85 — 86. 23. Коробцов Г. Н., Сахаров Д. А. 1971. Роль ионной среды в реализации нервных и гормональных влияний на биения ресничек у личинок моллюсков. — Журн. эвол. биох. физиол., 7, 238 — 240. 24. Коробцов Г. Н., Сахаров Д. Л. 1971. Использование ацетилхолина и серотонина в анализе природы потенциалов, генерируемых нейронами улитки в безнатриевой среде. — В сб. «Биофизика мембран». Каунас, Издание Каунасского мед. института, стр. 486 — 494. 25. Коробцов Г. Н., Сахаров Д. А. 1973. Реакция нервных клеток на ионофоретическую аппликацию медиаторных веществ в разных ганглиях виноградной улитки. Тезисы докладов 5 Всесоюзн. конф. по нейрокибернетике. Ростов-на-Дону, стр. 153 — 154. 26. Коробцов Г. Н., Сахаров Д. А., Чернопятова Н. К. 1972. Физиология и ультраструктура пептидергических нейронов виноградной улитки. Тезисы докладов 6 Совещания по эвол. физиологии. Л., Наука, стр. 107 — 108. 27. Костюк П. Г. 1969. Ионные процессы в гигантских нейронах моллюсков. — Журн. эвол. биох. физиол., 5, 218 — 225. 28. Костюк П. Г. 1972. Нейрон. В кн. «Итоги науки и техники. Сер.: Физиология человека и животных». Том 10, стр. 5 — 39. 29. Коштоянц X. С. 1936. Об адреналиноподобных веществах в организме беспозвоночных животных (нервные узлы моллюсков). — Бюлл. эксперим. биол. и мед., 2, 41 — 43. 30. Коштоянц X. С. 1957. Об особенностях нервной регуляции и действия «медиаторов» у моллюсков. — Изв. АН АрмССР, 10, 13 — 15. 31. Коштоянц X. С. 1950. Энзимохимическая гипотеза возбуждения. — Физиол. журн. СССР, 36, 92 — 96. 32. Коштоянц X. С. 1957. Основы сравнительной физиологии, Том 2. М., Изд-во АН СССР. 33. Коштоянц X. С. 1963. Проблемы энзимохимии процессов возбуждения и торможения и эволюции функций нервной системы. М., Изд-во АН СССР. 34. Леонтьева Г. Р. 1966. Распределение катехоламинов в стенке кровеносных сосудов у круглоротых, рыб, амфибий и рептилий. — Журн. эвол. биох. физиол., 2, 31 — 35. 35. Майорова В. Ф., Троицкая Л. П. 1972. Аксо-соматические синапсы в педальных ганглиях брюхоногих моллюсков. — Докл. АН СССР, 204, 1471 — 1473. 36. Мандельштам Ю. Е. 1972. Структурно-функциональные особенности синаптического аппарата насекомых. — Журн. эвол. биох. физиол., 8, 395 — 403. 37. Манохина М. С, Кузьмина Л. В. 1971. Распределение биогенных моноаминов в центральной нервной системе тихоокеанского голожаберного моллюска Tritonia sp. — Журн. эвол. биох. физиол., 7, 357 — 361. 38. Митрополитанская Р. Л. 1941. О содержании ацетилхолина и холинэстеразы у простейших, губок и кишечнополостных животных. — Докл. АН СССР, 31, 717 — 718. 39. Михелъсон М. Я., Зеймаль Э. В. 1970. Ацетилхолин. Л., Наука. 40. Нистратова С. Я. 1965. Возможный механизм ускользания сердца беззубки из-под действия ацетилхолина. — Физиол. журн. СССР, 51, 1012 — 1016. 41. Нистратова С. Н., Турпаев Т. М. 1965. Механизм инактивации холинорецепторов при действии ацетилхолина на сердце беззубки. — Журн. эвол.биох. физиол., 1, 158 — 165. 42. Орбели Л. А. 1958. Основные задачи и методы эволюционной физиологии. — В кн. «Эволюция функций нервной системы». Л., Медгиз, стр. 7 — 17. 43. Орлов И. В., Шерстнев В. В. 1972. Микроионофоретический анализ химической чувствительности коры головного мозга. — В сб. «Физиологическая роль медиаторов». Казань, стр. 169 — 171. 44. Остроумова Н. К., Сахаров Д. А. 1973. Ультраструктура нейро-эффекторных окончаний у гастропод. Тезисы докладов 5 Всесоюзн. конф. по нейрокибернетике. Ростов-на-Дону, стр. 227 — 228. 45. Пархоменко Н. Т. 1970. К характеристике гигантских нейронов виноградной улитки, генерирующих потенциалы действия в растворах, не содержащих натрия. — Нейрофизиология, 2, 100 — 106. 46. Пархоменко Н. Т. 1971. Исследование гетерогенности ионных процессов поверхностной мембраны нейронов моллюсков. Автореф. канд. дисс. Киев. 47. Плотникова С. И., Говырин В. А. 1966. Распределение нервных элементов, содержащих катехоламины, у некоторых представителей кишечнополостных и первичноротых. — Архив анатомии, гистологии и эмбриологии, 50, 79 — 87. 48. Поленов А. Л. 1971. Гипоталамическая нейросекреция. Л., Наука. 49. Сахаров Д. А. 1960. Об автоматизме педальных ганглиев у крылоногого моллюска Clione limacina L. — Научн. докл. высш. школы (биол. науки), 3, 60 — 62. 50. Сахаров Д. А. 1962. Гигантские нервные клетки у голожаберных моллюсков Aeolidia papillosa и Dendronotus frondosus. — Журн. общ. биол., 23, 308-311. 51. Сахаров Д. А. 1965. Функциональная организация гигантских нейронов моллюсков. — Успехи совр. биол., 60, 365 — 383. 52. Сахаров Д. А. 1968. Некоторые вопросы нейробиологии брюхоногих. Тезисы докладов 5 Совещания по эвол. физиологии. Л., Наука, стр. 225. 53. Сахаров Д. А. 1968. Гомология отдельных нервных клеток и способы их идентификации в ганглиях брюхоногих. — В сб. «Моллюски и их роль в экосистемах». Л., Наука, стр. 8. 54. Сахаров Д. А. 1970. Основания к построению системы нервных клеток. — Журн. общ. биол., 31, 449 — 457. 55. Сахаров Д. А. 1971. Цитологические подходы к пониманию эволюции нервной системы брюхоногих моллюсков. — В сб. «Моллюски: пути, методы и итоги их изучения». Л., Наука, стр. 20 — 21. 56. Сахаров Д. А. 1972. Почему нейроны разные? — Природа, № 10, 52 — 62. 57. Сахаров Д. А., Нистратова С. Н. 1963. Особенности холинэргической реакции в сердце беззубки. — Физиол. журн. СССР, 49, 1475 — 1481. 58. Сахарова А. В., Сахаров Д. А. 1968. Люминесценция биогенных моноаминов на срезах нервной ткани, фиксированной водным формальдегидом. — Цитология, 10, 389 — 391. 59. Сахарова А. В., Сахаров Д. А. 1968. Дальнейшая разработка простого «водного» метода выявления внутриклеточных моноаминов. Люминесценция адренергических волокон в тканях теплокровных. — Цитология, 10, 1460 — 1466. 60. Сентаготаи Я., Флерко Б., Меш Б,, Халас Б. 1965. Гипоталамическая регуляция передней части гипофиза. Будапешт, Академиаи Киадо. 61. Сытинский И. А. 1972. Гамма-аминомасляная кислота в деятельности нервной системы. Л., Наука. 62. Турпаев Т. М. 1962. Медиаторная функция ацетилхолина и природа холинорецептора. М., Изд-во АН СССР. 63. Турпаев Т. М. 1967. Регуляция медиаторного процесса биохимическим механизмом обратной связи. В кн. «Биохимия и функция нервной системы». Л., Наука, стр. 234 — 243. 64. Турпаев Т. М., Нистратова С. Н., Сахаров Д. А. 1967. Эволюция холинергической регуляции сердечной деятельности у моллюсков.- Журн. общ. биол., 28, 618 — 626. 65. Турпаев Т. М., Сахаров Д. А. 1967. Эволюция холинергического медиаторного процесса. — Журн. эвол. биох. физиол., 3, 482 — 488. 66. Хлопин Н. Г. 1946. Общебиологические и экспериментальные основы гистологии. М., Изд-во АН СССР. 67. Чипенс Г. И., Папсуевич О. С. 1971. Анализ сигнатур нейрогипофизарных гормонов. — В сб. «Химия и биология пептидов». Рига, Зинатне, стр. 66 — 112. 68. Шилейко А. А. 1972. Некоторые аспекты изучения современных континентальных брюхоногих моллюсков. — В кн. «Итоги науки и техники. Сер.: Зоология беспозвоночных». Том I. М., ВИНИТИ, стр. 1 — 188. 69. Шуба М. Ф. 1972. Механизм возбуждающего и тормозящего действия катехоламинов на гладкомышечные клетки. — В сб. «Физиологическая роль медиаторов». Казань, стр. 255 — 256. 70. Экклс Дж. 1966. Физиология синапсов. М., Мир. 71. Экклс Дж. 1971. Тормозные пути центральной нервной системы. М., Мир. 72. Abraham A. 1965. Die Struktur der Synapsen im Ganglion viscerale von Aplysia californica. — Z. mikrosk.-anat. Forsch., 73, 45 — 59. 73. Agarwal R. A., Ligon R. J. В… Greenberg M. J. 1972. The distribution of cardioactive agents among molluscan species and tissues. — Compar. Gen. Pharmacol., 3, 249 — 260. 74. Akert K., Kawana E., Sandri С. 1971. ZIO-positive and ZIO-negative vesicles in nerve terminals. — Progr. Brain Res., 34,305 — 318. 75. Akert K., Sandri С. 1968. An electron-microscopic study of zinc iodide — osmium impregnation of neurons. I. Staining of synaptic vesicles at cholinergic junctions. — Brain Res., 7, 286 — 295. 76. Amin A. H., Crawford Т. В. В., Gaddum J. H. 1954. Distribution of substance P and 5-hydroxytryptamine in the central nervous system of dog. — J. Physiol., 126, 596 — 618. 77. Andén N.-E., Dahlström A., Fuxe K., Larsson K., Olson L., Ungerstedt U. 1966. Ascending monoamine neurons to the telencephalon and diencephalon. — Acta physiol. scand., 67, 313 — 326. 78. Andrew A. 1971. The origin on intramural ganglia. — J. Anat., 108, 169 — 184. 79. Andrews E. B. 1971. The fine structure of the nervous system of Bithynia tentaculata (Prosobranchia) in relation to possible neurosecretory activity. — Veliger, 14, 13 — 22. 80. Arch S. 1972. Polypeptide secretion from the isolated parietovisceral ganglion of Aplysia californica. — J. Gen. Physiol., 59, 47 — 59. 81. Arch S. 1972. Biosynthesis of the egg-laying hormone (ELH) in the bagcell neurons of Aplysia californica. — J. Gen. Physiol., 60, 102 — 119. 82. Arvanitaki A., Chalazonitis N. 1968. Electrical properties and temporal organization in oscillatory neurons (Aplysia). — In «Neurobiology of invertebrates». Budapest, Akad. Kiado, p. 169 — 199. 83. Ascher P. 1972. Inhibitory and excitatory effects of dopamine on Aplysia neurones. — J. Physiol., 225, 173 — 209. 84. Axelsson J. 1971. Catecholamine functions. Annual Rev. Physiol., 33,1 — 30. 85. Bargmann W., Lindner E., Andres К. К. 1967. Über Synapsen an endocrinen Epithelzellen und die Definition sekretorischer Neurone. — Z. Zellforsch., 77, 282 — 298. 86. Barker D. L., Herbert E., Hildebrand J. G., Kravitz E. A. 1972. Acetylcholine and lobster sensory neurones. — J. Physiol., 225, 205 — 229. 87. Barrantes F. J. 1970. The neuromuscular junctions of a pulmonate mollusc. I. Ultrastructural study. — Z. Zellforsch., 104,205 — 212. 87a. Barry J., Dubois M. P., Poulain P. 1973. LRF-producing cells of the mammalian hypothalamus: A fluorescent antibody study.- Z. Zellforsch., 146, 351 — 366. 88. Baumgarten H. G. 1972. Biogenic monoamines in the cyclostome and lower vertebrate brain. — Progr. Histochem. Cytochem., 4 (1). 89. Bennett J., Bueding E. 1971. Localization of biogenic amines in Schistosoma mansoni. — Compar. Biochem. Physiol., 39A, 859 — 867. 90. Bennett J. P., Jr., Logan W. J., Synder S. H. 1972. Amino acid neurotransmitter candidates: Sodium-dependent high-affinity uptake by unique synaptosomal fractions. — Science, 178, 997 — 999. 91. Bennett M. R. 1972. Autonomic neuromuscular transmission. Cambridge, Univ. Press. 92. Berry M. S., Cottrell G. A. 1973. Dopamine: excitatory and inhibitory transmission from a giant dopamine neurone.- Nature New Biol., 242,250 — 253. 93. Besson M. J., Cheramy A., Feltz P., Glowinski J. 1969. Release of newly synthesized dopamine from dopamine-containing terminals in the striatum of the rat. — Proc. Nat. Acad. Sci. USA, 62, 741-748. 94. Bjorklund A., Cegrell L, Falck В ., Ritzen M., Rosengren E. 1970. Dopamine-containing cells in sympathetic ganglia. -Acta physiol. scand., 78, 334 — 338. 95. Bjorklund A., Nobin A., Stenevi U. 1973. Effects of 5,6-dihydroxytryptamine on nerve terminal serotonin and serotonine uptake in the rat brain. — Brain Res., 53, 117 — 127. 96. Blankenship J. E., Wachtel H., Kandel E. R. 1971. Ionic mechanisms of excitatory, inhibitory, and dual synaptic actions mediated by an identified interneuron in abdominal ganglion of Aplysia. — J. Neurophysiol., 34, 76-92. 97. Bloom F. E. 1970. The fine structural localization of biogenic monoamines in nervous tissue. — Internat. Rev. Neurobiol., 13, 27 — 66. 98. Bloom F. E., Hoffer B. J., Siggins G. R., Barker J. L., Nicoll R. A. 1972. Effects of serotonin on central neurons: Microiontophoretic administration. — Fed. Proc, 31, 97 — 106. 99. Boehme D. H., Fordice M. W., Marks N., Vogel W. 1973. Distribution of glycine in human spinal cord and selected regions of brain. — Brain Res., 50, 353 — 359. 100. Bogusch G. 1972. Zur Innervation des glatten Penisretraktormuskels von Helix pomatia: allgemeine Histologie und Histochemie des monoaminergen Nervensystems. — Z. Zellforsch., 126,383-401. 101. Bowers C. Y., Schally A. V., Enzmann F., Boler J., Folkers K. 1970. Porcine thyrotropin releasing hormone is (pyro)glu-his-pro(NH2). — Endocrinology, 86, 1143-1153. 102. Bullock T. H., Horridge G. A. 1965. Structure and function in the nervous systems of invertebrates. San Francisco, Freeman. 102a. Burgus R., Butcher M., Amoss M., Ling N., Monahan M., Rivier J., Fellows R., Blackwell R., Vale W., Guillemin R. 1972. Primary structure of the ovine hypothalamic luteinizing hormone releasing factor. — Proc. Nat. Acad. Sci. USA, 69, 278 — 287. 102б. Burgus R., Dunn T. F., Desiderio D., Ward D. .V., Vale W., Guillemin R. 1970. Characterization of the ovine hypothalamic hypophysiotropic TSH-releasing factor. — Nature, 226, 321 — 325. 103. Burn J. H., Rand M. J. 1965. Acetylcholine in adrenergic transmission. — Annual Rev. Pharmacol., 5, 163 — 182. 104. Burnstock G. 1969. Evolution of the autonomic innervation of visceral and cardiovisceral systems in vertebrates. — Pharmacol. Revs, 21, 247 — 324. 105. Burnstock G. 1971. Neural nomenclature. — Nature, 229, 282-283. 106. Burnstock G. 1972. Purinergic nerves. — Pharmacol. Revs, 24, 509 — 581. 107. Burnstock G., Dumsday В., Smythe A. 1972. Atropine resistant excitation of the urinary bladder; the possibility of transmission via nerves releasing a purine nucleotide. — Brit. J. Pharmacol, 44, 451 — 461. 108. Burnstock G., Iwayama T. 1971. Fine-structural identification of autonomic nerves and their relation to smooth muscle. — Progr. Brain Res., 34, 389 — 404. 109. Campbell С. В. G., Hodos W. 1970. The concept of homology and the evolution of the nervous system. — Brain Behaviour. Evolut., 3, 353 — 367. 110. Cardot J. 1969. Mise en évidence de monoamines dans les fibres nerveuses du coeur des mollusques Helix pomatia Linné et Archachatina marginata Swainson par l'histochimie du fluorescence. — C. r. Acad. sci. Paris, 269, 345 — 347. 111. Carlsson A. 1965. Introductory remarks. In: «Pharmacology of cholinergic and adrenergic transmission». Praha, p. 157 — 163. 112. Carpenter D. O. 1973. Ionic mechanisms and models of endogenous discharge of Aplysia neurones. In «Neurobiology of invertebrates: Mechanisms of rhythm regulation». Budapest, Akad. Kiado, p. 35 — 58. 113. Carpenter D. O., Alving B. O. 1968. A contribution of an electrogenic Na+ pump to membrane potential in Aplysia neurons. — J. Gen. Physiol., 52, 1 — 21. 114. Chakrin L. W., Marchbanks R. M., Mitchell J. F., Whittaker V. P. 1972. The origin of the acetylcholine released from the surface of the cortex. — J. Neurochem., 19, 2727-2736. 115. Chalazonitis N. 1968. Synaptic properties of oscillatory neurones (Aplysia and Helix). In «Neurobiology of invertebrates». Budapest, Akad. Kiado, p. 201 — 226. 116. Chamberlain S. G., Kerkut G. A. 1969. Voltage clamp analysis of the sodium and calcium inward currents in snail neurones. — Compar. Biochem. Physiol., 28, 787 — 803. 117. Chase T. N., Murphy D. L. 1973. Serotonin and central nervous system function. — Annual Rev. Pharmacol., 13, 181 — 198. 118. Chen C. F., Baumgarten R. von, Takeda R. 1971. Pacemaker properties of completely isolated neurones in Aplysia californica. — Nature New Biol., 233, 27 — 29. 119. Chiarandini D. J., Stefani E., Gerschenfeld H. M. 1967. Inhibition of membrane permeability to chloride by copper in molluscan neurones. — Nature, 213,97-99. 120. Christoffersen G. R. J. 1972. Steady state contribution of the Na, K-pump to the membrane potential in identified neurons of a terrestrial snail, Helix pomatia. — Acta physiol. scand., 86, 498 — 514. 121. Cobb J. L. S. 1969. The distribution of mono-amines in the nervous system of echinoderms. — Compar. Biochem. Physiol., 28, 967 — 971. 122. Coggeshall R. E. 1967. A light and electron microscope study of the abdominal ganglion of Aplysia californica. — J. Neurophysiol., 30, 1263 — 1287. 123. Coggeshall R. E. 1970. A cytologic analysis of the bag cell control of egg laying in Aplysia. — J. Morphol., 132, 461 — 469. 124. Coggeshall R. E. 1971. A possible sensory-motor neuron in Aplysia californica. — Tissue and Cell, 3, 637 — 648. 125. Coggeshall R. E., Kandel E. R., Kupfermann L, Waziri R. 1966. A morphological and functional study of a cluster of identifiable neurosecretory cells in the abdominal ganglion of Aplysia californica. — J. Cell Biol., 31, 363 — 368. 126. Collier B., Mitchell J. F. 1967. The central release of acetylcholine during consciousness and after brain lesions. — J. Physiol., 188, 83 — 98. 127. Connor J. D. 1970. Caudate nucleus neurones: Correlation of the effects of substantia nigra stimulation with iontophoretic dopamine. — J. Physiol., 208, 691 — 703. 128. Consolo S., Garattini S., Ladinski H., Theonen H. 1972. Effect of chemical sympathectomy on the content of acetylcholine, choline and choline acetyltransferase activity in the cat spleen and iris. — J. Physiol., 220, 639 — 646. 129. Corrodi H., Jonsson G. 1967. The formaldehyde fluorescence method for the histochemical demonstration of biogenic monoamines. — J. Histochem. Cytochem., 15, 65 — 78. 130. Cottrell G. A. 1970. Direct postsynaptic responses to stimulation of serotonin-containing neurones. — Nature, 225, 1060 — 1062. 131. Cottrell G. A., Macon J. B. 1974. Synaptic connections of two symmetrically placed giant serotonin-containing neurones. — J. Physiol., 236, 435 — 464. 132. Cottrell G. A., Laverack M. S. 1968. Invertebrate pharmacology. — Annual Rev. Pharmacol., 8, 273 — 298. 133. Cottrell G. A., Osborne N. N. 1969. Localization and mode of action of cardioexcitatory agents in molluscan hearts. — Experientia, suppl., 15, 220 — 231. 134. Cottrell G. A., Osborne N. N. 1969. A neurosecretory system terminating in the Helix heart. — Compar. Biochem. Physiol., 28, 1455 — 1460. 135. Cottrell G. A., Osborne N. N. 1970. Subcellular localization of serotonin in an identified serotonin-containing neurone. — Nature, 225, 470 — 472. 136. Cottrell G. A., Pentreath V. W. 1970. Localization of catecholamines in the nervous system of a starfish, Asterias rubens, and of a brittlestar, Ophiothrix fragilis. — Compar. Gen. Pharmacol., 1, 73 — 81. 137. Cottrell G. A., Powell B. 1971. Formation of serotonin by isolated serotonin-containing neurons and by isolated non-amine-containing neurons. — J. Neurochem., 18, 1695 — 1698. 138. Curtis D. R., Crawford J. M. 1969. Central synaptic transmission — microelectrophoetic studies. — Annual Rev. Pharmacol., 1969, 9, 209 — 240. 139. Curtis D. R., Johnston G. A. R. 1970. Amino acid transmitters. — In «Handbook of neurochemistry», vol. 4. N. Y., Acad. Press, p. 115 — 134. 140. Dahl E., Falck В., Mecklenburg C. von, Myhrberg H. 1963. An adrenergic nervous system in sea anemones. — Quart. J. Microsc. Sci., 104, 531 — 534. 141. Dahl E., Falck В., Mecklenburg C. von, Myhrberg H., Rosengren E. 1966. Neuronal localization of dopamine and 5-hydroxytryptamine in some Mollusca. — Z. Zellforsch., 71, 489 — 498. 142. Dale H. H. 1933. Nomenclature of fibres in the autonomic system and their effects. — J. Physiol., 80, 10 — 11P. 142a. Dempsey G. P., Bullivant S., Watkins W. B. 1973. Ultrastructure of the rat posterior pituitary gland and evidence of hormone release by exocytosis as revealed by freeze-fracturing.- Z. Zellforsch., 143, 465 — 484. 143. Donovan В. Т. 1970. Mammalian neuroendocrinology. London, McGraw-Hill. 144. Dorsett D. A. 1967. Giant neurons and axon pathways in the brain of Tritonia. — J. Exper. Biol., 46, 137 — 151. 145. Dreifuss J. J., Gran J. D., Bianchi R. E. 1971. Antagonism between Ca and Na ions at neurohypophysial nerve terminals. — Experientia, 27, 1295 — 1296. 146. Dreifuss J. J., Kelly J. S. 1972. The activity of identified supraoptic neurones and their response to acetylcholine applied by iontophoresis. — J. Physiol., 220, 105 — 119. 147. Dunkan D., Morales R. 1973. Location of large cored synaptic vesicles in the dorsal gray matter of the cat and dog spinal cord. — Amer. J. Anat., 136, 123-127. 148. Ehinger В., Falck В., Persson H., Rosengren A.-M., Sporrong B. 1970. Acetylcholine in adrenergic terminals of the cat iris. — J. PhysioL, 209, 557 — 565. 149. Ehinger В., Myhrberg H. E. 1971. Neuronal localization of dopamine, noradrenaline, and 5-hydroxytryptamine in the central and peripheral nervous system of Lumbricus terrestris (L.). — Histochemie, 28, 265 — 275. 150. Emson P. C, Fonnum F. 1973. Application of microchemical analysis to the characterization of neurones from the brain of Helix aspersa. — Symposium on «simple» nervous systems. Glasgow. (An abstract). 151. Euler U. S. von. 1948. Identification of the sympathomimetic ergone in adrenergic nerves of cattle (Sympathin N) with laevo-noradrenaline. — Acta physiol. scand., 16, 63. 152. Faber D. S., Klee M. R. 1972. Membrane characteristics of bursting pacemaker neurones in Aplysia. — Nature New Biol., 240, 29 — 31. 153. Fischer H. 1971. Vergleichende Pharmacologie von Ubertragersubstanzen ir tiersystematischer Darstellung. Berlin, Springer. 154. Fischer H. 1972. Animal evolution in the field of synaptic substances. — Naturwissenschaften, 59, 425 — 435. 155. Florey E. 1967. Neurotransmitters and modulators in the animal kingdom. — Fed. Proc, 26, 1164 — 1178. 156. Florey E. 1971. Excitatory actions of GABA and of inhibitory neurons. — Experientia, 27, 1112 (An abstract). 157. Florey E. 1973. Acetylcholine as sensory transmitter in Crustacea. New evidence from experiments demonstrating release of ACh during sensory stimulation. — J. Compar. Physiol., 83, 1 — 16. 158. Foh E., Bogusch G. 1969. Die Nervennetze im Penisretraktor von Helix pomatia. — Z. Zellforsch., 93, 439 — 446. 159. Fonnum F., Storm-Mathisen J., Walberg F. 1970. Glutamate decarboxylase in inhibitory neurons. A study of the enzyme in Purkinje cell axons and boutons in the cat. — Brain Res., 20, 259 — 275. 160. Frazier W. Т., Kupfermann I., Coggeshall R. E., Kandel E. R., Waziri R. 1967. Morphological and functional properties of identified neurons in the abdominal ganglion of Aplysia californica. — J. Neurophysiol., 30, 1288 — 1351. 161. Fuxe K., Hokfelt Т., Ungerstedt U. 1968. Localization of indolealkylamines in CNS. — Advanc. Pharmacol., 6, 235 — 251. 161a. Fuxe K., Hokfelt T. 1971. Histochemical fluorescence detection of changes in central monoamine neurones provoked by drugs acting on the CNS. — Triangle, 10, 73 — 84. 162. Gainer H. 1971. Micro-disc electrophoresis in sodium dodecyl sulfate: An application to the study of protein synthesis in individual, identified neurons. — Analyt. Biochem., 44, 589 — 605. 162a. Gainer H. 1972. Patterns of protein synthesis in individual, identified molluscan neurons. — Brain Res., 39, 369 — 385. 162б. Gainer H. 1972. Electrophysiological behaviour of an endogenously active neurosecretory cell. — Brain Res., 39, 403 — 418. 163. Gersch M. 1971. General aspects of neuroendocrinology in insects. — Proc. XIII Internat. Congr. Entomology, vol. 1. Leningrad, Nauka, p. 87 — 92. 164. Gerschenfeld H. M. 1963. Observations on the ultrastructure of synapses in some pulmonate molluscs. — Z. Zellforsch., 60, 258 — 275. 165. Gerschenfeld H. M. 1971. Serotonin: Two different inhibitory actions on snail neurons. — Science, 171, 1252 — 1253. 166. Gerschenfeld H. M. 1973. Chemical transmission in invertebrate central nervous systems and neuromuscular junctions. — Physiol. Revs, 53. 167.Gerschenfeld H. M., Lasansky A. 1964. Action of glutamic acid and other naturally ocurring amino-acids on snail central neurons. — Internat. J. Neu ropharmacol., 3, 301 — 314. 168. Gerschenfeld H. M., Stefani E. 1966. An electrophysiological study of 5-hydroxytryptamine receptors ofneurones in the molluscan nervous system. — J. Physiol., 185, 684 — 700. 169. Giller E., Jr, Schwartz J. H. 1971. Choline acetyltransferase in identified neurons of abdominal ganglion of Aplysia californica. — J. Neurophysiol., 34, 93 — 107. 170. Glaizner B. 1968. Pharmacological mapping of cells in the subeosophageal ganglia of Helix aspersa. — In: «Neurobiology of invertebrates». Budapest, Akad. Kiado, p. 267 — 283. 171. Gola M., Romey G. 1971. Reponses anomales à des courants sous-liminaires de certaines membranes somatiques (neurone géants d'Helix pomatia). — Pflügers Arch. ges. Physiol., 327, 105 — 131. 172. Greenberg M. J., Agarwal R. A., Wilkens L. A., Ligon P. J. B. 1973. Chemical regulation of rhythmical activity in molluscan muscle. — In «Neurobiology of invertebrates: Mechanisms of rhythm regulation». Budapest, Akad. Kiado, p. 123 — 142. 173. Groat W. C. D., Lalley P. M., Saum W. R. 1972. Depolarization of dorsal root ganglia in the cat by GABA and related amino acids: Antagonism by picrotoxin and bicuculline. — Brain Res., 44, 273 — 277. 174. Haldeman S., Huffman R. D., Marshall K. C., McLennan H. 1972. The antagonism of the glutamate-induced and synaptic excitation of thalamic neurones. — Brain Res., 39, 419 — 425. 174a. Haldeman S., McLennan H. 1972. The antagonistic action of glutamic acid diethylester towards amino acid-induced and synaptic excitations of central neurones. — Brain Res., 45, 393 — 400. 175. Натре M. M. V., Tesch A., Jaeger C. P. 1969. On the nature of cardioactive substances from Strophocheilus oblongus ganglia. — Life Sci., 8 (Part 1), 827 — 835. 176. Hanstrbm B. 1925. Über die sogennannten Intelligenzspharen des Mollus kengehirns und die Innervation des Tentakels von Helix. — Acta zool. (Stockholm), 6, 183 — 215. 177. Hayos F., Kerpel-Fronius S. 1973. Comparative electron cytochemical studies of presynaptic and other neuronal mitochondria. — Brain Res., 62, 425 — 429. 178. Hehn G. von. 1970. Über den Feinbau des hyponeuralen Nervensystems des Seesterner (Asterias rubens L.). — Z. Zellforsch., 105, 137 — 154. 179. Heyer С. В., Kater S. B. 1973. Neuromuscular systems in molluscs. — Amer. Zool., 13, 247 — 270. 180. Histochemistry of nervous transmission (Ed. by O. Eränkö). 1971. Amsterdam, Elsevier («Progr. Brain Research», 34). 181. Hokfelt T. 1969. Distribution of noradrenaline storing particles in peripheral adrenergic neurons as revealed by electron microscopy. — Acta physiol. scand., 76, 427 — 440. 182. Hokfelt T. 1971. Ultrastructural localization of intraneuronal monoamines. Some aspects on methodology. — Progr. Brain Res., 34, 213 — 222. 182a. Hokfelt T. 1973. On the origin of small adrenergic storage vesicles: Evidence for local formation in nerve endings after chronic reserpine treatment. — Experientia, 29, 580 — 582. 183. Hokfelt Т., Fuxe K-, Goldstein M., Johansson O., Hösli T. 1974. Immunochemical evidence for the existence of adrenaline neurons in the rat brain. — Brain Res., 66, 235 — 261. 184. Hokfelt Т., Fuxe K., Goldstein M. 1973. Immunohistochemical localization of aromatic L-amino acid decarboxylase (DOPA decarboxylase) in central dopamine and 5-hydroxytryptamine nerve cell bodies of the rat. — Brain Res., 53, 175 — 180. 185. Holland N. D. 1970. The fine structure of the axial organ of the feather star Nemaster rubiginosa (Echinodermata: Crinoidea). — Tissue and Cell, 2, 625 — 636. 186. Hornykiewicz O. 1972. Dopamine and its physiological significance in brain function. In «The structure and function of the nervous tissue», vol. 6. N. Y., Acad. Press, p. 367 — 416. 187. Horridge G. A. 1967. The origin of the nervous system.- In «The structure and function of the nervous tissue», vol. 1. N. Y., Acad. Press, p. 1 — 31. 188. Horridge G. A. 1968. Interneurons. London, Freeman. 189. Hortnagl H., Winkler H., Lochs H. 1973. Immunological studies on a membrane protein (chromomembrin B) of catecholamine-storing vesicles. — J. Neurochem., 20, 977 — 986. 190. Hosly L., Haas H. L. 1972. The hyperpolarization of neurones of the medullaoblongata by glycine. — Experientia, 28, 1057 — 1058. 191. Hubbard J. I. , Quastel D. M. J. 1973. Micropharmacology of vertebrate neuromuscular transmission. — Annual Rev. Pharmacol., 13, 199 — 216. 192. Hughes G. M. 1968. The left and right giant neurons (LGC and RGC) of Aplysia. In «Neurobiology of invertebrates». Budapest, Akad. Kiado, p. 423 — 441. 193. Jordan L. M., Frederickson R. С. A., Phillis J. W., Lake N. 1972. Microelectrophoresis of 5-hydroxytryptamine: A classification of its action on cerebral cortical neurones. — Brain Res., 40, 552 — 558, 194. Iversen L. L. 1971. Role of transmitter uptake mechanisms in synaptic neurotransmission. — Brit. J. Pharmacol., 41, 571 — 591. 195. Jaeger C. P., Jaeger E. C., Welsh J. H. 1971. Localization of monoamine containing neurones in the nervous system of Strophocheilus oblongus (Gastropoda). — Z. Zellforsch., 112, 54 — 68. 196. Jahan-Parwar В., Smith M., Baumgarten R. von. 1969. Activation of neurosecretory cells in Aplysia by osphradial stimulation. — Amer. J. Physiol., 216, 1246 — 1257. 197. Jasper H., Koyama J. 1969. Rate of release of amino acids from the cerebral cortex in the cat as affected by brainstem and thalamic stimulation. — Canad. J. Physiol. Pharmacol, 47, 889 — 905. 198. Jenkin P. M. 1970. Animal hormones. Part II: Control of growth and metamorphosis. Oxford, Pergamon Press. 199. Johnson I. L. 1972. Glutamic acid as a synaptic transmitter in the nervous system. — Brain Res., 1972, 37, 1 — 20. 199a. Jongkind J. F. 1972. Neurophysin. — Progr. Brain Res., 38, 59 — 68. 200. Joosse J. 1972. Endocrinology of reproduction in molluscs. — Gen. Compar. Endocrinol., Suppl. 3, 591 — 601. 201. Kalimo H. 1971. Ultrastructural studies on the hypothalamic neurosecretory neurones of the rat. I. The paraventricular neurones of the non-treated rat. — Z. Zellforsch,, 122, 283 — 300. 202. Kanai Т., Szerb J. C. 1965. Mesencephalic reticular activating system and cortical acetylcholine output. — Nature, 205, 80 — 82. 203. Kandel E. R., Frazier W. Т., Coggeshall R. E. 1967. Opposite synaptic actions mediated by different branches of an identifiable interneuron, in Aplysia. — Science, 155, 346 — 349. 204. Kandel E. R., Tauc L. 1966. Input organization of two symmetrical giant cells in the snail brain. — J. Physiol., 183, 269 — 286. 205. Kandel E. R., Tauc L. 1966. Anomalous rectification in the metacerebral giant cells and its consequences for synaptic transmission. — J. Physiol., 183, 287 — 304. 206. Kater S. В., Kaneko C. R. S. 1972. An endogenously bursting neuron in the gastropod mollusc, Helisoma trivolvis.- J. Compar. Physiol., 79, 1 — 14. 207. Katz D., Miledi R. 1969. Tetrodotoxin-resistent electric activity in presynaptic terminals. — J. Physiol., 203, 459 — 488. 208. Kehoe J. 1972. Three acetylcholine receptors in Aplysia neurones. — J. Physiol., 225, 115 — 146. 209. Kerkut G. A. 1973. Catecholamines in invertebrates. — Brit. Med. Bull., 29, 100 — 104. 210. Kerkut G. A., French M. C., Walker R. J. 1970. The location of axonal pathways of identifiable neurones of Helix aspersa using the dye Procion Yellow M-4R. — Compar. Biochem. Physiol., 32, 681 — 690 211. Kerkut G. A., Horn N., Walker R. J. 1969. Long-lasting synaptic inhibition and its transmitter in the snail Helix aspersa. — Compar. Biochem. Physiol., 30, 1061 — 1074. 212. Kerkut G. A., Ralph K., Walker R. J., Woodruff G., Woods R. 1970. Excitation in the molluscan central nervous system. In «Excitatory synaptic mechanisms». Oslo, Univ. Press, p. 105 — 117. 213. Kerkut G. A., Sedden С. В., Walker R. J. 1967. Uptake of DOPA and 5-hydroxytryptophan by monoamine-forming neurones in the brain of Helix aspersa. — Compar. Biochem. Physiol., 23, 159 — 162. 214. Kerkut G. A., Walker R. J. 1962. The specific chemical sensitivity of Helix nerve cells. — Compar. Biochem. Physiol., 7, 277 — 288. 215. Kerkut G. A., York B. 1969. The oxygen sensitivity of the electrogenic sodium pump in snail neurones. — Compar. Biochem. Physiol., 28, 1125 — 1134. 216. Kilias R. 1960. Weinbergschnecken. Berlin, VEB Deutscher Verlag der Wissenschaft. 217. Kiss Т., Elekes K. 1972. Myo-neural junctions in the ventricle of the snail Helix pomatia L. — Acta biol. hung., 23, 207 — 209. 218. Klein R. L., Thureson-Klein A. 1971. An electron microscopic study of noradrenaline storage vesicles isolated from bovine splenic nerve trunk. — J. Ultrastr. Res., 34, 473 — 491. 219. Klemm N., Bjorklund A. 1971. Identification of dopamine and noradrenaline in nervous structures of the insect brain. — Brain Res., 26, 459 — 464. 220. Knowles F., Vollrath L. 1965. Synaptic contacts between neurosecretory fibres and pituicytes in the pituitary of the eel. — Nature, 206, 1168 — 1169. 221. Knowles F., Vollrath L. 1965. A functional relationship between neurosecretory fibres and pituicytes in the eel. — Nature, 208, 1343 — 1344. 222. Koike H., Eisenstadt M., Schwartz I. M. 1972. Axonal transport of newly synthesized acetylcholine in an identified neuron of Aplysia. — Brain Res., 37, 152 — 159. 222a. Konishi S., Otsuka M. 1974. The effects of Substance P and other peptides on spinal neurons of the frog. — Brain Res., 65, 397 — 410. 223. Krnjevic K. 1971. Synaptic transmission in the brain. — Klin. Wochenschr., 49, 519 — 523. 224. Krnjevic K. 1971. Conductance changes produced by neurotransmitters. — 3rd Internat. Meeting Internat. Soc. Neurochemistry. Budapest, Akad. Kiado, p. 164 (an abstract). 225. Kumar S. 1971. Glutamic acid as a precursor of ACh synthesis in rat brain in vivo. — Brain Res., 33, 578 — 581. 226. Kunze H. 1918. Über das ständige Auftreten bestimmter Zellelemente im Centralnervensystem von Helix pomatia L. — Zool. Anz., 49, 123 — 137. 226a. Kunze H. 1919. Zur Topographie und Histologie des Cerebralnervensystems von Helix pomatia L. — Z. wiss. Zool., 118, 25 — 203. 227. Kupfermann I. 1970. Stimulation of egg laying by extracts of neuroendocrine cells (bag cells) of abdominal ganglion of Aplysia. — J. Neurophysiol., 33, 877 — 881. 228. Labos E. 1969. Repetition-sensitive soma-response to de- and hyperpolarization of an identified Helix-neuron. — Acta physiol. hung., 36, 357 — 364. 229. Leclerc M., Delavault R. 1971. Présence de fibres nerveuses dans la paroicoelomique chez Asterina gibbosa Pennant (Echinoderme, Astéride). — C. r. Acad. sci. Paris, 272, 3311 — 3313. 230. Lemche H. 1956. The anatomy and histology of Cylichna (Gastropoda, Tectibranchia). — Spolia Zool. Mus. Hanniensis, 16, 1 — 278. 231. Lentz T. L. 1968. Primitive nervous systems. New Haven, Yale Univ. Press. 232. Levitan H., Tauc L. 1972. Acetylcholine receptors: Topographic distribution and pharmacological properties of two receptor types on a single molluscan neurone. — J. Physiol., 222, 537 — 558. 233. Libet В., Tosaka T. 1970. Dopamine as a synaptic transmitter and modulator in sympathetic ganglia: A different mode of synaptic action. — Proc. Nat. Acad. Sci. USA, 67, 667 — 673. 233a. Liebeswar G., Koester J., Goldman J. E. 1973. Identified motor neurons controlling circulation in Aplysia: Synthesis of transmitters and synaptic pharmacology. — Society for Neuroscience, 3rd Annual Meeting. San Diego, Calif., p. 288 (abstr. 47. 4). 234. Ljungdaht A., Hökfelt T. 1973. Accumulation of 3H-glycine in interneurons of the cat spinal cord. — Histochemie, 33, 277 — 280. 235. Loewi O. 1921. Über humorale Übertragbarkeit der Herznervenwirkung. I Mitteilung. — Pflügers Arch, ges., Physiol., 189, 239 — 242. 236. Loewi O. 1922. Über humorale Übertragbarkeit der Herznervenwirkung. II Mitteilung.-Pflügers. Arch. ges. Physiol., 193, 201 — 213. 237. Lunt G. G. 1973. Hydrophobic proteins from locust (Shistocerca gregaria) muscle with glutamate receptor properties. — Compar. Gen. Pharmacol., 4, 75 — 80. 237a. Manaranche R., L'Hermite P. 1973. Étude des amines biogènes de Glycera convoluta K. (Annelide Polychète). — Z. Zellforsch., 137, 21 — 36. 238. Marsden C, Kerkut G. A. 1970. The occurrence of monoamines in Planorbis corneus: A fluorescence microscopic and microspectrometric study. — Compar. Gen. Pharmacol., 1, 101 — 116. 238a. Matsuo H., Baba Y., Nair R. M. G., Arimura A., Schally A. V. 1971. Structure of the porcine LH and FSH releasing hormone. I. The proposed amino acid sequence. — Biochem. and Biophys. Res. Communs, 43, 1334 — 1339. 239. Mayeri E., Kupfermann I., Koester J., Kandel E. R. 1971. Neural coordination of heart rate and gill contraction in Aplysia. — Amer. Zool., 11,667 (An abstract). 240. McCaman M. W., Weinreich D., McCaman R. E. 1973. The determination of picomole levels of 5-hydroxytryptamine and dopamine in Aplysia, Tritonia, and leech nervous tissues. — Brain Res., 53, 129-137. 241. McCaman R. E., Dewhurst S. A. 1970. Choline acetyltransferase in individual neurons of Aplysia californica. — J. Neurochem., 17, 1421 -1426. 241a. McCaman R. E., Weinreich D., Borys H. 1973. Endogenous levels of acetylcholine and choline in individual neurons of Aplysia. — J. Neurochem., 21, 473 — 476. 242. McLennan H. 1970. Synaptic transmission. Philadelphia, Saunders. 243. Michelson M. J. 1969. Chairman's conclusions. — Proc. 4th Internat. Congr. Pharmacol., vol. 5. Basel, Schwabe, p. 128 — 130. 244. Mitchell J. F. 1963. The spontaneous and evoked release of acetylcholine from the cerebral cortex. — J. Physiol., 165, 98 — 116. 245. Mitchell J. F., Srinivasan V. 1969. Release of 3H-gamma-aminobutyric acid from the brain during synaptic inhibition. — Nature, 224, 663 — 666. 246. Moss R. L., Dyball R. E. J., Cross B. A. 1971. Response of antidromically identified supraoptic and paraventricular units to acetylcholine, noradrenaline, and glutamate applied iontophoretically. — Brain Res., 35, 573 — 576. 247. Murdoch L. L. 1971. Catecholamines in arthropods: A review. — Compar. Gen. Pharmacol., 2, 254 — 274. 248. Newby N. A. 1973. Habituation to light and spontaneous activity in the isolated siphon of Aplysia: Pharmacological observations. — Compar. Gen. Pharmacol., 4, 91 — 100. 249. Newman G., Kerkut G. A., Walker R. J. 1968. The structure of the brain of Helix aspersa. Electron microscope localization of cholinesterase and amines. — Sympos. Zool Soc. London, № 22, 1 — 18. 250. Nicoll R. A. 1971. Pharmacological evidence for GABA as the transmitter in granule cell inhibition in the olphactory bulb. — Brain Res., 35, 137 — 149. 251. Nicoll R. A., Barker J. L. 1971. The pharmacology of recurrent inhibition in the supraoptic neurosecretory system. — Brain Res., 35, 501 — 512. 252. Nisbet R. H., Plummer J. M. 1969. Functional correlates of fine structure in the heart of Achatinidae. — Experientia, Suppl. 15, 47 — 68. 253. Nobin A., Bjorklund A. 1973. Topography of the monoamine neuron systems in the human brain as revealed in fetuses. — Acta physiol. scand., Suppl. 388. 254. Nordmann J. J., Dreifuss J. J., Legros J. J. 1971. A correlation of release of polypeptide hormones and of immunoreactive neurophysin from isolated rat neurohypophyses. — Experientia, 27, 1344 — 1345. 255. Oomura Y., Ooyama H., Sawada M., Tanigawa T. 1972. Effects of strychnine on glycine depolarization in an Onchidium neuron. — J. Physiol. Soc. Japan, 34, 536. 256. Ooyama H., Oomura Y., Sawada M. 1965. Ionic mechanism of the hyperpolarization of an Onchidium neurone induced by glutamate. — Proc. 23rd Congr. Internat. Physiol. Sci. Tokyo. Abstr. 33. 257. Osborne N. N. 1971. Occurrence of GABA and taurine in the nervous systems of the dogfish and some invertebrates. — Compar. Gen. Pharmacol, 2, 433 — 438. 258. Osborne N. N. 1972. Occurrence of glycine and glutamic acid in the nervous system of two fish species and some invertebrates. — Compar. Biochem. Physiol, 43B, 579 — 585. 259. Osborne N. N. 1972. The in vivo synthesis of serotonin in an identified serotonin-containing neuron of Helix pomatia. — Internat. J. Neuroscience, 3, 215 — 228. 260. Osborne N. N., Cottrell G. A. 1970. Occurrence of noradrenaline and metabolites of primary catecholamines in the brain and heart of Helix. — Com par. Gen. Pharmacol, 1, 1 — 10. 261. Osborne N. N., Cottrell G. A. 1970. Transport of amines along the visceral nerve of Helix pomatia. — Z. Zellforsch., 109,171 — 179. 262. Osborne N. N., Cottrell G. A. 1971. Distribution of biogenic amines in the slug, Limax maximus. — Z. Zellforsch., 112, 15 — 30. 263. Osborne N. N., Cottrell G. A. 1972. Amine and amino acid microanalysis of two identified snail neurons with known characteristics. — Experientia, 28, 656 — 658. 264. Paupardin-Tritsch D., Gerschenfeld H. M. 1973. Transmitter role of serotonin in identifield synapses in Aplysia nervous system.- Brain Res., 58, 529-534. 264a. Paupardin-Tritsch D., Gerschenfeld H. M. 1973. Neuronal responses to 5-hydroxytryptamine resulting from membrane permeability decreases. — Nature New Biol., 244, 171 — 173. 265. Pentreath V. W., Cottrell G. A. 1971. «Giant» neurons and neurosecretion in the hyponeural tissue of Ophiothrix fragilis Abildgaard. — J. Exper. Marine Biol. Ecol., 6, 249 — 264. 266.Pentreath V. W., Cottrell G. A. 1973. Uptake of serotonin, 5-hydroxytryptophan and tryptophan by giant serotonin-containing neurones and other neurones in the central nervous system of the snail (Helix pomatia). — Z. Zellforsch., 143, 21 — 36. 266a. Pentreath V. W., Osborne N. N., Cottrell G. A. 1973. Anatomy of giant serotonin-containing neurones in the cerebral ganglia of Helix pomatia and Limax maximus.- Z. Zellforsch., 143, 1 — 20. 267. Peteya D. J. 1973. A light and electron microscope study of the nervous system of Ceriartheopsis americanus (Cnidaria, Ceriantharia).- Z. Zellforsch., 141, 301 — 317. 268. Phillis J. W. 1970. The pharmacology of synapses. Oxford, Pergamon Press. 269. Phillis J. W., Limacher J. J. 1974. Substance P excitation of cerebral cortical Betz cells. — Brain Res., 69, 158 — 163. 270. Pitman R. M. 1971. Transmitter substances in insects. A review. — Compar. Gen. Pharmacol., 2, 347 — 371. 271. Plotnikova S. I. 1968. The structure of the sympathetic nervous system of insects. — In «Neurobiology of invertebrates». Budapest, Akad. Kiado, p. 59 — 68. 272. Portig P. J., Vogt M. 1969. Release into the cerebral ventricles of substances with possible transmitter function in the caudate nucleus. — J. Physiol., 204, 687 — 715. 273. Pscheidt G. R. 1963. Biochemical correlates with phyletic division of the nervous system. — J. Theor. Biol., 5, 50 — 65. 274. Roberts P. J., Mitchell J. F. 1972. The release of amino acids from the hemisected spinal cord during stimulation. — J. Neurochem., 19, 2473 -2481. 275. Rogers D. C. 1968. Fine structure of smooth muscle and neuromuscular junctions in the optic tentacles of Helix aspersa and Limax flavus. — Z. Zellforsch., 89, 80 — 94. 275a. Rogers D. С 1969. Fine structure of smooth muscle and neuromuscular junctions in the foot of Helix aspersa. — Z. Zellforsch., 99, 315 — 335. 276. Rogers D. C. 1969. Fine structure of the epineural connective tissue sheath of the subesophageal ganglion in Helix aspersa. — Z. Zellforsch., 102, 99 — 112. 276a. Rogers D. C. 1969. The fine structure of the collar cells in the optic tentacles of Helix aspersa. — J. Zellforsch., 102, 113 — 128. 277. Rogers D. C. 1971. The fine structure of sensory neurons and their processes in the optic tentacles of Helix aspersa. — Z. mikrosk.-anat. Forsch., 84, 52 — 64. 278. Sakharov D. A. 1966. Recent studies of the properties of nudibranch nerve сells. — Compar. Biochem. Physiol., 18, 957 — 959. 279. Sakharov D. A. 1970. Cellular aspects of invertebrate neuropharmacology. — Annual Rev. Pharmacol., 10, 335 — 352. 280. Sakharov D. A. 1972. Electrophysiological study of factors controlling the activity of molluscan neurosecretory cells. — Gen. Compar. Endocrinol., 18, 621 — 622 (an abstract).Sakharov D. A. 1974. Evolutionary aspects of transmitter heterogeneity. J. Neural Transmission, Suppl. 11, 43 — 59. 281. Sakharov D. A., Borovyagin V. L., Zs.-Nagy I. 1965. Light, fluorescence and electron microscopic studies on «neurosecretion» in Tritonia diomedia Bergh (Mollusca, Nudibranchia). — Z. Zellforsch., 68, 660 — 673. 282. Sakharov D. A., Salánki J. 1969. Physiological and pharmacological identification of neurons in the central nervous system of Helix pomatia L. Acta physiol. hung., 35, 19 — 30. 283. Sakharov D. A., Salánki J. 1971. Study of neurosecretory cells of Helix pomatia by intracellular dye injection. — Expenentia, 27, 655 — 656. 284. Sakharov D. A., Turpajev T. M. 1968. Evolution of cholinergic transmission. — In «Neurobiology of invertebrates». Budapest, Akad. Kiado, p. 305 — 314. 285. Sakharov D. A., Zs.-Nagy I. 1968. Localization of biogenic monoamines in cerebral ganglia of Lymnaea stagnalis. — Acta biol. hung., 19, 145 — 157. 286. Sakharova A. V., Sakharov D. A. 1971. Visualization of intraneuronal monoamines by treatment with formalin solutions. — Progr. Brain Res., 34,11 — 26. 287. Salánki J. 1968. Studies on the effect of iontophoretically applied 1-glutamate on the giant nerve cells of gastropode (Helix and Limnaea). — Annal Biol. Tihany, 35, 75 — 81. 288. Salánki J., Vadász I., Elekes K. 1972. Physiological and morphological characteristics of Br-type neuron in the central nervous system of the snail Helix pomatia L. — Acta physiol. hung., 42, 243 — 254. 289. Salánki J., Kiss I. 1969. Identified cells in the central nervous system of Lymnaea stagnalis L. — Annal. Biol. Tihany, 36, 63 — 75. 290. Schon F., Iversen L. L. 1972. Selective accumulation of 3H-GABA by stellate cells in rat cerebellar cortex in vivo. — Brain Res., 42, 503 — 507. 291. Sedden С. В., Walker R. J., Kerkut G. A. 1968. The localization of dopamine and 5-hydroxytryptamine in neurones of Helix aspersa. — Sympos. Zool. Soc. London, № 22, 19 — 32. 292. Sheard M. H., Zolovick A. J. 1971. Serotonin: Release in cat brain and cerebrospinal fluid on stimulation of midbrain raphe. — Brain Res., 26, 455 — 458. 293. Sherman R. G., Atwood H. L. 1972. Correlated electrophysiological and ultrastructural studies of a crustacean motor unit. — J. Gen. Physiol., 59, 586 — 615. 294. Smith A. D. 1971. Summing up: Some implications of the neuron as a secreting cell. — Philos. Trans. Roy. Soc. London, B261, 423 — 437. 295. Smith U. 1970. The origin of small vesicles in neurosecretory axons. — Tissue and Cell, 2, 427 — 433. 296. Sneddon J. D., Smythe A., Satchell D., Burnstock G. 1973. An investigation of the identily of the transmitter substance released by non-adrenergic, non-cholinergic excitatory nerves supplying the small intestine of some lower vertebrates. — Compar. Gen. Pharmacol., 4, 53 — 60. 297. S.-Rózsa K., Perényi L. 1966. Chemical identification of the excitatory substance released in Helix heart during stimulation of the extracardial nerve. — Compar. Biochem. Physiol., 19, 105 — 113. 298a. S.-Rózsa K., Salánki J. 1973. Responses of central neurones to the stimulation of heart chemoreceptors in the snail, Helix pomatia L. — Annal. Bio Tihany, 40, 95 — 108. 298б. S.-Rózsa K., Salánki J. 1973. Single neurone responses to tactile stimulation of the heart in the snail. Helix pomatia L. — J. Compar. Physiol., 84, 267 — 279. 299. Stinnakre J., Tauc L. 1973. Calcium influx in active Aplysia neurones detected by injected aequorin.- Nature New Biol., 242, 113 — 115. 300. Stroch V. 1971. Biogene Amine in Rezeptororganen von Gastropoden (Prosobranchia, Opisthobranchia). — Z. Zellforsch., 115, 94 — 99. 301. Strumwasser F. 1962. Post-synaptic inhibition and excitation produced by different branches of a single neuron and the common transmitter involved. — Excerpta Med., Internat. Congress Series, № 48, 801 (an abstract). 302.Strumwasser F. 1971. The cellular basis of behaviour in Aplysia. — J. Psychiat. Res., 8, 237 — 257. 303. Suburo A. M., Pellegrino de Iraldi A. 1969. An ultrastructural study of the rat's suprachiasmatic nucleus. — J. Anat., 105, 439 — 446. 304 Szczepaniak A. C, Cottrell G. A. 1973. Biphasic action of glutamic acid and synaptic inhibition in an identified serotonin-containing neurone. — Nature New Biol., 241,62 — 64. 305. Tauc L. 1966. Physiology of the nervous system. — In «Physiology of Mollusca», vol. 2. N. Y., Acad. Press, p. 387 — 454. 306. Tauc L. 1967. Transmission in invertebrate and vertebrate ganglia. — Physiol. Revs, 47, 521 — 593. 307. Tauc L. 1969. Polyphasic synaptic activity. — Progr. Brain Res., 31, 247 — 2 57. 308. Tauc L., Gerschenfeld H. M. 1961. Cholinergic transmission mechanisms for both excitation and inhibition in molluscan central synapses. — Nature, 192, 366 — 367. 309. Tauc L., Gerschenfeld H. M. 1962. A cholinergic mechanism of inhibitory synaptic transmission in a molluscan nervous system. — J. Neurophysiol., 25, 236 — 262. 310. Tauc L., Hughes G. M. 1963. Modes of inhibition and propagation of spikes in the branching axons of molluscan central neurons. — J. Gen. Physiol., 46, 533 — 549. 311. Taylor K. M., Snyder S. H. 1973. The release of histamine from tissue slices of rat hypothalamus. — J. Neurochem., 21, 1215 — 1223. 312. Tebecis A. K., DiMaria A. 1972. Strichnine-sensitive inhibition in the medullary reticular formation: Evidence for glycine as an inhibitory transmitter. — Brain Res., 40, 373 — 383. 313. Thoenen H., Tranzer J. P. 1973. The pharmacology of 6-hydroxydopamine. — Annual Rev. Pharmacol., 13, 169 — 180. 314. Thureson-Klein Å., Klein R. L., Yen S.-S. 1973. Ultrastructure of highly purified sympathetic nerve vesicles: Correlation between matrix density and norepinephrine content. — J. Ultrastr. Res., 43, 18 — 35. 315. Toevs L. A., Brackenbury R. W. 1969. Bag cell-specific proteins and the humoral control of egg laying in Aplysia californica. — Compar. Biochem. Physiol., 29, 207 — 216. 316. Tranzer J. P., Thoenen H. 1967. Significance of empty vesicles in postganglionic sympathetic nerve terminals. — Experientia, 23, 123 — 124. 317. Turpaev T. M., Sakharov D. A. 1973. Comparative physiology of the mechanisms terminating the synaptic action of acetylcholine. — In «Comparative pharmacology», vol. 1. Oxford, Pergamon Press, p. 345 — 355. 318. Uchizono K. 1965. Characteristics of excitatory and inhibitory synapses in the central nervous system of the cat. — Nature, 205, 642 — 643. 319. Ungerstedt U. 1971. Stereotaxic mapping of the monoamine pathways in the rat brain. — Acta physiol. scand., Suppl. 367, part 1. 320. Usherwood P. N. R. 1972. Glutamate as an excitatory neuromuscular transmitter in insects. — Neurosci. Res. Progr. Bull., 10, 136 — 143. 321. Van Mol J.-J. 1967. Etude morphologique et phylogénétique du ganglion cérébroide des Gastéropodes Pulmonés (Mollusques). — Mém. Acad. roy. Belgique Cl. sci., 37, 1 — 168. 322. Vogt M. 1954. The concentration of sympathin in different parts of the central nervous system under normal conditions and after administration of drugs. — J. Physiol., 123, 451 — 481. 323. Vollrath L. 1969. Über die Herkunft «synaptischer» Bläschen in neurosekretorischen Axonen. — Z. Zellforsch., 99, 146 — 152. 324. Voskresenskaya A. K. 1968. The regulating function of the invertebrate nervous system. In «Neurobiologv of invertebrates». Budapest, Akad. Kiado, p. 367 — 380. 325. Wachtel В., Wilson W. A. 1973. Voltage clamp analysis of rhythmic slow wave generation in bursting neurones. In «Neurobiology of invertebrates: Mechanisms of rhythm regulation». Budapest, Akad. Kiado, p. 59 — 78. 325a. Wald F. 1972. Ionic differences between somatic and axonal action potentials in snail giant neurones. — J. Physiol., 220, 267 — 281. 326. Walker R. J., Crossman A. R., Woodruff G. N., Kerkut G. A. 1971. The effect of bicuculline on the gamma-aminobutyric acid (GABA) receptors of neurones of Periplaneta and Helix aspersa. — Brain Res., 34, 75 — 82. 327. Walker R. J., Lambert J. D. C, Woodruff G. N., Kerkut G. A. 1970. Action potential shape and frequency as criteria for neuron identification in the snail, Helix aspersa. — Compar. Gen. Pharmacol., 1, 409 — 425. 328. Walker R. J., Ralph K. L., Woodruff G. N., Kerkut G. A. 1971. Evidence for a dopamine inhibitory post-synaptic potential in the brain of Helix aspersa. — Compar. Gen. Pharmacol., 2, 15 — 26. 329. Weight F. F., Votava J. 1971. Inactivation of potassium conductance in slow postsynaptic excitation. — Science, 172, 504. 330. Weinreich D., Dewhurst S. A… McCaman R. E. 1972. Metabolism of putative transmitters in individual neurones of Aplysia californica: Aromatic amino acid decarboxylase. — J. Neurochem., 19, 1125 — 1130. 331. Weinreich D., McCaman M. W., McCaman R. E., Vaughn J. E. 1973. Chemical, enzymatic and ultrastructural characterization of 5-hydroxytryptamine-containing neurons from the ganglia of Aplysia californica and Tritonia diomedia. — J. Neurochem., 20, 969 — 976. 332. Welsh J. H. 1957. Serotonin as a possible neurohumoral agent: Evidence obtained in lower animals. — Ann. N. Y. Acad. Sci., 66, 618 — 630. 333. Welsh J. H. 1971. Neurohumoral regulation and the pharmacology of a molluscan heart. — Compar. Gen. Pharmacol., 2, 423 — 432. 334. Welsh J. H. 1972. Catecholamines in the invertebrates. In «Handbook of experimental Pharmacology», vol. 33. Stuttgart, Springer, p. 79 — 109. 335. Welsh J. H., Williams L. D. 1970. Monoamine-containing neurons in Planaria. — J. Compar. Neurol., 138, 103 — 112. 336. Wendelaar Bonga S. E. 1970. Ultrastructure and histochemistry of neurosecretory cells and neurohaemal areas in the pond snail Lymnaea stagnalis (L.). — Z. Zellforsch., 108, 190 — 225. 337. Werman R. 1966. A review — criteria for identification of a central nervous system transmitter. — Compar. Biochem. Physiol., 18, 745 — 766. 338. Werman R. 1969. An electrophysiological approach to drug-receptor mechanisms. — Compar. Biochem. Physiol., 30, 997 — 1017. 339. Werman R. 1970. Amino acids as central neurotransmitters. In «Neurotransmitters» (Res. Publ. Assoc. Res. Nerv. Mental. Disease, vol. 50), p. 147 — 181. 340. Werman R. 1972. CNS cellular level: Membranes. — Annual Rev. Physiol., 34, 337 — 374. 341. Westfall J. A. 1970. Ultrastructure of synapses in a primitive coelenterate. — J. Ultrastr. Res., 32, 237 — 246. 342. Westfall J. A. 1970. Synapses in a sea anemone, Metridium (Anthozoa). — 7th Internat. Congr. Electron Microscopy, vol. 3. Paris, p. 717 (an abstract). 342a. Westfall J. A. 1973. Ultrastructural evidence for a granule-containing sensory-motor-interneuron in Hydra littoralis. — J. Ultrastr. Res., 42, 268 — 282. 343. Westfall J. A., Yamataka S., Enos P. D. 1971. Ultrastructural evidence of polarized synapses in the nerve net of Hydra. — J. Cell Biol., 51, 318 — 323. 344. Whittaker V. P. 1973. The biochemistry of synaptic transmission. — Naturwissenschaften, 60, 281 — 289. 345. Whittaker V. P., Dowe G., Scotto J. 1971. Vesiculine: A possible counter-ion for acetylcholine in the cholinergic synaptic vesicle. — 3rd Internat. Meeting Internat. Soc. for Neurochem., Budapest, Akad. Kiado, p. 266 (an abstract). 346. Wilgenburg H. van, Leeuwen A. A. van. 1971. Pharmacological charecterization of neurons in the visceral and parietal ganglia of Helix pomatia, — Currents in Modern Biology, 3, 335 — 341. 347. Willows A. O. D., Hoyle G. 1968. Correlation of behaviour with the activity of single identifiable neurons in the brain of Tritonia. In «Neurobiology of invertebrates». Budapest, Akad. Kiadó, p. 443 — 461. 347a. Willows A. O. D., Dorsett D. A., Hoyle G. 1973. The neuronal basis of behaviour in Tritonia. I. Functional organization of the central nervous system. — J. Neurobiol., 4, 207 — 237. 348. Wolman M. 1971. A fluorescent histochemical procedure for gamma-aminobutyric acid. — Histochemie, 28, 118-130. 349. Woods R. I. 1969. Acrylic aldehyde in sodium dichromate as a fixative for identifying catecholamine storage sites with the electron microscope. — J. Physiol., 203, 35 — 36P. 350. York В., Twarog В. М. 1973. Evidence for the release of serotonin by relaxing nerves in molluscan muscle. — Compar. Biochem. Physiol., 44A, 423 — 430. 351. York D. H. 1967. The inhibitory action of dopamine on neurones of the caudate nucleus. — Brain Res., 5, 263 — 266. 352. Zs.-Nagy I. , Sakharov D. A. 1969. Axo-somatic synapses in procerebrum of Gastropoda. — Experientia, 25, 258 — 259. 353. Zs.-Nagy I. , Sakharov D. A. 1970. The fine structure of the procerebrum of pulmonate molluscs, Helix and Limax. — Tissue and Cell, 2, 399 — 411.ОГЛАВЛЕНИЕ 1. Введение… 3 2. Неоднородность клеточного состава нервной системы млекопитающих…7 2.1. Терминология… 7 2.2. Медиаторная специфичность нейрона и идентификация медиатора… 8 2.3. Типы нейронов у млекопитающих… 12 2.3.1. Холинергические нейроны… 12 2.3.2. Моноаминергические нейроны… 16 2.3.3. Нейроны, имеющие медиаторами аминокислоты… 22 2.3.4. Пептидергические нейроны… 25 2.3.5. Пуринергические нейроны… 29 2.4. Заключение… 29 3. Постановка вопроса о природе медиаторной специфичности… 31 3.1. Аналогия или гомология?… 31 3.2. Гипотеза полигенеза… 32 3.3. Альтернатива: функциональная специализация… 33 3.4. Что важно знать для проверки гипотез… 35 4. Неоднородность клеточного состава нервной системы виноградной улитки… 37 4.1. Виноградная улитка… 37 4.2. Результаты электрофизиологических исследований… 41 4.2.1. Развитие исследований на идентифицируемых нейронах гастропод… 41 4.2.2. Обозначение нейронов и их визуальная идентификация 44 4.2.3. Электрофизиологические критерии идентификации 47 4.2.4. Специфичность клеточных рецепторов… 54 4.2.5. Идентифицированные клетки… 59 4.3. Гистохимическая локализация медиаторных биогенных аминов… 70 4.3.1. Метод формальдегидной конденсации и его варианты… 70 4.3.2. Моноаминергические нейроны виноградной улитки… 71 4.4. Электронно-микроскопическое исследование… 77 4.4.1. История вопроса… 77 4.4.2. Материал… 80 4.4.3. Клетки и синапсы ЦНС… 81 4.4.4. Нейро-эффекторные окончания… 83 4.5. Заключение… 93 5. Клеточные гомологии в нервных системах гастропод… 95 5.1.Критерии установления гомологии… 95 5.2. Сравнительная анатомия о гомологии нервных органов у гастропод… 96 5.3. Установление клеточных гомологий… 100 5.3.4. Гомологи катехоламиновых нейронов педальных ганглиев… 111 5.3.5. Другие примеры клеточных гомологий… 117 5.4. Заключение… 122 6. Происхождение специфического химизма нервных клеток… 125 6.1. Медиаторная специфичность нейронов гастропод… 125 6.2. У кого больше медиаторов?… 133 6.3. Каковы отношения между медиаторной специфичностью и функциональной специализацией нейрона?… 134 6.4. Специфичен ли химизм примитивных нейронов?… 138 6.5. Един ли гистогенетический источник нервных клеток?… 141 6.6. Трудности функционального объяснения… 143 6.7. Трудности гипотезы полигенеза нейронов… 144 6.8. Заключение… 148 7. Эволюционная сторона проблемы специфичности нейронов… 150 7.1. Методологические замечания… 150 7.2. Эволюция медиаторов и естественная система нервных клеток… 151 7.3. Совершенствование механизмов передачи… 154 7.4. Неравноценность медиаторов и отбор нейронов… 157 7.5. Вопросы, встающие на очередь… 160 7.6. Заключение… 163 8. Общее заключение… 164 Литература…165
Дмитрий Антонович Сахаров Генеалогия нейронов Утверждено к печати Институтом биологии развития Академии наук СССР Редактор Э. А. Фролова Художник А. А. Кущенка Художественный редактор Я. Я. Власик Технические редакторы Э. Л. Кунина, А. М. Сатарова Сдано в набор 2/VIII 1974 г. Подписано к печати 28/Х 1974 г. Формат 60X90 1/16. Бумага № 2. Усл. печ. л. 11,5. Уч.-изд. л. 12,8. Тираж 1700 экз. Т-15993. Тип. зак. 966. Цена 86 коп. Издательство «Наука». 103717 ГСП, Москва, К-62, Подсосенский пер., 21, 2-я типография издательства «Наука». 121099, Москва, Г-99, Шубинский пер, 10

©1974 ИЗДАТЕЛЬСТВО «НАУКА»

Последние комментарии
15 минут 51 секунд назад
18 минут 46 секунд назад
2 дней 6 часов назад
2 дней 11 часов назад
2 дней 12 часов назад
2 дней 14 часов назад